
放牧对若尔盖高寒草甸土壤氮矿化及其温度敏感性的影响
2014-09-19 10:00:48张洪轩王若梦杨满业赵小宁于贵瑞何念鹏
生态学报 2014年15期
赵 宁, 张洪轩, 王若梦, 杨满业, 张 艳, 赵小宁,于贵瑞,何念鹏,*
(1.中国科学院地理科学与资源研究所, 生态系统网络观测与模拟重点实验室, 北京 100101; 2. 中国科学院研究生院, 北京 100049; 3. 四川省草原科学研究院, 成都 611731; 4. 山西农业大学动物科技学院, 太谷 030801; 5. 北京林业大学林学院, 北京 100083)
可利用性氮素是草地生态系统初级生产力的主要限制因子[1]。土壤氮矿化(soil nitrogen mineralization)是生态系统氮循环的重要过程,某种程度决定了氮素有效性[2]。放牧是草地的主要利用方式,对草地氮素循环有重要影响[3]。目前科学家关于放牧与草地土壤氮矿化的研究结论仍存在较大争议,有研究认为放牧促进了土壤氮矿化[4],也有研究认为放牧抑制了土壤氮矿化[5]。Shariff等[6]认为中度放牧下土壤净氮矿化速率最高,通过控制放牧强度能提高土壤有效氮素水平。放牧对土壤氮矿化的影响受到放牧强度、放牧制度、土壤肥力、植被类型等多种因素的共同制约[7-10],是造成不同研究结论存在差异的可能原因。
青藏高原东缘的若尔盖高寒草甸是我国主要牧区之一。独特的地理位置和气候环境使该地区的高寒草甸对外界干扰和气候变化异常敏感[11]。近几十年来,若尔盖高寒草甸的放牧强度持续增加[12],相比其它草地生态系统而言,放牧强度的持续增长对高寒草甸土壤氮矿化乃至氮循环的影响可能更为复杂和不确定。此外,近几十年来该地区气候呈现明显的暖干化趋势[13],由此也可能影响土壤氮矿化过程[14]。放牧和温度升高可能会对若尔盖高寒草甸的土壤氮矿化造成重要的影响,并最终影响到高寒草甸生态系统结构和功能,但迄今为止仍未见相关的研究报道。本研究利用野外放牧实验与室内培养相结合的方法,利用温度敏感性(Q10)来揭示土壤氮矿化对放牧和温度变化的响应趋势,以期为合理地制定高寒草甸放牧制度提供理论支持,并为预测气候变暖对若尔盖高寒草甸氮素循环的影响提供科学依据。
1 研究区域概况与实验方法
1.1 研究区域概况
若尔盖地处青藏高原东缘,属大陆性高原气候。严寒湿润、霜冻期长、四季变化不明显;年平均气温0.7—1.1 ℃,极端最低和最高温度出现在1月和6月,分别为-36 ℃和26 ℃。年均降水量为749.1 mm,年均蒸发量达1262.5 mm[15]。高寒草甸和高寒湿地是该地区主要的景观类型。
1.2 研究方法
野外实验依托于四川省草原科学研究院红原基地的放牧实验样地,该样地位于红原县瓦切乡(样地中心地理坐标:33°06′ N,102°35′ E,海拔3456 m)。实验样地对若尔盖地区的高寒草甸具有较好的代表性。植被为典型的高寒草甸,以嵩草属(Kobresia)、蓼属(Polygonum)植物为主,优势物种有羊茅(FestucaovinaLinn.)、四川嵩草(KobresiasetchwanensisHand.-Mazz)、圆穗蓼(PolygonummacrophyllumD. Don)以及发草(DeschampsiacaespitosaBeauv.)、垂穗披碱草(ElymusnutansGriseb.)等。土壤类型为高山草甸土。
1.2.1 样地设置
