
应用线粒体基因序列分析虎的进化关系
2014-11-10 08:21石俊松田存锋许继国张秀娟张守全
华南农业大学学报 2014年2期
石俊松,田存锋,许继国,张秀娟,李 莉,张守全
(华南农业大学动物科学学院,广东广州 510642)
虎属于哺乳纲、食肉目、猫科、豹属、虎种,一般虎被分为8个亚种,现仅存5个亚种.在这仅存的5个亚种中,华南虎 Panthera tigris amoyensis、东北虎P.tigris altaica和孟加拉虎P.tigris tigris也濒临灭绝,尤其是华南虎.为了成功地保护物种,除了要保护其栖息地和最大限度地增加种群数量外,还必须保护其遗传多样性.遗传多样性贫乏的物种,在环境中将十分脆弱,因而其进化前景暗淡[1].
为了制定有效的繁殖对策,以最大限度地保存圈养虎群体的遗传多样性,必须首先弄清起源群体的遗传结构,这是任一圈养动物繁殖中迫切需要解决的问题.由于虎类是一个比较年轻的类群,亚种间基因进化较保守,单一基因的信息量尚不足以解决该类群的系统发育问题,增加新的DNA序列的研究是解决问题的关键之一.同时,采用多基因序列数据也是由基因树到物种树的必然要求.近年来,人们开始注意线粒体DNA(mtDNA)在系统进化研究中的作用,同时人们已经把它们用于猫科动物的系统进化研究[2-3].哺乳动物的线粒体DNA是双链环状DNA,能够独立复制,不与组蛋白结合,另外,线粒体DNA所具有的线粒体基因组复制与表达所需的多种酶则由核基因编码,因此,线粒体是一个半自主性细胞器[4].White等[5]研究发现线粒体 DNA 没有内含子,缺乏有效的基因修复系统,极易发生突变,与核DNA相比,mtDNA具有母系遗传、拷贝数多、突变率高和极少发生重组等特性,是研究分子生态学、系统进化和群体遗传学的一个关键工具;其中mtDNA高突变率和极少发生重组的特性,使得较短时间内积累的变异可以连续而忠实地遗传下去,通过分析这些变异的分布范围与频率等信息,从而可以更好地研究种群关系及系统进化.
郑涛[6]利用中国产13种猫科动物的12SrRNA基因(约371 bp)和Cytb部分序列(约355 bp)进行了分析,其构建的进化树显示了云豹、豹、雪豹和虎具有较近的亲缘关系,支持将它们同归于豹属的观点.Luo等[3]对134份虎样本的4 kb mtDNA序列进行了分析,提出现存的虎有6个亚种:东北虎、北印支虎、华南虎、马来虎、苏门答腊虎和孟加拉虎.张文平等[7]利用线粒体DNA的D-loop区及ND5部分序列构建了东北虎、孟加拉虎和华南虎的系统进化树.韦鹍[8]利用mtDNA的ND5基因研究了华南虎的遗传多样性.本文主要利用mtDNA COⅠ基因、COⅢ基因和ND4基因部分序列来构建东北虎、孟加拉虎和华南虎的系统进化树,探讨3种虎的进化关系,也为虎的物种树的建立提供一些依据.
1 材料与方法
1.1 材料来源和mtDNA提取
本试验采集了广西熊虎山庄东北虎和孟加拉虎的血液样品,每种取3~4个个体,其中东北虎个体编号为Pta01、Pta02、Pta03和Pta04,孟加拉虎编号为Ptt01、Ptt02、Ptt03和Ptt04.将虎麻醉后从尾部静脉采全血10 mL,按 V(血液)∶V(ACD 抗凝剂)=6∶1的比例混匀抗凝,于-80℃条件保存备用.华南虎肌肉组织样来自广州动物园自然死亡的1头华南虎,编号为Pts01.
线粒体DNA的提取结合了DNA总基因组和质粒DNA的提取方法,并对常规酚-氯仿抽提法改进.所得的mtDNA溶于TE缓冲液中,-20℃条件保存备用.
