
陆地棉突变系高秆1号的生物学特性
2014-10-11 02:31:56张丽萍狄佳春陈旭升
江苏农业学报 2014年2期
张丽萍, 狄佳春, 赵 亮, 陈旭升
(1.南京农业大学农学院,江苏 南京 210095;2.江苏省农业科学院经济作物研究所/农业部长江下游棉花与油菜重点实验室,江苏 南京 210014)
株高是棉花一个非常重要的农艺性状,与光合强度和丰产潜能等许多因素密切相关,同时也影响植株的株型。增加株高是获得生物产量的重要途径之一[1-3]。同时,增加棉花第一果节位高度有利于机械采收,当棉株第一果枝节位较高,棉朵离地高度大于20 cm,可提高籽棉的采净率[4]。
目前国内外对于高秆突变体的研究主要集中在水稻方面[5-13],其他作物研究甚少。陈旭升等以自主发现的棉花高秆突变体为研究材料,发现高秆突变性状是受1对显性基因控制的质量性状[14]。本研究将进一步分析棉花高秆突变系高秆1号的形态、生理与细胞学等生物学特性,探索植株高秆化机理,旨在为高秆突变体棉花在机械化采收中应用提供理论依据。
1 材料与方法
1.1 供试材料
供试材料为自主选育的棉花高秆突变系高秆1号以及正常株高野生型N089。
1.2 试验方法
1.2.1 表型性状调查 2013年在江苏省农业科学院实验地种植高秆1号与N089,种植方式为营养钵育苗移栽,行距为90 cm,株距为40 cm。从6月19日(蕾期)开始,每隔15 d调查高秆1号及N089的株高、节间长、节间数,于7月19日每隔15 d调查第一果枝节位高,至10月2日(吐絮期)结束,显著性差异分析采用t测验。
1.2.2 激素含量测定 取高秆1号与野生型N089饱满一致的种子,用解剖刀剥除种皮,用分析天平精确称取0.40 g种胚,3次重复,将其置于液氮中固定15 min,放于-70℃冰箱备用。采用酶联免疫法分别测量2个材料种胚中赤霉素(GA3)、生长素(IAA)、脱落酸(ABA)及玉米素(ZR)的含量。显著性差异分析采用t测验。
1.2.3 主茎节间细胞的比较 参照李正理的方法[15],在现蕾期取高秆突变体和野生型植株主茎的倒三节间制作横、纵切片。使用光学显微镜(BX51)在10×放大倍数下观察记录横切细胞的直径、纵切细胞的长度,每处理测量100个细胞,3次重复。观察纵切片同一视野内细胞的数目,每个试验材料随机取3株,每株观察一个切片,每片选取5个不同视野,光学显微镜观察,取平均值,显著性差异分析采用t测验。
2 结果
2.1 高秆突变系株高的发育动态
棉花高秆1号与野生型N089的株高变化动态见图1:从蕾期到吐絮期,高秆1号的株高都极显著地高于野生型(P<0.01)。从变化动态来看,6月19日蕾期到8月18日花期,株高处于快速上升期,且高秆1号的增长速度大于野生型,而后进入一个增长平台期,株高基本不再变化。从最终株高来看,高秆突变体为145.7 cm,而野生型的最终株高仅为79.8 cm,前者的高度是后者的1.82倍。

图1 棉株不同生育时期株高变化动态Fig.1 The heights of cotton Gaogan 1 and wild type N089 at different growth stages
2.1.1 节间长的变化 从图2可见,从棉花的蕾期到吐絮期,高秆1号的节间都极显著长于野生型(P<0.01)。从变化动态看,6月19日到7月4日高秆1号的节间增长速度小于野生型,之后至8月18日其增长速度均大于野生型,8月18日以后高秆1号和野生型的节间增长速度都减缓直至吐絮期停止生长。吐絮期高秆1号的节间长度是野生型的2倍左右。
2.1.2 第一果枝节位高度变化 由图3看出,从7月19日(初花期)开始至10月2日(吐絮期),高秆1号第一果枝节位的高度始终极显著地大于野生型(P<0.01)。从变化动态看,7月19日到8月18日第一果枝节位均处于快速增长期,且高秆1号的增长速度大于野生型;野生型第一果枝节位在8月18日盛花期以后生长减缓直至停止生长,而高秆1号仍保持一定的生长趋势,直至吐絮初期生长减缓直至停止生长。吐絮期高秆1号第一果节位的高度是野生型的4倍多。

图2 棉花不同生育时期节间长度变化动态Fig.2 The lengths of internodes of cotton Gaogan 1 and wild type N089 at different growth stages

