
白藜芦醇对视网膜色素上皮细胞增殖的影响*
2014-08-13 12:06王雯婕刘小勇曲艺欣袁晟辰郝晓宁
中国病理生理杂志 2014年10期
王雯婕, 陈 剑, 刘小勇, 曲艺欣, 周 清, 袁晟辰, 郝晓宁
(暨南大学附属第一医院眼科,广东广州 510632)
增生性玻璃体视网膜病变(proliferativevitreoretinopathy,PVR)是视网膜脱离的严重并发症也是视网膜脱离复位手术失败的常见原因[1]。PVR是指移行到脱离的视网膜表面和下方及脱离的玻璃体后表面的视网膜色素上皮细胞(retinalpigmentepithelium,RPE)和胶质细胞增殖形成膜,膜的收缩导致视网膜皱缩、固定及脱离的一种病变。PVR是常见的难治性致盲性眼病之一,约5%~10%的孔源性视网膜脱离发生PVR[2-3]。
目前,临床上治疗PVR的基本方法是玻璃体切割手术,尽管手术设备和技术都有很大提高,但是术后PVR复发仍很常见[4]。由于PVR是由细胞增生和收缩引起的病变,因此药物治疗抑制增殖膜的形成是治疗的关键,主要有抗代谢药、皮质类固醇、维生素及其衍生物等[5],而天然植物源性药物的研究目前也受到重视[6]。
白藜芦醇(resveratrol,Res)是天然多酚类化合物,主要来源于葡萄、虎杖等植物。Res是一种生物性很强的天然抗氧化剂,有抗氧化、抗炎、降低血小板聚集、抗增殖和抗肿瘤等作用[7-8]。本文以视网膜色素上皮细胞株ARPE-19为研究对象,探讨Res对ARPE-19细胞增殖的影响及其作用机制,为其用于PVR的治疗和术后预防复发提供资料。
材 料 和 方 法
1 细胞、试剂和仪器
人视网膜色素上皮细胞株ARPE-19由中山眼科中心惠赠。
Res、胰蛋白酶干粉和EDTA(Sigma);CCK-8试剂盒(Dojindo);DMEM/F12和胎牛血清(FBS)(HyClone);小鼠抗人增殖细胞核抗原(proliferating cell nuclear antigen,PCNA)(Proteintech);RNA提取试剂盒、cDNA逆转录试剂盒和SYBR荧光定量PCR试剂盒(TaKaRa)。倒置显微镜(Leica);酶标仪(Safire);FACSAria流式细胞仪(BD);激光共聚焦显微镜510(Zeiss)。
2 方法
2.1细胞培养及传代 将ARPE-19细胞置于含10%胎牛血清和1%青霉素-链霉素的DMEM/F12培养液中,并于37 ℃、5% CO2、饱和湿度的恒温培养箱中培养,每2~3 d换液1次,待细胞生长至基本汇满培养瓶底时,用0.25%胰酶和0.02% EDTA消化,按1∶2~1∶3传代培养。
2.2CCK-8检测Res对ARPE-19细胞增殖的影响 将ARPE-19以1×104cells/well接种于96孔板,待细胞贴壁后,用含0、50、100、150、200和300μmol/LRes的培养基分别培养24 、48 和72h,0μmol/LRes处理组为对照组,吸去上清,各孔加入含10%CCK-8的培养基,培养箱孵育2.5h后酶标仪测450nm处吸光度(A)值。细胞增殖指数(proliferationindex)=(实验组A值-空白组A值)/(对照组A值-空白组A值)×100%,空白组为不含细胞的培养基。每组5个复孔,实验重复3次。
