
天祝高寒草甸高原鼢鼠鼠丘植被演替调查
2014-09-11 06:19周建伟花立民王巧玲王贵珍
草原与草坪 2014年3期
周建伟,花立民,王巧玲,刘 丽,王贵珍
(甘肃农业大学 草业学院/草业生态系统教育部重点实验室/甘肃省草业工程实验室/中-美草地畜牧业可持续发展研究中心,甘肃 兰州 730070)
青藏高原高寒草甸生态系统的复杂性,为多种啮齿动物提供了必要的栖息场所[1]。高原鼢鼠(Myospalaxbaileyi)是青藏高原高寒草甸生态系统中主要的啮齿动物,也是高寒草甸的主要鼠害之一,其种群数量变化对高寒草甸生态系统的结构和功能有着深刻的影响[2,3]。由于高原鼢鼠在地下生活,其挖掘、采食等活动对天然草地造成不同程度的破坏和干扰[4-6]。干扰是影响草地植物群落多样性和植物种类组成的重要因素,高原鼢鼠的造丘活动破坏了草地原有的植物群落,但其土丘却为种子附着、植被更新提供了空间[7-9]。因此,研究鼠丘植物群落的演替及植物多样性变化,对研究高寒草地生态系统有重要意义[10-15]。试验以祁连山东段高寒草甸为背景,以高原鼢鼠土丘植被为研究对象,研究植被演替过程中植物多样性和生活型变化的结构和趋势,旨在为高寒草甸生物多样性的保护提供科学依据。
1 研究区概况与研究方法
1.1 研究区概况
研究地点设在天祝县甘肃农业大学高山草原试验站,位于东祁连山的天祝金强河河谷,南北宽5~15 km,东西长30 km。境内地形受马牙雪山和雷公山强烈隆起的影响,形成东西走向的峡谷地带,西高东低。天然草地主要为高寒草原和高寒草甸。地理坐标为N 37°10′~37°14′,E 102°40′~102°49′,海拔2 710~3 880 m,气候寒冷潮湿,太阳辐射强。年均温-0.1 ℃,1 月平均温度-18.3 ℃,7月平均温度12.7 ℃,>0 ℃年积温1 380 ℃;年降水量416 mm,多为地形雨,集中于7、8、9 月。无绝对无霜期,仅分冷热两季。天然草原主要为高寒草原和高寒草甸。主要植物有垂穗披碱草(Elymusdahuricus)、矮嵩草(Kobresiahumilis)、线叶嵩草(Kobresiacapillifoli)、二裂委陵菜(Potentillabifurca)、秦艽(Gentianamacrophylla)、扁蓿豆(Ruthenianmedic)、早熟禾(Poaceaeannua)、狗娃花(Heteropappushispidus)、黄芪(Astragalusmembranaceus)、棘豆(Oxytropisbella)等。啮齿动物组成包括高原鼢鼠、达乌尔黄鼠(Spermophilusdauricus)、根田鼠(Microtuslimnophylus)、五趾跳鼠(Allactagasibirica)、狭颅鼠兔(Ochotonathomasi)等10种,高原鼢鼠是该地区的优势鼠种,也是最主要的草原害鼠,对草地植被破坏非常严重,有些区域破坏率甚至能达到50%以上[16]。
1.2 研究方法