图3 棉花不同生育时期第一果枝节位变化动态Fig.3 The heights of the first fruiting branch of cotton Gaogan 1 and wild type N089 at different growth stages
2.1.3 节间数的变化 由图4可以看出,在棉花生育初期,高秆1号的节间数稍多于野生型,而盛花期以后两者的差距消失。从变化动态看,在6月19日到8月18日,突变系的节间数略多于野生型,两者均处于逐步增加状态,并在8月18日达到同一节间数,之后两者均基本停止变化。
通过以上分析可以推测,高秆突变系的株高主要由2个因素决定:一是较高的第一果枝节位高度,二是较长的节间长度。高秆突变系节间数与野生型则无显著差异。
2.2 高秆突变系种胚的激素含量
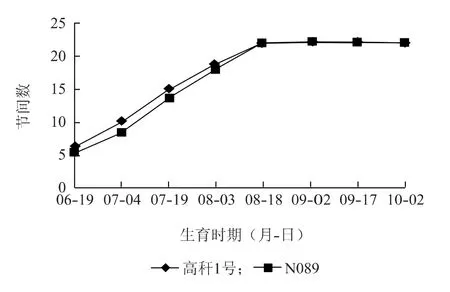
图4 棉花不同生育时期节间数变化动态Fig.4 The number of internodes of cotton Gaogan 1 and wild type N089 at different growth stages
激素是影响株高发育的主要生理因子。取激素合成比较活跃的棉种组织——种胚为材料,利用ELISA法测定4种激素GA3、IAA、ZR和ABA的含量见表1。经过显著性差异分析发现,突变系高秆1号种胚中3种与促进生长有关的激素——GA3、IAA、ZR的含量都极显著高于野生型,前者分别为后者的236.8%、167.7%、279.1%;而ABA含量则显著低于N089,高秆突变体中的含量仅为野生型的76.0%。

表1 不同棉花品种种胚中内源激素含量比较Table 1 The comparison of endogenous hormones in seed embryos of different cotton varieties
2.3 高秆突变体的细胞学观察
植株的高秆突变一般与细胞数目的增加或者细胞长度的增长有关,或是两者共同作用的结果。选择高秆突变体与野生型品种细胞生长最快的倒3节间制备横、纵切片,纵向切片突变体细胞的显微图见图5a、5b。高秆突变体的细胞平均长度为(378.64±23.71)μm,野生型为(263.81±13.89)μm,两者达到极显著差异水平(P<0.01)。同一视野内突变体倒三节间皮层细胞数平均为287.6个,野生型细胞的平均数为250.8个,差异不显著。横向切片对比细胞的直径大小(图5c、5d),突变系细胞直径为(78.04±2.28)μm,野生型的细胞直径为(112.03±5.59)μm,两者差异极显著(P<0.01)。表明,高秆突变体之所以植株高度显著高于野生型品种,是因为其纵向节间细胞长度的明显变长。