放牧实验共设置4种放牧强度,分别为禁牧(CK)、轻度放牧(L)、中度放牧(M)和重度放牧(H),3次重复。每个禁牧实验小区面积1.6 hm2,轻度放牧小区4.2 hm2、中度放牧小区2.5 hm2、重度放牧小区1.9 hm2。放牧实验于2010年开始,除禁牧小区外,各个放牧小区均放牧牦牛3头,放牧强度分别为0、0.7、1.2、1.6牛/ hm2。
2012年4月生长季初期,在每个放牧小区内均匀布置50—60个采集点;在去除地表凋落物后,采用土钻法对0—10 cm土壤样品进行取样,每个小区内所采集的土壤混合形成一个土壤样品(> 5 kg),运回实验室后4 ℃保存。在室内,土壤样品过筛(孔径2 mm)并手工挑除杂质和植物根系后,分为两部分,一部分用于室内培养来测定土壤矿化速率,另一部分自然风干用于测定土壤全碳和全氮等指标。放牧实验样地的基本土壤理化特性如表1所示。

表1 不同放牧强度土壤理化特性
表中数据为平均值,括号内数字代表标准误,n= 3;鉴于处理间各项指标差异均不显著(P> 0.05),表中未用字母标识差异性
1.2.2 实验室培养和化学分析
在测定土壤饱和含水量后,首先称取经过预处理的土壤样品10 g和石英砂5 g,装入清洗干净的50 mL塑料方瓶,摇匀后加蒸馏水调节至60%土壤饱和含水量。随后,将培养样品分别放入5、10、15、20、25 ℃的恒温恒湿培养箱。土壤硝态氮和铵态氮测定分7个批次进行,分别在培养0、7、14、21、28、35、42 d后。培养过程中土壤样品每隔2—3 d补水1次,补水量采用称重法确定,用带蓬头的针管将蒸馏水均匀喷洒在土壤表面使其维持60%土壤饱和含水量。
土壤经50 mL 1 mol/L的KC1浸提后,采用双波长紫外分光光度法测定硝态氮含量[16],靛酚蓝比色法测定铵态氮含量[17]。此外,烘干法测定土壤含水量,电位法测定土壤pH值,干烧法(元素分析仪)测定土壤全碳和全氮含量。
1.2.3 计算和统计分析
(1) 计算方法
土壤净氮矿化积累量由培养前后硝态氮和铵态氮的变化量决定,净氮矿化速率为单位培养时间无机氮含量的变化,可通过以下公式分别计算:
Δt=ti+1-ti
(1)
(2)
(3)
Amin=Aamm+Anit
(4)
Rmin=Amin/Δt
(5)
Rnit=Anit/Δt
(6)

净氮矿化速率与温度之间的关系采用f(T)=A×e(BT)指数模型拟合,式中T为培养温度(℃),A为基质质量指数(substrate quality index),表示温度为0 ℃时土壤净氮矿化的速率,B为温度反应系数。Q10通过下式计算:Q10= exp(10B),即温度每增加10 ℃土壤净氮矿化速率增加的倍数。
(2)统计方法
采用SPSS 11.0统计软件进行数据分析,分别对不同指标 (Rnit,Ramm和Rmin)进行双因素方差分析 (Two-way ANOVA),采用培养7 d时的数据得出放牧和温度的主效应及其交互作用对氮矿化速率的影响,采用Duncan法判断不同放牧强度(CK、L、M、H)和培养温度间的氮矿化速率是否存在显著差异。对培养温度为20 ℃的数据进行单因素方差分析 (One-way ANOVA)分析,探讨不同放牧强度的氮矿化动态。统计差异显著水平为P= 0.05。
2 实验结果
2.1 放牧对氮矿化速率的影响