1.2 PCR 扩增
根据从NCBI GenBank上搜索得到的家猫Felis catus(NC_001700)、猎豹 Acinonyx jubatus(NC_005212,)、云豹 Neofelis nebulosa(NC_008450) 的mtDNA全序列,利用DNAssist 1.0比对,选取序列的保守区,利用Primer permier 5.0软件设计引物,并由Oligo检验,最后由上海生工生物工程技术服务有限公司合成.COⅠ引物序列为:F-GCTCGAACCTCTGTCTTTAG,R-CCAAAACCGGGTAGGATTAA;COⅢ引物序列为:F-GCCTATGTTTTTACCCTGCT,R-CTACGTCTACGAAGTGTCAA;ND4引物序列为:F-GGCAATCAAACAGAGCG,R-GGAAGTTAGTCCGTGGG.
PCR扩增条件:25 μL反应体系含50 ng基因组模板,1 × PCR 缓冲液,8 μmol/L 引物,200 μmol/L dNTP,1.5 mmol/L Mg2+,1 U Taq DNA 聚合酶.94 ℃预变性5 min;94℃变性30 s,55℃退火30 s,35个循环;72℃延伸50 s,72℃后延伸5 min,4℃条件下保存产物.
1.3 产物的回收和测序
PCR产物经凝胶电泳检测后,由DNA胶回收试剂盒(上海生工)回收纯化,并与 pMD18-T载体(TaKaRa公司)连接,然后转化JM109感受态细胞后,经蓝白筛选,送阳性克隆的菌液到上海英骏生物科技有限公司测序.
1.4 序列分析及构建进化树
所有测得的序列比对(Sequence alignment)使用DNAStar软件包的 EditSeq、SeqMan、Megalign 等软件进行编辑,并进行人工校对,以确保序列的准确性.然后用DNAssit分析各物种间各个序列间的差异,单倍型多样度.用DNAStar等计算序列的碱基组成及序列间的转换/颠换值、遗传距离,并计算序列的碱基组成百分率等.
选择家猫、猎豹及云豹的mtDNA作为外群,利用Mega 4.0采用邻接法(Neighbour-joining,NJ)重建华南虎、东北虎、孟加拉虎的进化关系树,系统树中节点的自助置信水平(Bootstrap confidence level,BCL)应用自引导估计,共1000次循环.
2 结果
2.1 华南虎、孟加拉虎和东北虎的mtDNA序列
通过测序,共得到25个序列,其中COⅠ基因8个个体(东北虎3个、孟加拉虎4个、华南虎1个)各755 bp(NCBI GenBank登录号:FJ422123,FJ422145,FJ455122,FJ455124,FJ455125,FJ461529 ~461531);COⅢ基因8个个体(东北虎4个、孟加拉虎4个)各742 bp,华南虎1个个体737 bp(NCBI GenBank登录号:FJ461532 ~461534,FJ465508 ~465511,FJ469625);ND4基因8个个体(东北虎3个、孟加拉虎4个、华南虎1个)各561 bp(NCBI GenBank登录号:FJ608583~608585,FJ694968 ~694972).
2.2 序列差异及分析
2.2.1 COⅠ基因 通过DNAssist软件对这些序列的优化及比对,发现3只东北虎中出现3种线粒体COⅠ单倍型,共有2个变异位点;4只孟加拉虎出现2种线粒体COⅠ单倍型,只有1个变异位点.
由序列比对可知,8个个体的序列共检测到多态位点13个,占分析位点总数(750 bp)的1.73%,2核苷酸间的变异位点13个,无3核苷酸间的变异位点.因此,3个虎亚种的mtDNA COⅠ基因序列变异主要有转换和颠换2种类型,无转换与颠换共存的现象.东北虎、孟加拉虎、华南虎和云豹的序列两两碱基替换情况和遗传距离列于表1.