2.3流式细胞术检测Res对ARPE-19细胞周期分布和凋亡的影响 将ARPE-19细胞以2×105cells/well接种于6孔板中,待细胞贴壁后用0、100、150及200 μmol/L Res分别处理细胞48 h,0 μmol/L Res处理组为对照组。胰蛋白酶消化收集细胞,加入70%冰乙醇固定过夜,离心后弃上清,PBS洗涤两遍,加入200 μL碘化丙啶(propidium iodide, PI)染色,37 ℃避光孵育15 min,过滤后上流式细胞仪检测细胞周期各期分布。收集各组细胞悬液,离心弃上清,200 μL binding buffer重悬,5 μL Annexin V-FITC染色孵育10 min,离心弃上清,200 μL binding buffer重悬,10 μL PI染色,上流式细胞仪检测各组细胞凋亡率。
2.4免疫荧光检测Res对ARPE-19表达PCNA的影响 生长状况较好的ARPE-19消化制备细胞悬液,接种2滴细胞悬液于装有盖玻片的6孔板中,2h后待细胞贴壁后每孔加入含0μmol/L、150μmol/LRes的培养基2mL,培养48h,PBS冲洗后多聚甲醛室温固定20min,3g/L的TritonX-100破膜30min,5%驴血清封闭1h,加入PCNAⅠ抗4 ℃孵育过夜,洗去孵育液,加入Cy3荧光Ⅱ抗,室温孵育40min,洗去孵育液,加DAPI核染,室温孵育30min,洗去孵育液,滴抗荧光淬灭剂封片,4 ℃冰箱避光保存,共聚焦显微镜下观察拍照。
2.5Real-time PCR检测PCNA、P21和P27 mRNA的表达 细胞分组同流式检测,按Trizol试剂盒说明书提取细胞总RNA,用逆转录试剂盒进行逆转录合成cDNA,以逆转录产物为模板进行荧光实时定量PCR。反应体系为20 μL:2×SYBR®Premix Ex TaqTMⅡ 10 μL,上、下游引物各0.6 μL,cDNA 2 μL,dddH2O 6.8 μL。引物序列设计如下:PCNA的上游引物为5’-AGACTTTCCTCCTTCCCGCCTG-3’,下游引物为5’-AGGTCCTTGAGTGCCTCCAACA-3’;P21的上游引物为5’-TCCAGCGACCTTCCTCATCCA-3’,下游引物为5’-TCCATAGCCTCTACTGCCACCA-3’;P27的上游引物为5’-GGACGATCCTCCTCCAAGACAA-3’,下游引物为5’-CTGGGCATTCAGAGCGGGATTA-3’;β-actin的上游引物为5’-AGCAGATGTGGATCAGCAAG-3’,下游引物为5’-AAGGGTGTAACGCAGCAAAG-3’。反应条件为:95 ℃ 3 min;95 ℃ 30 s,60 ℃ 30 s,45个循环。结果用相对定量分析方法2-ΔΔCt分析。以β-actin为内参照计算PCNA、P21和P27的mRNA相对表达量。
3 统计学处理
采用SPSS 13.0 软件作统计处理,计量资料用均数±标准差(mean±SD)表示,组间差异比较用单因素方差分析,两两比较采用SNK-q检验,以P<0.05 为差异有统计学意义。
结 果
1 倒置显微镜观察ARPE-19细胞生物学特性
传代后ARPE-19细胞24 h内已贴壁, 细胞逐渐由圆形伸展呈多形性,多呈纺锤形、短梭形及三角形等, 可见黑色素颗粒围绕透明的细胞核周围, 呈集落生长;4~5 d细胞生长开始融合,见图1。

Figure 1. The morphology of ARPE-19 cells observed under inverted microscope (×50). A: 24 h after passage; B: 96 h after passage.