放牧显著影响了土壤硝化速率和氨化速率(P< 0.001,表2),不同放牧强度间硝化速率存在显著差异(P< 0.05)。4个放牧强度中CK的硝化速率最低 (0.29 mg N kg-1d-1),L的硝化速率最高 (0.42 mg N kg-1d-1),整体趋势为L > M > H > CK(图1)。氨化速率范围介于-0.056—-0.053 mg N kg-1d-1之间,除L的氨化速率显著偏低外(P< 0.05),其他处理间差异不显著。
放牧显著影响土壤净氮矿化速率(P< 0.001,表2),不同放牧强度间净氮矿化速率存在显著差异(P<0.05)。其中,CK的净氮矿化速率最低 (0.23 mg N kg-1d-1),M和H次之,L具有最高的净氮矿化速率(0.34 mg N kg-1d-1)(图1)。L、M和H的净氮矿化速率分别比CK增加了46.5%、35.2%和3.5%。

表2 净氮矿化速率、氨化速率和硝化速率的双因素方差分析结果
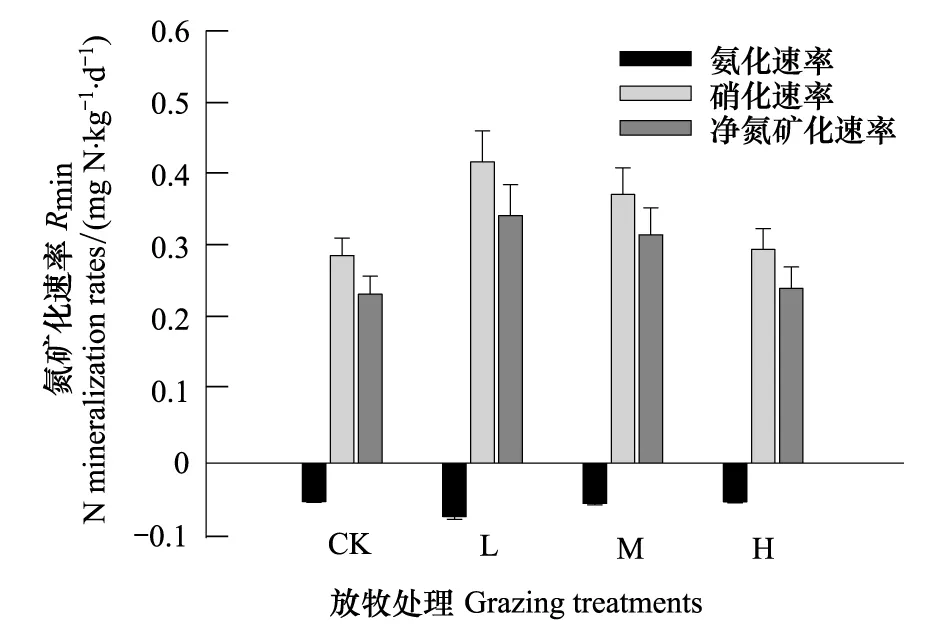
图1 放牧强度对氮矿化速率的影响(平均值 ± 标准误)
2.2 温度对氮矿化速率的影响
温度显著影响了土壤硝化速率(P< 0.001,表2),硝化速率随温度升高呈指数增长形式(图2)。培养温度较低时(5 ℃和10 ℃),温度升高对硝化速率无显著影响(P= 0.71),而培养温度较高时(15,20 ℃和25 ℃),温度升高显著增加了硝化速率(P< 0.05)。然而,温度对土壤氨化速率无显著影响(P> 0.05,表2)。
温度对净氮矿化速率具有显著影响(P< 0.001, 表2),培养温度较低时(5 ℃和10 ℃),不同温度下的净氮矿化速率间无显著差异(P= 0.71),而在较高温度时(15,20 ℃和25 ℃),净氮矿化速率间差异显著(P< 0.05)。由于温度对氨化速率影响较小,因此温度对净氮矿化速率的影响主要取决于硝化速率的变化(图2)。
2.3 土壤氮矿化动态
在培养温度为20 ℃条件下,4个放牧强度的硝态氮积累量随培养时间增长而显著增加(P< 0.001,图3B);然而,不同放牧强度下土壤硝态氮积累量的变化动态不尽相同。CK在6个培养阶段硝态氮积累量都有显著增加(P< 0.05),培养42 d时积累量达到15.51 mg N/kg;L在培养7 d和14 d时硝态氮积累量增加不显著(P> 0.05),之后的4个培养阶段硝态氮都有显著增加(P< 0.05),培养末期积累量达到21.72 mg N/kg;M在培养中期(14,21,28 d)时硝态氮没有显著增加(P> 0.05),培养后期(35 d,42 d)增加显著(P< 0.05),培养末期积累量达到14.62 mg N/kg;H在每个培养阶段硝态氮积累量都有显著增加(P<0.05),培养末期积累量达到17.90 mg N/kg。铵态氮积累量随培养时间增长没有显著变化(P>0.05,图3)。无机氮积累量主要受硝态氮积累的影响,其变化动态和与硝态氮积累量变化动态相似,随培养时间增长显著增加(P< 0.001,图3)。
4个放牧强度的硝化速率和净氮矿化速率随培养时间延长没有显著变化(P> 0.05,图3);在早期培养阶段(0—14 d),4个放牧强度的氨化速率都有显著降低(P< 0.05),在随后的培养过程中氨化速率没有显著变化(P> 0.05,图3)。

图2 温度对净氮矿化和硝化速率的影响

图3 不同放牧强度土壤氮矿化动态
2.4 放牧对土壤净氮矿化速率温度敏感性的影响(Q10)