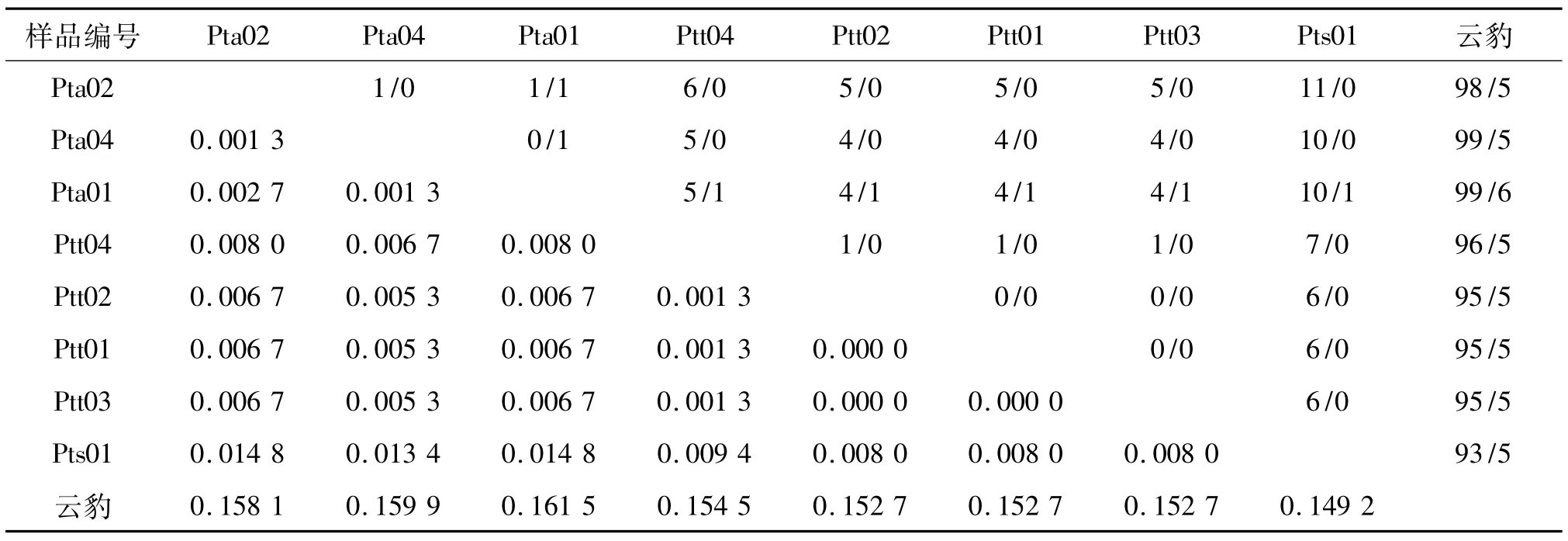
表1 东北虎、孟加拉虎、华南虎和云豹的COⅠ序列间的转换/颠换(上三角)和遗传距离(下三角)Tab.1 Numbers of transition/transversion(upper-right matrix)and genetic distances(Kimura 2-parameter model,lowerleft matrix)for COⅠsequences of Siberian tiger,Bengal tiger,South China tiger and clouded leopard
从表1中可以看出:虎亚种间的Kimura 2-parameter遗传距离范围为0.0013~0.0148,变异程度较大,亚种之间距离最小的为东北虎和孟加拉虎,最大的为东北虎同华南虎;东北虎个体之间的遗传距离和孟加拉虎个体之间遗传距离均小于亚种之间的遗传距离.云豹和虎亚种之间的距离范围是0.1492~0.1615,其中和华南虎最小,和东北虎最大.
2.2.2 COⅢ基因 对4个东北虎个体、1个华南虎个体和4个孟加拉虎个体的mtDNA进行了克隆测序,得到了这9个个体各737 bp的序列.通过DNAssist软件对这些序列的优化及比对,发现4只东北虎中只有1种线粒体COⅢ基因单倍型,4只孟加拉虎也只有1种线粒体COⅢ基因单倍型.可见在虎中线粒体COⅢ基因个体差异极小.
由序列比对可知,9个个体的序列共检测到多态位点4个,约占分析位点总数(737 bp)的0.54%,2核苷酸间的变异位点4个,无3核苷酸间的变异位点.因此,3个虎亚种的mtDNA COⅢ基因序列变异主要有转换和颠换2种类型,无插入/缺失和转换与颠换共存的现象.东北虎、孟加拉虎、华南虎和云豹的序列两两碱基替换情况和遗传距离列于表2.
从表2中可以看出:虎亚种间的Kimura 2-parameter遗传距离范围为0.0000~0.0055,变异程度较大,遗传距离最大为东北虎和华南虎,而东北虎和孟加拉虎、华南虎和孟加拉虎之间的遗传距离是一样的(D=0.0027).东北虎个体之间、孟加拉虎个体之间遗传距离均为0.云豹和虎亚种之间的遗传距离范围是0.1181~0.1249,变异程度较小,其中和华南虎遗传距离最小,和东北虎遗传距离最大.
