
黑河下游绿洲胡杨生长状况与叶生态特征
2014-08-08 02:15赵传燕彭守璋李文娟
生态学报 2014年16期
赵传燕,赵 阳,彭守璋,王 瑶,李文娟
(兰州大学,草地农业生态系统国家重点实验室, 兰州 730000)
黑河下游绿洲胡杨生长状况与叶生态特征
赵传燕*,赵 阳,彭守璋,王 瑶,李文娟
(兰州大学,草地农业生态系统国家重点实验室, 兰州 730000)
选择黑河下游额济纳绿洲为研究区,以优势物种胡杨(PopuluseuphraticaOliv.)为研究对象。在2009年和2010年对研究区胡杨的生长状况、叶生态特征和生境进行了调查。调查结果表明:胡杨受水分胁迫的程度增强,生长态势变差,其枯枝比由2.45%增加到81.00%,其比叶面积由11.84 m2/kg减少到5.35 m2/kg。叶气孔密度变化很大,最小值为105(个/mm2),最大值为218(个/mm2),平均值为158.4(个/mm2),随着地下水埋深的增加,叶气孔密度先减少后增加之后再显著减少(P<0.05),呈三次函数关系。研究得出枯枝比能够反映胡杨分布区地下水的变化状况,是林相描述比较重要的指标,比叶面积和叶气孔密度能够指示胡杨种群环境变化。研究结果可为荒漠河岸林恢复和生态输水效应评价提供科学依据。
黑河下游;胡杨;生长态势;枯枝比;比叶面积; 气孔密度
胡杨(Populuseuphratica)是第三纪上新世遗留下的杨柳科杨属中最古老、最原始的孑遗树种[1],是维持荒漠河岸林生态平衡的关键物种和被联合国粮农组织(Food and Agriculture Organization of the United Nations, FAO)确定为最急需优先保护的林木基因资源。它主要生长在中纬度的干旱荒漠地区,其生境非常严酷,冬季酷寒,夏季炎热,干燥多风,降水稀少,风蚀、风积强烈,土壤盐渍化程度高。在极端环境下,胡杨生存的最关键的限制因子是浅层地下水[2- 3]。由于农业的发展, 上中游来水量急剧减少,如黑河下游来水量由20世纪60年代以前的10亿m3减少到现在2—4亿m3[4],从而大大降低了胡杨分布区的地下水位。胡杨失去水源,长势明显衰退,表现在冠幅变小,枯稍、枯枝明显增加[5]。调查表明:胡杨在地下水埋深小于4 m 时,生长正常;当地下水埋深在4—6 m 时,胡杨生长不良, 开始出现秃顶、叶枯现象, 少数死亡;当地下水埋深达6—10 m 时, 大部分胡杨枯死[2]。枯死枝条占所有枝条的比例(枯枝比)能够反映胡杨在不同的生境条件下的生长状态[6],但是目前对于胡杨枯枝比的定量研究鲜见。
叶是植物与环境接触面积最大的器官,由此成为植物对逆境胁迫最敏感的部位,在长期自然选择的过程中常常形成与逆境相适应的特殊结构和功能[7]。例如,在干旱区,叶面积变小或退化成刺、叶拥有厚的角质层并被蜡质层、叶细胞浓度大等,这些特殊结构和功能用以减少水分散失或增加水分吸收来适应干旱或者水分短缺[8]。反映叶片生态特征的主要参数有:气孔密度、叶变形系数、比叶面积等[9- 11]。在环境胁迫下,它们依据胁迫的程度有不同的响应。结合胡杨生长状况综合利用叶片生态参数反映水分胁迫的研究报道较少。因此,本研究的目的是:(1)利用胡杨枯枝比定量描述胡杨的生境与生长状态的关系;(2)在不同的生长状态下叶的形态特征。该研究不仅有助于了解胡杨适应干旱的内部调节机制,而且也有利于制订胡杨的保护措施和恢复对策。