利用指数模型f(T) =A×e(BT),计算了6个培养阶段不同放牧强度处理下的土壤净氮矿化Q10值(表3),发现放牧强度显著影响Q10(P< 0.001),轻牧和中牧间Q10差异不显著(P= 0.33),而其它放牧强度间Q10存在显著差异(P< 0.05)。4个放牧强度Q10的变化范围在2.01—2.72之间。与禁牧处理相比,放牧样地具有较高的Q10值,总体上呈现随放牧强度增加而增加的趋势,重度放牧样地的土壤具有最高的氮矿化温度敏感性,分别比禁牧、轻度放牧和中度放牧增加了35%、14%和19%。

表3 土壤净氮矿化速率的指数模型参数
表中数值为平均值(标准误),n= 18;A为基质质量指数(substrate quality index),B为温度反应系数,R2为指数方程拟合优度,Q10为土壤氮矿化温度敏感性指数; 不同字母代表显著水平0.05下存在显著差异指数回归模型常数A称为基质质量指数,可以作为衡量土壤有机质质量的指标。通过一元线性回归分析A与Q10变异的关系,如图 4所示A与Q10呈负相关关系,4个放牧强度中A值对Q10变异的解释度可分别为63.4%、43.9%、93.8%和 58.7%。

图4 温度敏感性(Q10)和指数回归模型常数基质质量指数(A)的关系
3 结论与讨论
放牧增加了若尔盖高寒草甸土壤的净氮矿化速率和硝化速率,轻度放牧在各放牧强度中具有最高的氮矿化速率。在其它生态系统也得到类似的结论,例如青藏高原高寒草甸[18]、黄土高原半干旱区草地[19]、内蒙古草原[20]、以及温带盐沼[21]。然而,Andrioli等2010年对阿根廷半干旱草地的研究发现放牧对土壤氮矿化没有显著影响[22]。科学家对放牧强度与土壤氮矿化的关系还存在很大的争议,有研究表明随着放牧强度的增加土壤氮矿化速率增加,重牧时氮矿化速率最高[18-20],也有研究证明中度放牧下净氮矿化速率有最高值[4]。本文得出的轻度放牧下高寒草甸土壤氮矿化速率最高,一方面表明放牧对土壤氮矿化的影响存在多样性,这可能受不同区域植物群落、土壤类型和放牧强度设置等多种因素的影响;另一方面,目前不同研究间关于放牧强度的界定(轻牧、中牧和重牧)的差异也可能是造成上述不一致性的重要原因。
放牧对土壤氮矿化影响的机制目前仍然存在争议。通常认为,轻度放牧造成土壤氮矿化速率升高的主要原因在于轻度放牧下食草动物对植物地上部分的啃食可以刺激根系分泌,从而增加根际微生物的活性,加快氮矿化速率。当放牧强度超过一定阈值后,由于植物地上部分严重受损,造成根系分泌减少,导致氮矿化速率下降[4]。其次,土壤质量也是影响土壤氮矿化的重要因素,肥沃土壤具有较高的氮矿化潜力,相反,贫瘠的土壤氮矿化潜力较低[23]。食草动物的践踏和啃食会对植物群落的物种多样性及功能群产生影响[24-25],从而通过凋落物质量影响土壤质量;食草动物的粪便和尿液的输入也会影响土壤质量。在室内培养状况下, 适宜的温度及土壤含水量为微生物活动提供了适宜环境,因此土壤质量较高时土壤氮矿化速率也相应较高。
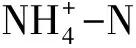