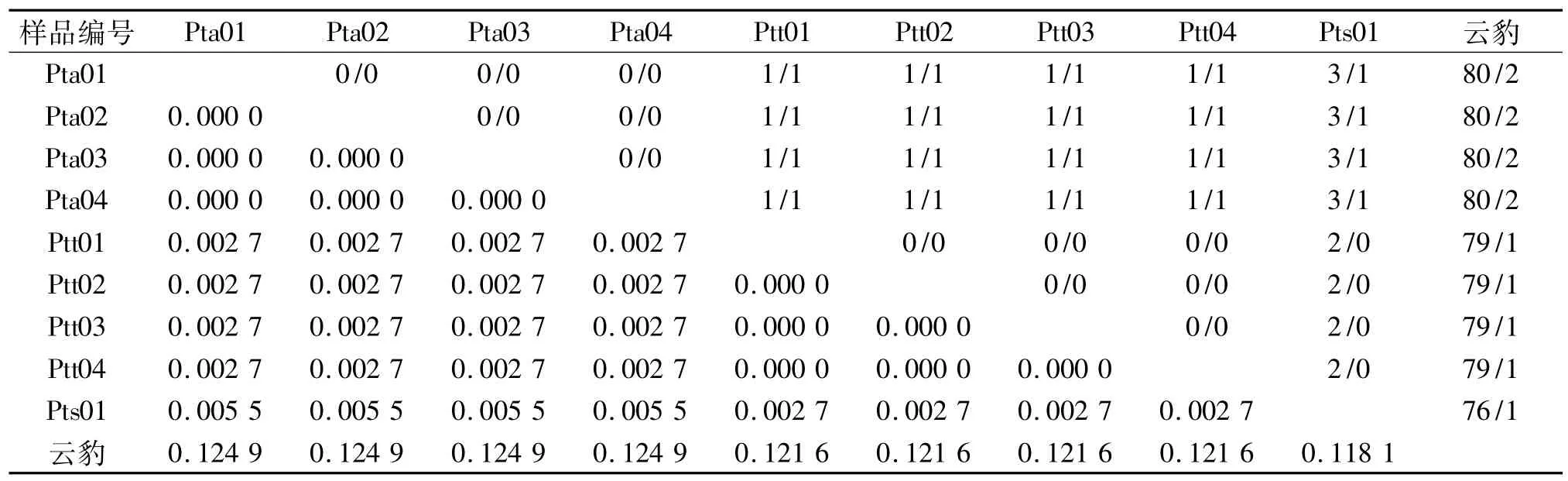
表2 东北虎、孟加拉虎、华南虎和云豹的COⅢ序列间的转换/颠换(上三角)和遗传距离(下三角)Tab.2 Numbers of transition/transversion(upper-right matrix)and genetic distances(Kimura 2-parameter model,lowerleft matrix)for COⅢsequences of Siberian tiger,Bengal tiger,South China tiger and clouded leopard
2.2.3 ND4基因 对3个东北虎个体、1个华南虎个体和4个孟加拉虎个体的mtDNA进行了克隆测序,得到了这8个个体各561 bp的序列.通过DNAssist软件对这些序列的优化及比对,发现3只东北虎只有1种线粒体ND4单倍型;4只孟加拉虎有2种线粒体ND4单倍型,总共有1个变异位点.
由序列比对可知,8个个体的序列共检测到多态位点3个,约占分析位点总数(561 bp)的0.53%,2核苷酸间的变异位点3个,无3核苷酸间的变异位点.因此,3个虎亚种的mtDNA ND4区序列变异主要有转换和颠换2种类型,无缺失/插入、转换与颠换共存的现象.东北虎、孟加拉虎、华南虎和云豹的序列两两碱基替换情况和遗传距离列于表3.