1 研究区域与数据收集
1.1 研究区概况
黑河下游位于内蒙古自治区最西端的额济纳旗境内,地理位置介于东经100°10′—101°30′、北纬41°48′—42°42′之间。本区为极端干旱的温带荒漠。年平均气温8.3 ℃,7月平均气温26.3 ℃,1月平均气温-12 ℃, 极端最高气温43.1 ℃, 极端最低气温-37.6 ℃。全年日照时数3171.2 h,年平均降水41.3 mm,年平均蒸发量3706 mm,年平均8 级以上大风日数52 d[12]。黑河流域下游是个只有侧向流入而无侧向流出的半封闭盆地,黑河干流的河水是地下水的主要补给源。在干旱的大陆性气候、自身盆地地形条件以及黑河河水浸渍作用的影响下,土壤类型主要为盐化草甸土、潮土(灰色草甸土)、盐土和风沙土。从绿洲内部沿河地区到外围戈壁、低山丘陵区,地下水埋深逐渐加深[3],受地下水埋深的控制,植被的演变趋势是: 芦苇(Phragmitesaustralis)、芨芨草(Achnatherumsplendens)→胡杨(Populuseuphratica)、沙枣(Elaeagnusangustifolia)乔木林→柽柳(Tamarixramosissima)灌木林→苏枸杞(Lyciumruthenicum)、盐爪爪(Kalidiumfoliatum)耐旱盐草→梭梭(Haloxylonammodendron)、红沙(Reaumuriasoongorica)超旱生灌木。在研究区内胡杨多为成熟林,生长状况不良,平均密度为163.4株/ hm2,平均树高11.1 m,平均胸径59.26 cm,地下水埋深在2.00—8.46 m。胡杨叶形多样,根据树龄和分布空间不同可出现披针形叶、卵圆形叶和锯齿卵圆形叶。另外, 胡杨叶片厚,角质层厚,表面被蜡质层,对大气和土壤干旱能力适应性强。
1.2 数据收集
1.2.1 样地的选择与采样
野外观测于2009年8月和2010年8月,研究区选择在黑河下游东河中下部区域,位于101°1′13.94″—101°8′12.63″E,42°9′46.36″—42°4′28.36″N,面积约100 km2。在研究区布设42个胡杨样圆,分布在距河道不同的距离内,样圆半径分布在10—50 m之间,其大小依据采样点植株的密度而定。用手持式GPS (Garmin Map60csx,美国)在样圆中心进行定位,利用罗盘和激光测高仪测定每株树木在样圆分布的相对位置(图1),同时记录每木树高和胸径。
在样圆内选择平均木1—4株,利用日本Nikon公司生产的数码照相机(D80)对每株按东、南、西、北四个方向进行水平方向拍照,获取冠层图像,利用专业的面向对象分类软件(eCognition)对图像进行分类,将分类图像导入ArcGIS软件中计算枯枝占所有枝条的比例,每株的枯枝比为四个方向的平均。

图1 样圆中树木的分布及相对位置Fig.1 Distribution and relative position of trees in a plot