温度敏感性是衡量土壤氮矿化对未来气候变化响应的重要参数[27-28]。大多数据研究均利用Q10来表示土壤氮矿化的温度敏感性,即温度每升高10 ℃土壤氮矿化速率增加的倍数。不同放牧梯度高寒草甸土壤Q10的变化范围在2.01—2.72,与其它研究结论基本相符。例如,Oliver Koch等采用厌氧培养的方法得到的Q10值在1.3—2.8之间[27],高俊琴等研究表明若尔盖湿地沼泽土和泥炭土的净氮矿化速率Q10分别在0. 9—4.5和1.0—4.4之间[29]。由于不同的培养方法(好养培养、厌氧培养)和Q10的计算方法使得不同研究间Q10值在直接比较时受到影响。指数模型参数A可以作为衡量基质质量的指标,被称为基质质量指数[27]。Q10值和参数A呈负相关关系(图4),说明基质质量越差的土壤氮矿化温度敏感性越高。矿化过程是有机质转换为无机质的过程,主要通过微生物介导的酶促反应实现。基质质量越差矿化时所需的酶促反应步骤越多,需要的活化能也越高,因此对温度升高的响应更加剧烈,从而具有较高的Q10值。
放牧显著增加了若尔盖高寒草甸土壤氮矿化的温度敏感性,对高寒草甸土壤氮循环研究具有重要意义。这意味着在未来温度升高的情景下,土壤氮矿化速率增速加快,短期内可以提高植被的氮素可利用性,促进植物快速生长以弥补牲畜取食所造成的氮素流失,维持草地生产力和正常的氮素循环;但从长远来看,它可能会造成土壤氮素的加速流失,最终降低土壤可利用性氮素量,从而对植物生长和生态系统结构和功能形成负反馈。该假说不仅需要在其它生态系统进行验证,还应采用原位培养、稳定性同位素技术和室内培养相结合的方法进行进一步验证;然而,该假说为高寒草甸生态系统的氮素利用和氮素管理提供了新的视角。
:
[1]Vitousek P M, Howarth R W. Nitrogen limitation on land and in the sea: how can it occur? Biogeochemistry, 1991, 13(2): 87-115.
[2]Matson P, Mooney H. Principles of Terrestrial Ecosystem Ecology. New York: Springer Verlag, 2002.
[3]McNaughton S J, Banyikwa F F, McNaughton M M. Promotion of the cycling of diet-enhancing nutrients by African grazers. Science, 1997, 278(5344): 1798-1800.
[4]Xu Y Q, Li L H, Wang Q B, Chen Q S, Cheng W X. The pattern between nitrogen mineralization and grazing intensities in an Inner Mongolian typical steppe. Plant and Soil, 2007, 300(1/2): 289-300.
[5]Accoe F, Boeckx P, Busschaert J, Hofman G, Van Cleemput O. Gross N transformation rates and net N mineralisation rates related to the C and N contents of soil organic matter fractions in grassland soils of different age. Soil Biology and Biochemistry, 2004, 36(12): 2075-2087.