2 Res对ARPE-19细胞增殖的影响
Res对RPE增殖有一定的抑制作用,且随着浓度和作用时间增加,其抑制作用增强,见表1。虽处理24 h时Res各浓度组与对照组有显著差异(P<0.05),但50~200 μmol/L组增殖率都大于60%,不能有效抑制细胞增殖;处理48 和72 h能有效抑制细胞增殖,各浓度组与对照组有统计学差异(P<0.05),因此为使细胞达到半数抑制且不增加作用时间,处理48 h是最佳抑制时间,150 μmol/L是最佳浓度,见图2。
表1不同浓度Res作用ARPE-19细胞24h、48h、72h后CCK-8结果
Table 1. TheA450 values of ARPE-19 cells treated with resveratrol at different doses for 24 h, 48 h and 72 h (Mean±SD.n=15)
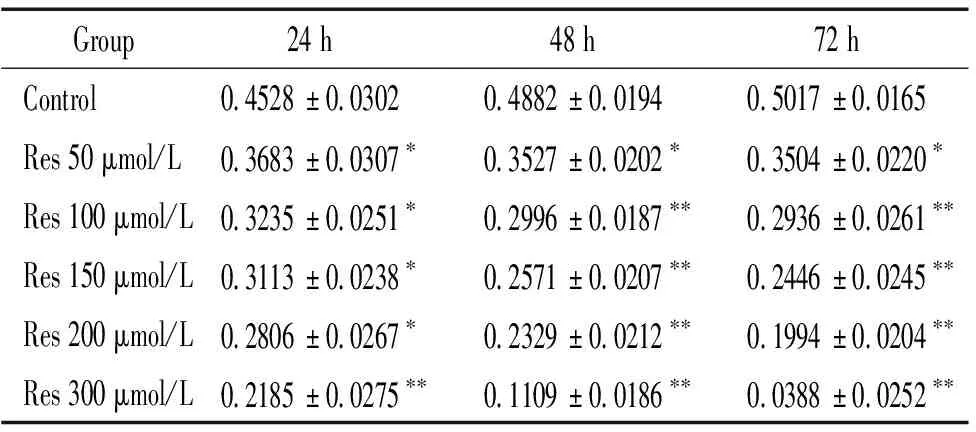
Group24 h48 h72 hControl0.4528±0.03020.4882±0.01940.5017±0.0165Res 50 μmol/L0.3683±0.0307∗0.3527±0.0202∗0.3504±0.0220∗Res 100 μmol/L0.3235±0.0251∗0.2996±0.0187∗∗0.2936±0.0261∗∗Res 150 μmol/L0.3113±0.0238∗0.2571±0.0207∗∗0.2446±0.0245∗∗Res 200 μmol/L0.2806±0.0267∗0.2329±0.0212∗∗0.1994±0.0204∗∗Res 300 μmol/L0.2185±0.0275∗∗0.1109±0.0186∗∗0.0388±0.0252∗∗
*P<0.05,**P<0.01vscontrol.
3 Res对ARPE-19细胞周期的影响
100、150和200 μmol/L 的Res处理ARPE-19细胞48 h后,S期细胞所占百分比分别为(38.80±0.28)%、(41.28±0.07)%和(49.57±0.41)%,与对照组[(11.33±0.03)% ]相比,比例增加,差异有统计学意义(P<0.05),见图3。实验结果显示Res对ARPE-19细胞的S期阻滞作用具有浓度-效应关系,随作用浓度的增加S期所占比例逐渐增加。