图5 高秆突变体与野生型的节间细胞Fig.5 The internode cell of the tall cotton mutant and wild type
3 讨论
3.1 高秆突变体株高形成的主要相关因素
马玉银等[16]研究曾发现,水稻高秆突变体的株高增加,在各个节间和穗长上均有体现。肖辉海等[17]的研究也显示,水稻长穗颈隐性高秆突变体双低培eS的穗颈节间伸长是由幼穗分化后期细胞数增多和细胞长度增加双重作用引起的,其中以后者的作用更显著。本研究表明:在棉花生长的各个时期,高秆1号的株高、节间长、第一果枝节位高度等表型与野生型均具有极显著性差异。通过细胞学观察,在纵向切片中突变体细胞的平均长度明显大于野生型,这说明突变体节间细胞伸长受到了促进。另外,纵向切片观察发现突变体的皮层细胞数明显多于野生型。由此可以推测,棉株纵向节间细胞数目的增多和细胞的伸长,引起了节间伸长,并最终导致突变体植株的高秆化。
3.2 株高发育与激素含量的关系
马良勇等[18]研究发现高秆水稻品种内源生长素、赤霉素类物质显著高于矮秆品种,而脱落酸、乙烯类物质明显低于矮秆品种;汤日圣等[19]研究结果表明13个粳稻品种的赤霉素(GA)和吲哚乙酸(IAA)等内源激素含量远低于高秆品种,矮秆品种GA含量为高秆品种的70% ~80%,而IAA只有高秆品种的20%左右。宋平等[20]认为,半矮秆型的水稻品种的内源ABA含量显著高于高秆型,且水稻株高、苗高与ABA含量呈显著负相关。本研究发现,高秆突变体种胚中 GA3、IAA、ZR含量极显著(P<0.01)高于野生型,ABA含量极显著(P<0.01)低于野生型,与前人研究相符。
3.3 高秆突变体棉花在机械化采收中的应用潜力
随着劳动力价格不断攀升,棉花生产用工量大,与其他作物的比较效益低,中国棉花种植面积正面临不断萎缩的困境。中国棉花的未来之路,在于棉花生产的全程机械化,减少棉花收花用工量,提高植棉综合效益。但机械化收花对棉花株型有比较严格的要求。适合于机采棉的棉花品种株型最好是筒形、木枝少,株型紧凑、疏朗,并且要求第一果枝节位较高,棉朵离地高度至少要大于20 cm,这样可提高籽棉的采净率。如果第一果枝节位高度能达40 cm,则可明显减少土壤以及地膜在机械收花时的夹带,对提高籽棉的品级更加有利。本研究显示高秆突变系高秆1号的第一果枝节位高达38.5 cm,通过杂交转育该性状,选育第一果枝节位高的机采棉,在未来生产中将具有良好的应用前景。
[1] 袁隆平.杂交水稻学[M].北京:中国农业出版社,2002.
[2] 袁隆平.高产优质杂交水稻的选育.杂交水稻,1997,12(16):1-6.
[3] 程式华,翟虎渠.高产优质杂交水稻的育种策略[J].农业现代化研究,2001,21(3):147-150.
[4] 李永宏,李小兵.浅谈推广机采棉技术的成功经验[J].新疆农机化,2009(5):54-55.
[5] OKUNO K,KAWFIT.Gentic analysis of induced long-culm mutants in rice[J].Japan JBreed,1978,28(4):336-342.
[6] RUTGER JN,CARNAHAN H L.A fourth genetic element to facilitate hybrid cereal production-A recessive tall in rice[J].Crop Science,1981,21:373-376.
[7] 廖昌礼,倪克鱼,刘远坤,等.高秆隐性Grlc的遗传与利用研究——Grlc及其测交F1的株高等特征和秆型[J].西南农业学报,1988,1(1):43-46.
[8] 吴世弼,张琦华.水稻诱变获得隐性高秆基因[J].福建农学院学报,1988,3(1):41-45.
[9] 李和标,孙立华,邹江石.水稻隐性高秆广亲和种质02428h的鉴定与研究[J].江苏农业学报,1992,8(3):48-50.
[10] 张 毅,何光华,杨光伟.水稻长穗颈性状的一种新遗传行为的发现与分析[J].中国水稻科学,2004,18(3):213-217.
[11] 陈建民,顾世粱,汤述翥,等.水稻隐性高秆突变的遗传研究[J]. 扬州大学学报,1998,1(3):36-41.
[12] 沈季孟,韩敬花,樊 路,等.一个与高秆基因连锁的太谷核不育硬粒小麦的发现[J].北京农业科学,1997,15(1):9-10.
[13] 李志勇.一个高秆显性四倍体小麦材料的发现与其DELLA蛋白丰度相关性分析[D].郑州:河南农业大学,2012.
[14] 陈旭升,张丽萍,狄佳春,等.陆地棉高秆突变体的质量遗传规律研究[J].江西农业学报,2013,25(3):4-6.
[15] 李正理.植物制片技术[M].2版.北京:科学出版社,1987.
[16] 马玉银,李 磊,李育红,等.一个新的水稻隐性高秆突变体的遗传分析和基因定位[J].中国农业科学,2008,41(12):3967-3973.
[17] 肖辉海.水稻长穗颈隐性高秆突变体穗颈节间的细胞学观察[J]. 西北农林科技大学学报,2008,36(1):131-136.
[18] 马良勇,李西明,朱旭东.水稻株高性状的研究进展[J].福建稻麦科技,2001,19(4):20-23.
[19] 汤日圣,张远海,张金渝.矮秆基因对水稻性状控制的机理探讨[J].中国农业科学,1991,24(2):51-56.
[20] 宋 平,高红胜,谢迎兰.不同籼稻品种的矮生性与内源ABA水平及其结合蛋白的关系[J].西北植物学报,1998,18(3):380-385.
猜你喜欢
工矿自动化(2022年7期)2022-08-13 08:01:04
安徽医科大学学报(2016年12期)2017-01-15 14:21:44
山东农业工程学院学报(2016年6期)2016-12-01 05:38:19
天津职业技术师范大学学报(2016年1期)2016-04-27 02:35:36
农民致富之友·下半月(2015年6期)2015-07-09 10:46:48
农民致富之友(2015年12期)2015-06-11 19:33:04
山东医药(2015年40期)2015-02-28 14:28:45
中国农资(2015年22期)2015-02-01 12:30:07
乡村科技(2014年15期)2014-10-29 07:32:46
哈尔滨商业大学学报(自然科学版)(2014年6期)2014-09-14 04:49:30