1.2.1 样地设置 2011~2013年,在研究地选择高原鼢鼠典型分布区,以定点标记结合鼠丘年龄划分[17]的方法,在放牧强度、植被一致的同一块天然草地选择不同年限形成的鼠丘,按形成时间划分为5个演替阶段,即阶段Ⅰ:当年;阶段Ⅱ:1年;阶段Ⅲ:2年;阶段Ⅳ:3年;阶段Ⅴ:4年。前3个阶段采用定点标记的方法确定,即以每年6月新形成的鼠丘用木桩标记为准,阶段Ⅳ和阶段Ⅴ利用鼠丘年龄划分法,即在2011年分别以鼠丘植被盖度在20%和60%以下确定两个阶段的鼠丘,同样在6月用木桩标记。于2013年8月在每个阶段的标记鼠丘设置5个重复,以0.5 m×0.5 m样方法测定各鼠丘上各植物的盖度、生物量,同时在鼠丘附近选择同样大小的自然植被样方作为对照,每组样方重复5次。
1.2.2 植物功能群划分及多样性计算 根据高寒草甸群落组成的特点,按照植物的生活型划分为4类群:(1)1~2年生植物;(2)莎草科植物;(3)多年生禾草;(4)多年生杂类草。多样性指标选用物种数(S)、Mar galef丰富度指数(O)、Shannon-Wiener 指数(H′)和Pielou均匀度指数(E)4 个指标。
1.2.3 数据处理 生物多样性计算采用孔凡洲等[18]在Excel软件中制作的生物多样性指数计算方法处理。方差分析和制图由SPSS 17.0软件和Excel 2003软件完成。
2 结果与分析
2.1 鼠丘植物群落外貌特征
植物群落组成受环境异质性的影响,在局部环境均表现出不同的外貌特征。由图1可以看出,不同的演替阶段高原鼢鼠鼠丘植物群落的总盖度差异显著(P<0.05)。随演替的进行,鼠丘植被总盖度呈递增趋势,但仍与原生植被差异显著(P<0.05)。
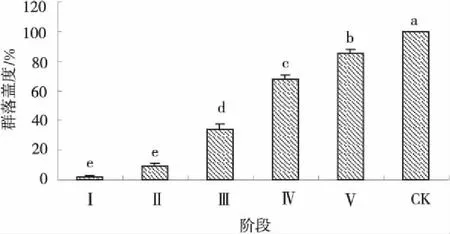
图1 5个演替阶段鼠丘植物群落盖度变化

图2 5个演替阶段鼠丘功能群植物群落特征
各生活型类群植物演替表明(图2),随鼠丘植被演替年限的增加,1~2年生植物盖度呈递增趋势,且第4年的盖度61.4%显著高于其他阶段及原生植被13.3%。杂类草也表现出同样的递增趋势,但是各阶段都低于原生植被。多年生禾草及莎草科植物盖度在各阶段群落中的比例较小(<5%),且都显著低于原生植被,且各阶段变化差异不显著。由表1可知,与生活型类群盖度变化不同,1~2年生植物生物量比例在阶段Ⅰ~Ⅳ呈显著递减,而在阶段Ⅴ比例又开始上升,但各阶段比例都显著高于原生植被。杂类草生物量变化却表现出与1~2年生植物相反的变化趋势,但在达到4年的鼠丘群落的生物量比例与原生植被接近,分别为44.66%和41.86%。多年生禾草及莎草科植物在各生活型类群中所占比例仍然相对较低。

表1 5个演替阶段鼠丘植物生物量在群落中的比例
2.2 植物物种多样性分析