4 Res对ARPE-19细胞凋亡的影响
Res作用于ARPE-19细胞48 h后,Annexin V-FITC/PI双染流式细胞术检测结果显示,正常细胞在流式细胞分析图的B3区;早期凋亡在流式细胞分析图的B4区;晚期凋亡在流式细胞分析图的B2区;流式细胞分析图的B1区为机械损伤细胞。白藜芦醇对ARPE-19细胞没有毒性作用,方差分析显示,100、150 和200 μmol/L Res处理ARPE-19细胞48 h后的凋亡率与对照组没有差异(P>0.05),见图4。
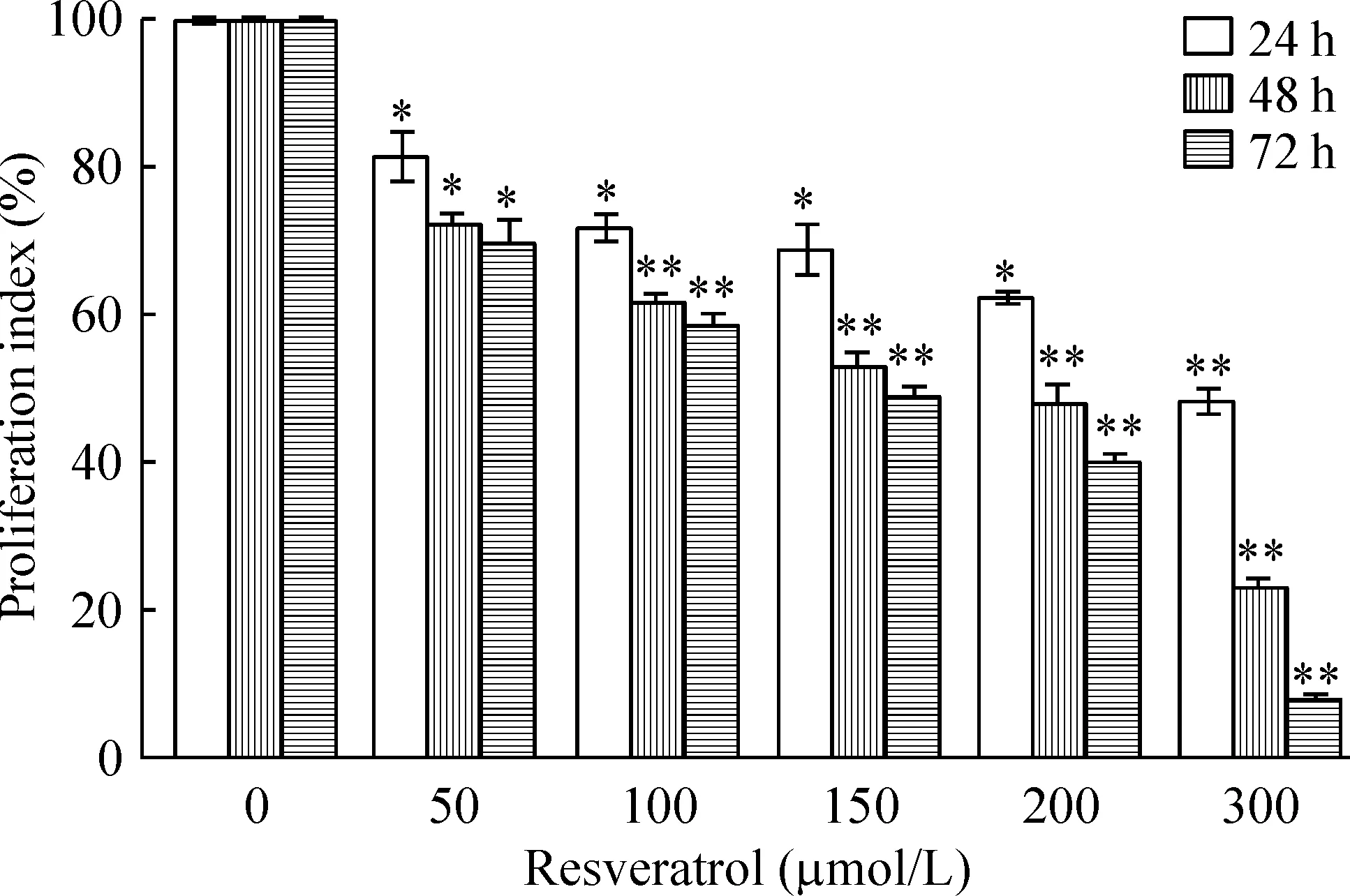
Figure 2. The inhibitory effects of resveratrol on ARPE-19 cell proliferation. Mean±SD. n=15. * P<0.05, ** P<0.01 vs 0 μmol/L.

Figure 3. The effects of resveratrol on the cell cycle of ARPE-19 cells. n=9.
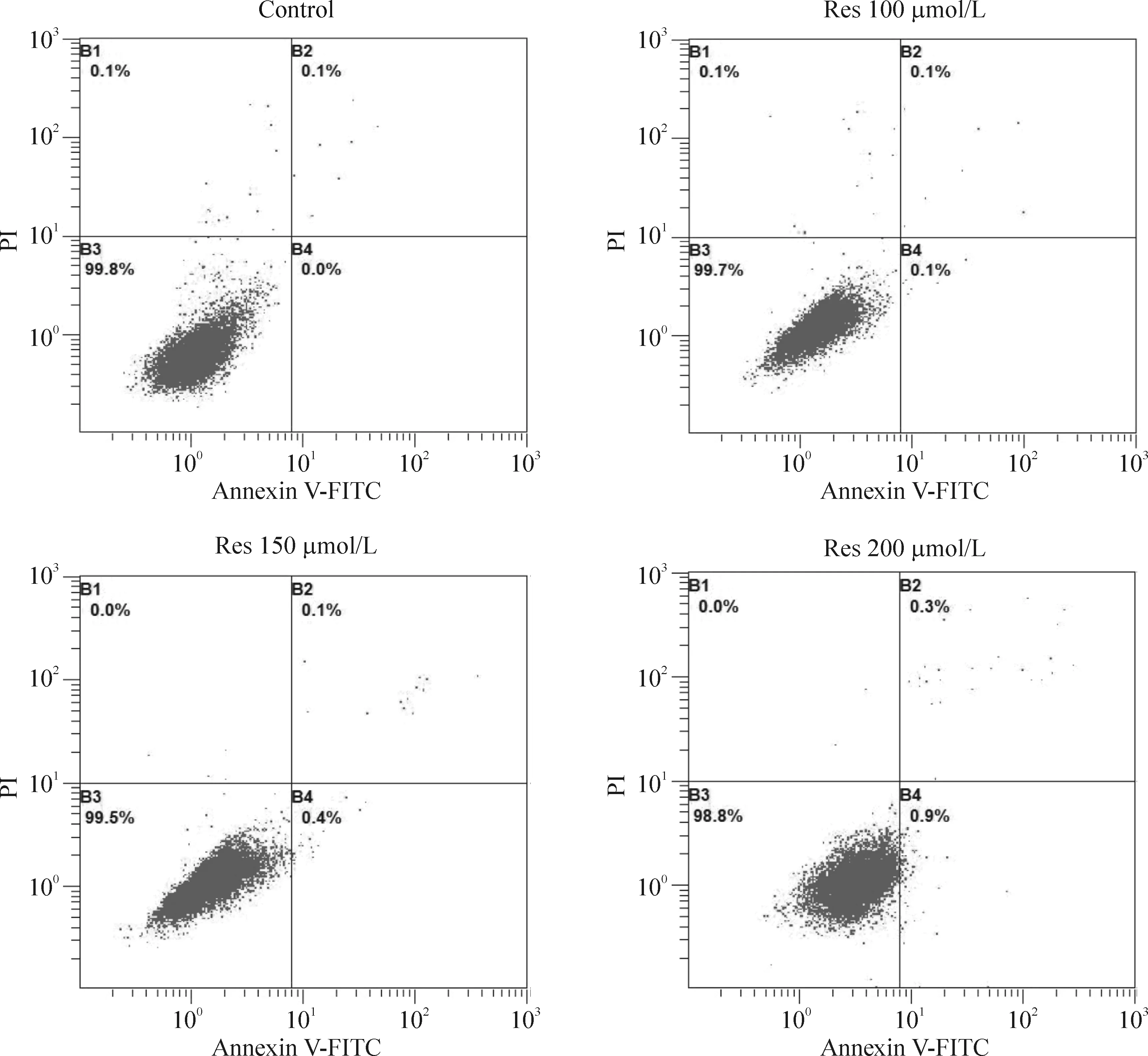
Figure 4. The effects of resveratrol on apoptosis of ARPE-19 cells.
5 免疫荧光检测Res对ARPE-19细胞表达PCNA蛋白的影响
共聚焦显微镜下观察,与对照组相比,150 μmol/L Res作用ARPE-19细胞48 h,胞核荧光明显减弱,PCNA蛋白表达减少,见图5。 这说明Res对ARPE-19细胞PCNA蛋白的表达有抑制作用。
6 Real-time PCR检测Res对ARPE-19细胞PCNA、P21和P27 mPNA表达的影响
与对照组相比,100、150和200 μmol/L Res组PCNA mRNA表达下调(P<0.01),且随作用浓度的增加PCNA mRNA的表达逐渐减少;而P21和P27的mRNA表达上调(P<0.05),且随作用浓度的增加,P21和P27 的mRNA表达逐渐增多,见图6。
讨 论