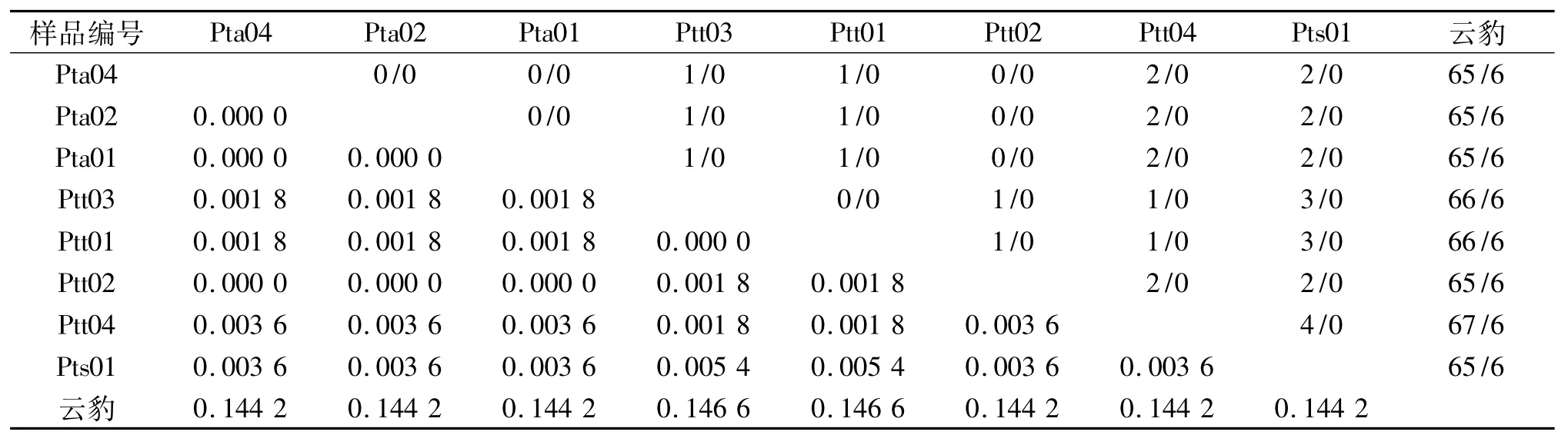
表3 东北虎、孟加拉虎、华南虎和云豹的ND4序列间的转换/颠换(上三角)和遗传距离(下三角)Tab.3 Numbers of transition/transversion(upper-right matrix)and genetic distances(Kimura 2-parameter model,lowerleft matrix)for ND4 sequences of Siberian tiger,Bengal tiger,South China tiger and clouded leopard
从表3可看出:虎亚种间的Kimura 2-parameter遗传距离范围为0.0000~0.0054,变异程度较小;华南虎和孟加拉虎2个个体的遗传距离最大,和其他个体的遗传距离一样;而东北虎个体之间的遗传距离为0.云豹和虎亚种之间的遗传距离范围是0.1442~0.1466,变异程度较小,其中和孟加拉虎2个个体最大,和其他个体的遗传距离一样.
2.3 系统进化树的构建
利用Mega 4.0软件,基于Kimura两参数距离采用邻接法(NJ法)来构建系统进化树,以云豹作为外群比较,同时引进家猫和猎豹的同区域序列作对照.从图1~3中可以看出该拓扑结构分为3个支系,东北虎同孟加拉虎聚在一起,自助置信水平为99%;然后再与华南虎的支系聚合.
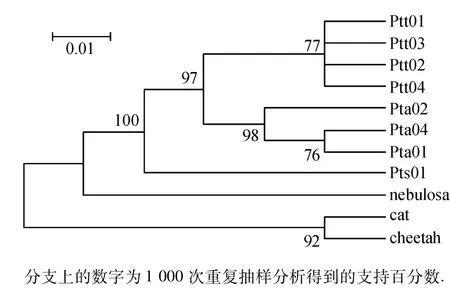
图1 基于COⅠ基因构建的系统进化树Fig.1 Neighbour-joining phylogenetic trees with the Kimura-2 parameter model based on the COⅠgene
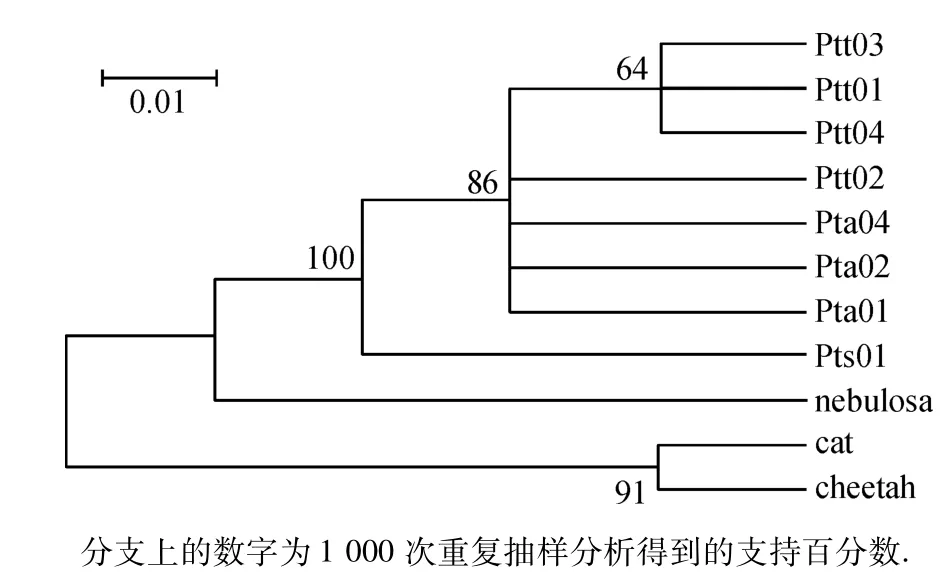
图2 基于COⅢ基因构建的系统进化树Fig.2 Neighbour-joining phylogenetic trees with the Kimura-2 parameter model based on the COⅢgene
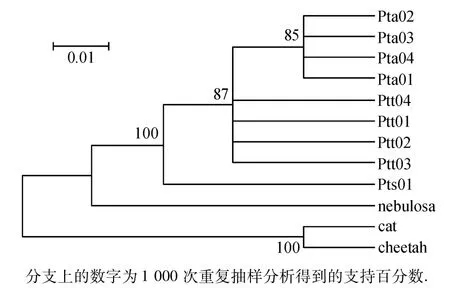