物种多样性是衡量群落结构与功能的指标。物种多样性可以很好反应出群落的组成、变化及发展[19]。鼠丘群落中植物的多样性指数在不同阶段表现出一定的差异性,原生植被(对照)物种丰富度指数均高于不同演替阶段鼠丘群落。随着演替的进行,鼠丘群落物种丰富度呈增加趋势,顺序为原生植被>阶段Ⅴ>阶段Ⅳ>阶段Ⅲ>阶段Ⅱ>阶段Ⅰ,且阶段Ⅴ显著高于其他各阶段。Pielou均匀度指数与物种丰富度指数相互独立,在群落演替的早期,其均匀度一般相对较低,但是研究中发现从阶段2到原生植被未表现出均匀度递增的趋势。Shamon-wiener指数在演替阶段呈先增后降低的趋势,且形成3年以上的鼠丘显著高于1~2年(表2)。
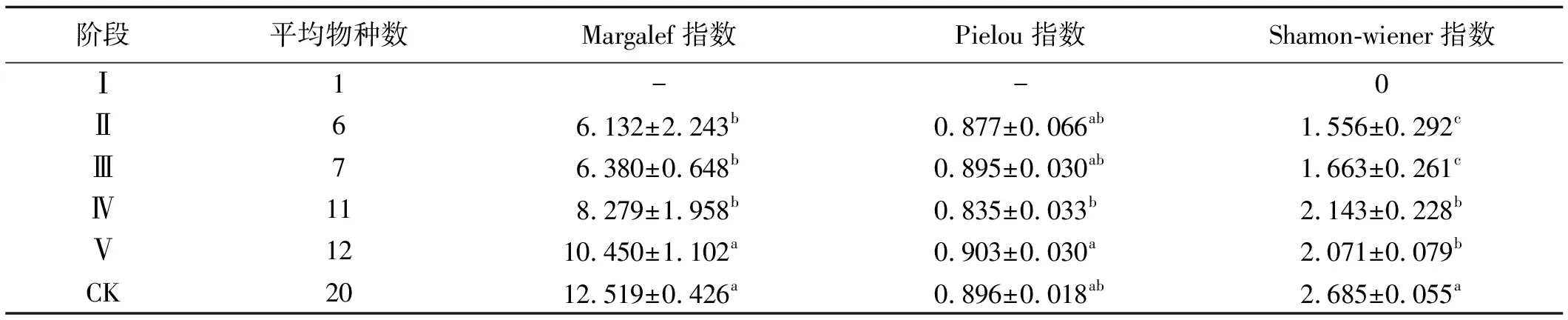
表2 5个演替阶段鼠丘植物多样性比较
注:同列不同小写字母表示差异显著(P<0.05)
2.3 植物生活型
植物生活型是指不同种类的植物对相似环境的趋同适应而在形态、结构、生理、尤其是外貌上所反映出来的植物类型[20]。从各阶段鼠丘植物的物种分布可以看出(表3),阶段Ⅰ基本没有植物覆盖,只有零星分布的一年生杂类草灰绿藜(Chenopodiumglaucum)。从阶段Ⅱ开始,1年生、2年生及多年生杂类草等演替初期物种,如灰绿藜、早熟禾、扁蕾(Gentianopsisbarbata)、香薷(Elsholtziaciliata)、黄鹌菜(Youngiajaponica)、角茴香(Hypecoumerectum)、繁缕(Stellariamedia)、二裂委陵菜等迅速入侵,在植物群落中占据优势地位,但是群落总盖度相对较低。随着鼠丘形成时间的推移,鼠丘土壤变得紧实,能够适应紧实土壤的具备较强根茎繁殖能力及直根系的物种在空间竞争中获得优势,成为群落中的优势种,有的甚至成为建植种,如二裂委陵菜、蒲公英(Taraxacummongolicum)、黄芪(Astragalusmembranaceus)等多年生杂类草相继出现。同时群落中出现了一些高寒草甸的代表植物,如垂穗披碱草、矮嵩草、线叶嵩草(Kobresiacapillifolia)、针茅(Stipacapillata)等,但这些植物在群落中的盖度较低。在演替各阶段1、2年生植物物种数未发生变化,物种组成较为接近,多年生禾草及莎草物种变化较大。阶段Ⅰ~Ⅳ多年生杂类草物种数量及组成也较为相似,但在形成4年的鼠丘群落中多年生杂类草数量却显著增加,物种组成与原生植被接近(表4)。

表3 5个演替阶段鼠丘植物群落组成

表4 5个演替阶段鼠丘物种数量分布
3 讨论与结论
高原鼢鼠鼠丘植被恢复演替的研究国内开展较多[10-15,17,25],张卫国等[8]对不同放牧条件下鼠丘植被恢复进行了研究,江小蕾等[12]对甘南玛曲不同演替阶段高原鼢鼠土丘植物群落的多样性进行了研究,结果表明,随着演替的进行,物种丰富度呈累积递增趋势,物种多样性指数在阶段Ⅳ达到峰值,阶段Ⅴ略有降低,这与Odum[21]对鼠丘植被恢复演替模型预测一致,即在鼠丘形成4年达到演替后期。这与江小蕾[12]的研究略有不同,其认为鼠丘植被在8~10年达到演替后期,可能是各自研究区植被物种组成、放牧强度、土壤结构及气候等因素不同而造成差异。而何俊龄、张黎敏等[22,23]通过土壤种子库与鼢鼠鼠丘植被恢复关系的研究推测鼠丘植被恢复周期在4~5年。以上结果都说明鼠丘植物群落的演替是一个快速恢复过程。