[6]Shariff A R, Biondini M E, Grygiel C E. Grazing intensity effects on litter decomposition and soil nitrogen mineralization. Journal of Range Management, 1994, 47(6): 444-449.
[7]Barger N N, Ojima D S, Belnap J, Shiping W, Chen Z. Changes in plant functional groups, litter quality, and soil carbon and nitrogen mineralization with sheep grazing in an Inner Mongolian grassland. Rangeland Ecology and Management, 2004, 57(6): 613-619.
[8]Augustine D J, McNaughton S J. Interactive effects of ungulate herbivores, soil fertility, and variable rainfall on ecosystem processes in a semi-arid savanna. Ecosystems, 2006, 9(8): 1242-1256.
[9]Singer F J, Schoenecker K A. Do ungulates accelerate or decelerate nitrogen cycling? Forest Ecology and Management, 2003, 181(1/2): 189-204.
[10]Shan Y M, Chen D M, Guan X X, Zheng S X, Chen H J, Wang M J, Bai Y F. Seasonally dependent impacts of grazing on soil nitrogen mineralization and linkages to ecosystem functioning in Inner Mongolia grassland. Soil Biology and Biochemistry, 2011, 43(9): 1943-1954.
[11]Yu B H, Lü C H. Assessment of ecological vulnerability on the Tibetan Plateau. Geographical Research, 2011, 30(12): 2289-2295.
[12]Han D Y, Yang Y X, Yang Y, Li K. Characteristics and influence factors of the swamp degradation under the stress of grazing in the Zoigё Plateau. Acta Ecologica Sinica, 2011, 31(20): 5946-5955.
[13]Dai Y, Luo Y, Wang C K, Shen Y P, Ma Z F, Wang X L. Climate variation and abrupt change in wetland of Zoigё Plateau during 1961—2008. Permafrost and Glacier, 2010, 32(1): 35-42.
[14]Wang C H, Xing X R, Han X G. The effects of temperature and moisture on the soil net nitrogen mineralization in anAneulolepidiumchinensisgrassland, Inner Mongolia, China. Acta Ecologica Sinica, 2004, 24(11): 2472-2476.
[15]Sichuan Province Hongyuan County Annals Compilation Committee. Hongyuan County Annals. Chengdu: Sichuan People′s Publishing House, 1996.
[16]Song G, Sun B, Jiao J Y. Comparison between ultraviolet spectrophotometry and other methods in determination of soil nitrate-N. Journal of Soil, 2007, 44(2): 288-293.
[17]Lu R K. Soil Agro-Chemistrical Analysis. Beijing: China Agricultural Sciences Press, 1999: 107-147.
[18]Chen D D, Sun D S, Zhang S H, Du G Z, Shi X M, Wang X T. Soil N mineralization of an alpine meadow in eastern Qinghai-Tibetan plateau. Acta Agraestia Sinica, 2011, 19(3): 420-424.
[19]Liu T Z, Nan Z B, Hou F J. Grazing intensity effects on soil nitrogen mineralization in semi-arid grassland on the Loess Plateau of northern China. Nutrient Cycling in Agroecosystems, 2011, 91(1): 67-75.
[20]Wu H H, Dannenmann M, Fanselow N, Wolf B, Yao Z S, Wu X, Brüeggemann N, Zheng X H, Han X G, Dittert K, Butterbach-Bahl K. Feedback of grazing on gross rates of N mineralization and inorganic N partitioning in steppe soils of Inner Mongolia. Plant and Soil, 2011, 340(1/2): 127-139.
[21]Olsen Y S, Dausse A, Garbutt A, Ford H, Thomas D N, Jones D L. Cattle grazing drives nitrogen and carbon cycling in a temperate salt marsh. Soil Biology and Biochemistry, 2011, 43(3): 531-541.
[22]Andrioli R J, Distel R A, Didoné N G. Influence of cattle grazing on nitrogen cycling in soils beneathStipatenuis, native to central Argentina. Journal of Arid Environments, 2010, 74(3): 419-422.