Res是一种生物性很强的天然多酚类物质,有抗氧化、抗肿瘤、抗血小板、抗增殖等多种生物学功能,可以预防和治疗动脉粥样硬化[9]、心脑血管疾病[10]、癌症[11]、年龄相关性黄斑病变[12]等多种疾病。关于Res抗增殖功能,抑制肿瘤细胞增殖及调节凋亡的作用和机制是目前研究的热点,而Res对视网膜色素上皮细胞增殖影响的报道却较少。
本实验用不同浓度Res作用ARPE-19细胞24、48和72 h后,通过CCK-8法检测发现Res对ARPE-19细胞的增殖有抑制作用,且在同一作用时间下,不同浓度的Res对ARPE-19细胞增殖的抑制程度不同,相同作用时间下,抑制率随药物浓度的增加而增强,呈现一定的浓度-效应依赖关系,同一浓度的Res对ARPE-19细胞增殖的抑制程度随作用时间延长而增强,呈现一定的时间-效应依赖关系,150 μmol/L Res作用48 h,能使ARPE-19细胞增殖抑制率达到50%以上,与其它研究结果相同[13]。细胞增殖是一个复杂的生物学过程,其依赖于细胞周期的正常运行且涉及多个基因正常的有序表达。本实验观察到Res作用48 h可诱导ARPE-19细胞周期阻滞在S期,且随药物浓度增加S期阻滞越明显。此外,Res各浓度组与对照组比较,对ARPE-19细胞凋亡率的影响无显著差异,表明Res对ARPE-19细胞没有毒性,与之前的报道一致[14]。
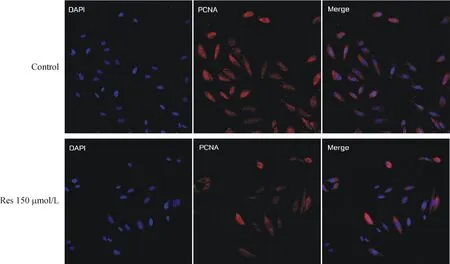
Figure 5. The effect of resveratrol on the protein expression of PCNA in ARPE-19 cells tested by immunofluorescence (×200).

Figure 6. The effects of resveratrol on the mRNA expression of PCNA, P21 and P27 tested by real-time PCR. Mean±SD. n=3. * P<0.05, ** P<0.01 vs 0 μmol/L.
PCNA是一种核内蛋白,是真核生物DNA聚合酶的亚单位、δ聚合酶的激活因子,通过与不同复制相关蛋白结合调节DNA复制过程,同时PCNA还通过不同调控方式与多种细胞因子作用,参与DNA损伤修复、细胞周期调控及凋亡等许多重要细胞事件,另外PCNA作为细胞增殖的指标,参与肿瘤等细胞增殖性疾病的发生发展[15-16]。通过免疫荧光方法对PCNA表达的检测,根据PCNA的表达强弱明确DNA复制多少来确定细胞增殖的促进或抑制。实验结果显示Res作用组ARPE-19细胞中PCNA表达下降,表明ARPE-19增殖减慢,生长受到抑制,与CCK-8、流式结果一致。