植物群落的生活型类群组成能综合反映草地生态环境对植物的影响。在不同阶段的群落中,随着演替的进行,鼠丘植被群落盖度显著增加,同时各阶段植被表现出不同群落外貌特征。从不同阶段鼠丘植被各种生活型类群占有的比例可以看出,当年形成的鼠丘基本上处于全裸露状态。越年形成的鼠丘上,1、2年生植物占据了主要地位,随着鼠丘植被演替,1、2年生及多年生杂类草共同构成鼠丘群落优势种,演替初期它们大多数属于相对的所谓“机会种”,如香薷、兰石草、西伯利亚蓼、黄鹌菜等[24],在高原鼢鼠种群密度区,此类植物成为群落优势种[25]。在演替的各阶段,鼠丘群落物种组成伴有一定的变异性,可能是受现有植被及土壤种子库的影响[26],也可能受外界物种的入侵的影响[27]。在所有的群落中,莎草科植物和多年生禾本科植物所占比例都比较低,分别在0.1%~3.1%和2.3%~5.5%。除1年鼠丘外,其余4个群落中多年生杂类草占相对优势。高寒草甸的优势种如矮嵩草、线叶嵩草等在鼠丘植物群落中却未能占据优势地位,而杂类草长期占有竞争优势,受放牧及鼢鼠对鼠丘的二次利用等因素影响,短时间内不能完成顶级演替,有些鼠丘植被甚至开始新一轮的演替,这也是高原鼢鼠分布区草地退化的一个标志。随着高原鼢鼠的造丘活动,不同时期鼠丘特征各异,植物生活型功能群也发生变化。同时,鼠丘植被的恢复受环境因子的影响,如土壤微生物、水分等,研究中应考虑这些因素的影响。
在天祝高寒草甸高原鼢鼠鼠丘植被演替过程中,植被迅速恢复,物种组成趋于原生植被,但外貌特征差异较大;1~2年生及多年生杂类草在群落中占据优势地位,同时伴有外来物种的入侵;多年生禾草与莎草科植物在竞争中处于劣势。
参考文献:
[1] 韩天虎,张卫国,丁连生.高原鼢鼠栖息地的植被特征[J].草业学报,1999,8(2):43-49.
[2] 张堰铭,刘季科.地下鼠生物学特征及其在生态系统中的作用[J].兽类学报,2002,22(5):144-153.
[3] Zhang Yanming,Zhang Zhibin,Liu Jike.Burrowing rodents as ecosystem engineers:the ecology and management of plateau zokors (Myospalaxfontanierii) in alpine meadow ecosystems on the Tibetan Plateau[J].Mammal Review,2003,33(3-4):284-294.
[4] 张宏利,卜书海,韩崇选,等.鼠害及其防治方法研究进展[J].西北农林科技大学学报(自然科学版),2003,31:167-171.
[5] 王宝山,尕玛加,张玉.青藏高原“黑土滩”退化高寒草甸草原的形成机制和治理方法的研究进展[J].草原与草坪,2007(2):72-77.