[23]Deenik J. Nitrogen mineralization potential in important agricultural soils of Hawai′i. Soil Crop Management, 2006, 15(1): 1-5.
[24]Altesor A, Oesterheld M, Leoni E, Lezama F, Rodriguez C. Effect of grazing on community structure and productivity of a Uruguayan grassland. Plant Ecology, 2005, 179(1): 83-91.
[25]Anderson P M L, Hoffman M. The impacts of sustained heavy grazing on plant diversity and composition in lowland and upland habitats across the Kamiesberg mountain range in the Succulent Karoo, South Africa. Journal of Arid Environments, 2007, 70(4): 686-700.
[26]Sierra J. Temperature and soil moisture dependence of N mineralization in intact soil cores. Soil Biology and Biochemistry, 1997, 29(9/10): 1557-1563.
[27]Koch O, Tscherko D, Kandeler E. Temperature sensitivity of microbial respiration, nitrogen mineralization, and potential soil enzyme activities in organic alpine soils. Global Biogeochemical Cycles, 2007, 21(4): GB4017.
[28]Wang C H, Wan S Q, Xing X R, Zhang L, Han X G. Temperature and soil moisture interactively affected soil net N mineralization in temperate grassland in Northern China. Soil Biology and Biochemistry, 2006, 38(5): 1101-1110.
[29]Gao J Q, Ouyang H, Zhang F, Wang C M. The response of soil nitrogen mineralization to soil temperature and soil moisture in Zoigё alpine wetland. Wetland Science, 2008, 6(2): 229-234.
参考文献:
[11]于伯华, 吕昌河. 青藏高原高寒区生态脆弱性评价. 地理研究, 2011, 30(12): 2289-2295.
[12]韩大勇, 杨永兴, 杨杨, 李珂. 放牧干扰下若尔盖高原沼泽湿地植被种类组成及演替模式. 生态学报, 2011, 31(20): 5946-5955.
[13]戴洋, 罗勇, 王长科, 沈永平, 马振峰, 王小兰. 1961—2008 年若尔盖高原湿地的气候变化和突变分析. 冰川冻土, 2010, 32(1): 35-42.
[14]王常慧, 邢雪荣, 韩兴国. 温度和湿度对我国内蒙古羊草草原土壤净氮矿化的影响. 生态学报, 2004, 24(11): 2472-2476.
[15]四川省红原县志编撰委员会. 红原县志. 成都: 四川人民出版社, 1996.
[16]宋歌, 孙波, 教剑英. 测定土壤硝态氮的紫外分光光度法与其他方法的比较. 土壤学报, 2007, 44(2): 288-293.
[17]鲁如坤. 土壤农业化学分析方法. 北京: 中国农业科技出版社, 1999: 107-147.
[18]陈懂懂, 孙大帅, 张世虎, 杜国祯, 史小明, 王向涛. 青藏高原东缘高寒草甸土壤氮矿化初探. 草地学报, 2011, 19(3): 420-424.
[29]高俊琴, 欧阳华, 张锋, 王春梅. 若尔盖高寒湿地土壤氮矿化对温度和湿度的响应. 湿地科学, 2008, 6(2): 229-234.
猜你喜欢
儿童故事画报·自然探秘(2024年5期)2024-05-22 22:26:58
疯狂英语·新读写(2023年4期)2023-05-10 10:44:22
北京大学学报(自然科学版)(2022年4期)2022-08-18 06:57:34
青海草业(2022年2期)2022-07-23 09:34:58
草业学报(2021年9期)2021-09-22 08:55:16
成都信息工程大学学报(2021年1期)2021-07-22 07:21:50
活力(2019年21期)2019-04-01 12:17:10
青年文学家(2016年34期)2017-03-31 18:32:36
农业环境科学学报(2017年2期)2017-03-20 14:57:30
中国诗歌(2016年7期)2016-12-05 20:43:22