p21基因,全称p21WAF1/CIP1,定位于6号染色体p21.2,是p53基因的下游基因,其编码的蛋白P21是目前所知最广泛的细胞周期依赖性蛋白激酶抑制因子,可与周期蛋白(cyclin)、周期蛋白依赖性激酶(cyclin-dependent kinase,CDK)结合,使多数的cyclin-CDK复合物的激酶活性受到抑制,调控细胞周期使细胞停滞[17]。此外,P21WAF1/CIP1的羧基末端可与PCNA结合,阻止DNA全酶在DNA单链的滑动,从而抑制DNA的复制,调节细胞增殖,且P21蛋白可通过P53蛋白的调节间接参与DNA损伤修复过程[18]。p27基因定位于12号染色体,其编码的蛋白质P27类似于P21,是一种广谱CDK抑制剂,抑制cyclin D1-CDK4和cyclin E-CDK2复合物活性,对细胞周期及细胞增殖进行负性调节[19]。本实验结果显示,Res作用ARPE-19细胞48 h,ARPE-19细胞PCNA mRNA表达下降,而P21和P27 mRNA表达增加,呈一定的浓度依赖效应。
综上,本研究发现100~200 μmol/L的Res作用48 h可显著抑制ARPE-19细胞增殖,使细胞阻滞在S期,且没有细胞毒性,其作用机制可能与上调P21和P27表达及下调PCNA表达有关。因此,Res有望成为新型抗增殖药物,且其天然来源广泛。但是,Res对活体眼的药效和有无毒性反应仍需进一步实验验证,以期为临床治疗和预防增生性玻璃体视网膜疾病寻找新方法。
[参 考 文 献]
[1] Iandiev I, Bringmann A, Wiedemann P. Proliferative vitreoretinopathy pathogenesis and therapy[J]. Klin Monatsbl Augenheilkd, 2010, 227(3):168-174.
[2]PastorJC.Proliferativevitreoretinopathy:anoverview[J].SurvOphthalmol, 1998, 43(1):3-18.