图3 基于ND4基因构建的系统进化树Fig.3 Neighbour-joining phylogenetic trees with the Kimura-2 parameter model based on the ND4 gene
3 讨论
3.1 3个基因的保守性及在进化关系中的应用
在基因序列方面,3个基因序列呈现较多的一致性.3个基因序列都富含A、T,而G的含量都是最少的.张方等[9]认为脊椎动物线粒体基因组遗传密码的使用存在偏倚性,密码子的第3位碱基为A和C的比例明显高于G和T,其中A的比例最高,G最低.
从种内的单倍体个数来看,3个基因单倍体个数都较少,呈现保守性,且种间多态性位点也较少,在位点的突变中也全部是转换和颠换,而且以转换为主,多为无义突变.衡量一个群体mtDNA的遗传多样性的指标有2个:单倍型多样度和核苷酸多样度.这2个指标的值越大,群体的多样性程度越高,遗传多样性越丰富,反之群体的多样性程度越低,遗传多样性越贫乏.因此一方面说明这3个基因比较保守,另一方面也说明3种虎的遗传多样性较低.
从遗传距离上看,不论个体还是亚种间,3个基因呈现的遗传距离均较低,亚种间最高的为COⅠ基因,其最高遗传距离才为 0.0148.Nei[10]认为地理种群之间的遗传距离在0.00~0.05,亚种间的遗传距离约为0.05或更大.然而有许多例外,在有些情况下,亚种间的遗传距离可能与地理种群间的遗传距离一样小.由于亚种分类可能受到人为因素的影响,在分类学实践中亚种形态分化的标准以约75%的个体呈现不同为界限;另一方面,形态学变化和分子歧异之间既存在相关性,又存在独立性[11],在分子水平研究亚种的亲缘关系时,得到的遗传距离可能会大于或小于 Nei[10]估算的亚种间的遗传距离.例如,Ball等[12]对红翅黑鹂 Agelaius phoeniceus的研究和Carr等[13]对维基尼亚鹿 Odocoileus virginianus的研究都表明种群间mtDNA差异程度较小,最大的分别为 0.008 和 0.016.Ashley 等[14]对黑犀牛 Diceros bicormis 2亚种的研究也证实了这一点,2亚种之间的遗传距离仅为0.0029.吴平等[15]利用 RFLP和PCR-RFLP技术研究东北虎和华南虎线粒体DNA多态性,结果显示东北虎和华南虎的遗传距离只有0.00135.