[6] 王权业,樊乃昌.高原鼢鼠的挖掘活动及其种群数量统计方法探讨[J].兽类学报,1987,7(4):283-290.
[7] 李春鸣,张凯,徐长林,等.高寒草甸鼢鼠新旧鼠丘中种子数量特征[J].草原与草坪,2011(6):17-26.
[8] 张卫国,江小雷.鼢鼠的造丘活动及不同休牧方式对草地植被生产力的影响[J].西北植物学报,2004,24(10):1882-1887.
[9] 魏万红,王权业,周文扬,等.捕食干扰对Myospalaxbaileyi种群繁殖力的影响[J].生态学报,1998,18(2):176-180.
[10] Spence S R.Influence of pocket gopher mounds on a Texas coastal prairie[J].Oecologia,1985,66:111-115.
[11] 何俊龄,张金沙,杨莹博,等.高原鼢鼠土丘空间格局及主要特征研究[J].草业学报,2006,15(1):107-112.
[12] 江小雷,张卫国,杨振宇,等.不同演替阶段鼢鼠土丘群落植物多样性变化研究[J].应用生态学报,2004,15(5):814-818.
[13] Tilman D.Plant succession and gopher disturbance along an experimental gradient[J].Oecologia,1988,60:285-292.
[14] 付伟,赵俊权,杜国祯,等.青藏高原高寒草地放牧生态系统可持续发展研究[J].草原与草坪,2013(1):84-88.
[15] 杜国祯,王刚.甘南亚高山草甸人工草的演替和质量变化[J].植物学报,1995,37(4):306-313.
[16] 胡发成,段军红,向金成,等.高寒草甸草地高原鼢鼠的防治指标研究[J].畜牧兽医杂志,2012,31(6):6-7.
[17] 辛小娟,杨莹博,任正炜,等.鼢鼠土丘植物群落演替生态位动态及草地质量指数[J].生态学杂志,2011,30(4):700-706.
[18] 孔凡洲,于仁成,徐子钧,等.应用Excel软件计算生物多样性指数[J].海洋科学,2012,36(4):57-62.
[19] Loreau W.Biodiversity and ecosystem functioning:Recent theoretical advances[J].Oikos,2000,91:3-17.
[20] 任青山.西藏冷杉原始林群落物种多样性初步研究[J].生态学杂志,2002,21(2):67-70.
[21] Odum E P.The strategy of ecosystem development[J].Science,1969,164:262-270.
[22] 何俊龄.土壤种子库与鼢鼠土丘植被恢复的关系[D].兰州:兰州大学,2007.
[23] 张黎敏,龙瑞军,汪永红,等.土壤筛对一定深度不同退化高寒草甸土壤种子库的分选效果[J].草原与草坪,2005(5):46-49.
[24] 王刚,杜国祯.鼢鼠土丘植被演替过程中的种的生态位分析[J].生态学杂志,1990(1):1-6.
[25] 张堰铭.高原鼢鼠对高寒草甸群落特征及演替的影响[J].动物学研究,1999,20(6):435-440.
[26] Rogers W,Hartnett D.Temporal vegetation dynamics and recolonization mechanisms on different-sized soil disturbances in tallgrass prairie[J].American Journal of Botany,2001,88(9):1634-1642.
[27] Pakeman R J,Small J L.The role of the seed bank,seed rain and the timing of disturbance in gap regeneration[J].Journal of Vegetation Science,2005,16:121-130.
猜你喜欢
成都信息工程大学学报(2022年4期)2022-11-18
青海草业(2022年2期)2022-07-23
Zoological Research(2022年3期)2022-06-07
南方农业(2021年30期)2021-12-25
兽医导刊(2020年3期)2020-12-30
家教世界·创新阅读(2020年4期)2020-06-03
当代工人·精品C(2020年1期)2020-05-20
家教世界·创新阅读(2020年1期)2020-05-11
家教世界·创新阅读(2020年2期)2020-04-07
活力(2019年21期)2019-04-01