[3] Rusnak S, Vrzalova J, Hecova L, et al. Defining the seriousness of proliferative vitreoretinopathy by aspiration of cytokines from the anterior chamber[J]. Biomark Med, 2013, 7(5):759-767.
[4]SadakaA,GiuliariGP.Proliferativevitreoretinopathy:currentandemergingtreatments[J].ClinOphthalmol, 2012, 6(8):1325-1333.
[5] 赵春晖,王毓琴,程凌云. 增生性玻璃体视网膜病变药物治疗新进展[J]. 中华眼科杂志, 2013, 49(8):752-757.
[6]ChenR,HollbornM,GroscheA,etal.Effectsofthevegetablepolyphenolsepigallocatechin-3-gallate,luteolin,apigenin,myricetin,quercetin,andcyanidininprimaryculturesofhumanretinalpigmentepithelialcells[J].MolVis, 2014, 20(3):242-258.
[7] Walle T. Bioavailability of resveratrol[J]. Ann N Y Acad Sci, 2011, 1215(1):9-15.
[8] 赵克森. 白藜芦醇的生物学特性和效应[J]. 中国病理生理杂志, 2012, 28(9):1709-1711.
[9] Tomayko EJ, Cachia AJ, Chung HR, et al. Resveratrol supplementation reduces aortic atherosclerosis and calcification and attenuates loss of aerobic capacity in a mouse model of uremia[J]. J Med Food, 2014, 17(2):278-283.
[10]OomenCA,FarkasE,RomanV,etal.Resveratrolpreservescerebrovasculardensityandcognitivefunctioninagingmice[J].FrontAgingNeurosci, 2009, 1(4):1-9.
[11] Hsieh T-C, Wong C, Bennett DJ, et al. Regulation of p53 and cell proliferation by resveratrol and its derivatives in breast cancer cells: an in silico and biochemical approach targeting integrin avβ3[J]. Int J Cancer, 2011, 129(11):2732-2743.
[12]SheuSJ,LiuNC,ChenJL.Resveratrolprotectshumanretinalpigmentepithelialcellsfromacrolein-induceddamage[J].JOculPharmacolTher, 2010, 26(3):231-236.
[13] Alex AF, Spitznas M, Tittel AP, et al. Inhibitory effect of epigallocatechin gallate (EGCG), resveratrol, and curcumin on proliferation of human retinal pigment epithelial cellsinvitro[J]. Curr Eye Res, 2010, 35(11):1021-1033.
[14]KingRE,KentKD,BomserJA.Resveratrolreducesoxidationandproliferationofhumanretinalpigmentepithelialcellsviaextracellularsignal-regulatedkinaseinhibition[J].ChemBiolInteract, 2005, 151(2):143-149.
[15] Strzalka W, Ziemienowicz A. Proliferating cell nuclear antigen (PCNA): a key factor in DNA replication and cell cycle regulation[J]. Ann Bot, 2011, 107(7):1127-1140.
[16]WangSC.PCNA:asilenthousekeeperorapotentialtherapeutictarget?[J].TrendsPharmacolSci, 2014, 35(4):178-186.
[17] Gartel AL, Serfas MS, Tyner AL. p21: negative regulator of the cell cycle[J]. Proc Soc Exp Biol Med, 1996, 213(2):138-149.
[18] 廖小方, 赵中洪, 郭绍文,等. 甘氨双唑钠协同顺铂有效促进肺腺癌细胞中p21表达[J]. 中国病理生理杂志, 2014, 30(2):256-261.
[19] Moller MB. p27 in cell cycle control and cancer[J]. Leuk Lymphoma, 2000, 39(1-2):19-27.
猜你喜欢
现代仪器与医疗(2022年2期)2022-08-11
中风与神经疾病杂志(2021年9期)2021-11-08
中医眼耳鼻喉杂志(2021年1期)2021-07-22
中医眼耳鼻喉杂志(2021年2期)2021-07-21
世界最新医学信息文摘(2021年12期)2021-06-09
中成药(2017年9期)2017-12-19
中成药(2017年9期)2017-12-19
中成药(2017年7期)2017-11-22
中成药(2017年5期)2017-06-13
标记免疫分析与临床(2016年9期)2016-11-21