3.2 从3个基因的序列片段构建的进化树看虎的进化关系
本研究结果可以看出华南虎和东北虎、孟加拉虎的遗传距离要大于东北虎和孟加拉虎之间的遗传距离,而以云豹为外群构建的进化树中也可以看出,3个基因序列片段构建的进化树拓扑结构相近,均是东北虎和孟加拉虎先合为一枝,然后与华南虎汇合;且相同的亚种都聚在各自的树枝上,个体间的遗传差异较小,内群的平均遗传距离均小于外群的遗传距离,所以能够很好地区分和3个亚种间的关系.Kimball等[16]研究表明,置信度大于70%的进化枝是比较可信的,而本文构建的基因树除基于COⅢ基因构建的系统进化树有1支为64%外,其余均大于70%,因此构建的基因树是可信的.从COⅠ、COⅢ和ND4基因构建的系统进化树我们可以得出东北虎和孟加拉虎均是由华南虎进化而来的结论.而谭邦杰[17]从地理位置上、张文平[1]从线粒体 DNA D-loop区和ND5基因构建的进化树也均说明了3种虎的进化由来,鉴于现存华南虎的数量及华南虎在虎进化史上的地位,需要我们在华南虎的保护上作出更多的努力.
[1]张文平.华南虎、东北虎、孟加拉虎的 D-loop、ND5、16SrRNA、CytB序列及其在系统进化研究的应用[D].成都:四川大学,2005.
[2]WARREN E,JOHNSON S,O’BRIEN J.Phylogenetic reconstruction of the felidae using 16S rRNA and NADH25 mitochondrial[J].Genes J Mol Evol,1997,44(1):S98-S116.
[3]LUO S J,KIM J H,JOHNSON W E,et al.Phylogeography and genetic ancestry of tigers(Panthera tigris)[J].PLoS Biol,2004,2(12):2275-2293.
[4]陈诗书,汤雪明.医学细胞与分子生物学[M].上海:上海医科大学出版社,1996.
[5]WHITE D J,WOLFF J N,PIERSON M,et al.Revealing the hidden complexities of mtDNA inheritance[J].Mol Ecol,2008,17(23):4925-4942.
[6]郑涛.云豹线粒体DNA全序列及中国13种猫科动物的系统发生关系分析[D].芜湖:安徽师范大学,2006.
[7]张文平,张志和,沈富军,等.华南虎、东北虎、孟加拉虎的D-loop和ND5序列及其在系统进化分析中的应用[J].四川动物,2005,24(1):22-26.
[8]韦鹍.圈养华南虎线粒体基因NADH脱氢酶亚基5序列多态性研究[D].成都:四川大学,2006.
[9]张方,米志勇.动物线粒体DNA的分子生物学研究进展[J].生物工程进展,1998,18(3):25-31.
[10]NEI M.Molecular evolutionary genetics[M].New York:Columbia University Press,1987.
[11]WILSON A C,CARLSON S S,WHITE T J.Biochemical evolution[J].Annu Rev Biochem,1977,46(1):573-639.
[12]BALL R M,FREEMAN S,JAMES F C,et al.Phylogeographic population structure of red-winged blackbirds assessed by mitochondrial DNA[J].Proc Natl Acad Sci USA,1988,85(5):1558-1562.
[13]CARR S M,BALLINGER S W,DERR J N,et al.Mitochondrial DNA analysis of hybridization between sympatric white-tailed deer and mule deer in west texas[J].Proc Natl Acad Sci USA,1986,83(24):9576-9580.
[14]ASHLEY M V,MELNICK D J,WESTERN D.Conservation genetics of the black rhinoceros(Diceros bicornis):I:Evidence from the mitochondrial DNA of three populations[J].Conservation Biology,1990,4(1):71-77.
[15]吴平,周开亚,王亚明.用RFLP和PCR2RFLP技术研究东北虎和华南虎线粒体DNA多态性[J].生物多样性,1997,5(3):173-178.
[16]KIMBALL R T,BRAUN E L,ZWARTJES P W.A molecular phylogeny of the pheasants and part-ridges suggests that these lineages are not monophyletic[J].Mol Phylogenet Evol,1999,11(1):38-54.
[17]谭邦杰.虎在中国的分布[J].中国动物园年刊,1986(1985/1986):165-170.
猜你喜欢
今日畜牧兽医(2022年11期)2022-12-21
汽车与运动(2022年8期)2022-09-16
热带亚热带植物学报(2022年4期)2022-08-18
小星星·阅读100分(低年级)(2022年3期)2022-06-02
小雪花·成长指南(2022年1期)2022-04-09
昆虫学报(2021年11期)2021-12-08
——和田盘羊
野生动物学报(2021年4期)2021-11-14
小哥白尼(野生动物)(2020年1期)2020-06-12
食品与发酵工业(2019年18期)2019-10-11
浙江林业(2018年5期)2018-05-14