选择胡杨3—5株,挂牌标记,对每株树木上部按东、南、西、北四个方位选取完全成熟、健康叶30—40片,取出10片左右投入FAA (Formalin-Acetic acid-Alcohol)固定液中,防止由于叶片失水而导致的气孔变化。其余用KP- 90N(日本造)量积仪测量叶面积,叶片面积测量完毕,装入牛皮信封,放入75 ℃烘箱中烘至恒量,这些数据用于计算单位重量叶面积(即比叶面积)。
1.2.2 气孔密度的估算
将FAA固定液中的叶片捞出,洗净。在每个叶片主叶脉一侧,选上中下3个部分均匀涂抹无色透明的指甲油,待其晾干后,用透明胶带将其粘取下来置于载玻片上,将载玻片于40倍Leica DM6000显微镜(德国造)下拍照。每一载玻片上随机选5个视野,即每一叶片共拍15幅图片。利用图像处理技术提取气孔数量的方法见参考文献[13]。
2 结果分析
2.1 胡杨种群的基本特征
42个样圆观测得到的胡杨种群特征统计值见表1。从表1可以看出:胡杨树高度分布在6.3—20.1 m之间,平均值为11 m,胸径分布在13.53—135.92 cm之间,平均值为59.26 cm,枯枝比(枯枝占所有枝条的比例)差异较大,分布在2.45%—81.00%之间,各样圆树木密度差异显著(P<0.001),分布在8.0—828.0株/hm2之间,平均值为163.4株/hm2,地下水位在198—846 cm之间,平均地下水位为435.6 cm。

表1 42个样圆调查统计
2.2 枯枝比的变化
枯枝比(Ratio of died branches to full branches, RBF)是枯死枝条占所有枝条的比例。由图2可以看出,在42个样圆中胡杨的枯枝比差异较大,分布在2.45%—81%,最大值是最小值的40倍。以1 m为间隔,对样圆所在地的地下水进行归类,不同类别的枯枝比见图3。总之,随着地下水位的增加,枯枝比明显增加。当地下水位< 4 m时,平均枯枝比为13.84%,变化范围在2.45%—21.84%之间,林木生长较好,林相整齐,有较多的更新幼苗,平均树木年龄较小;当地下水位4—6 m之间,平均枯枝比为32.68%,变化范围在21.84%—46.27%之间,林木生长不良,林相不整齐,更新幼苗不多,平均树木年龄较大;当地下水位6—10 m之间,平均枯枝比为57.55%,变化范围在31%—81.04%之间,林木生长较差,林相残败,立木多枯顶,树干中空,林木濒临死亡。
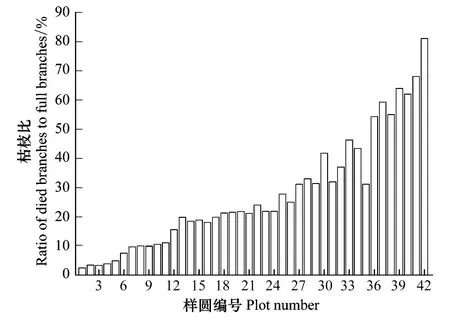
图2 不同样圆中枯枝比的变化Fig.2 Variation of ratio of died branches to full branches in 42 plots
2.3 比叶面积的变化
比叶面积(SLA)是叶单位重量的叶面积,由图4可以看出,胡杨的比叶面积分布在5.35—11.84 m2/kg,最大值是最小值的近2倍,随着水分胁迫程度的增加,比叶面积逐渐减小。比叶面积大于10 m2/kg
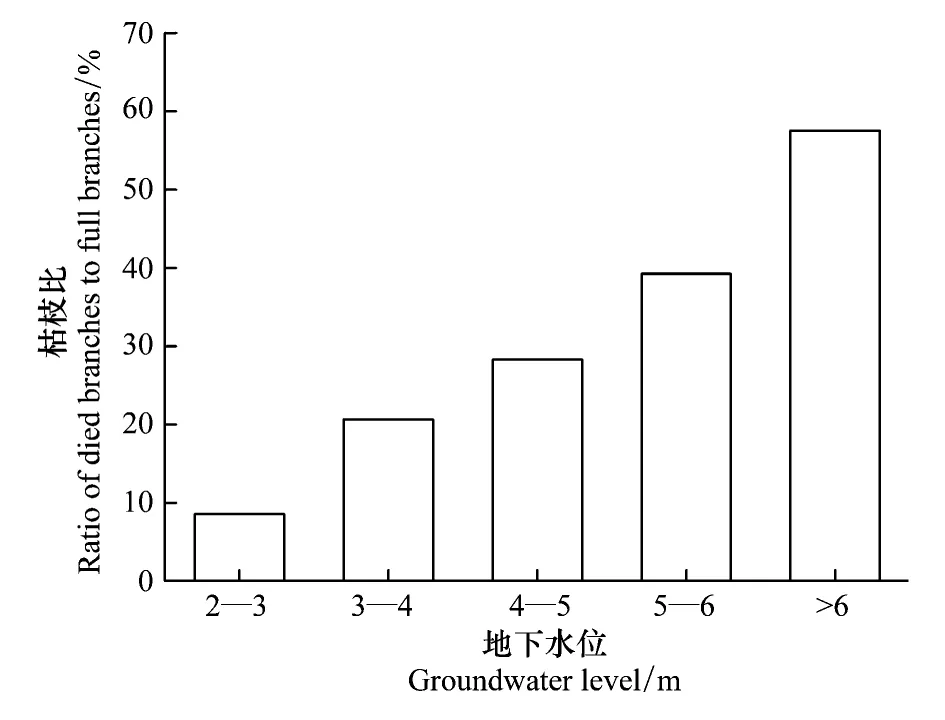
图3 不同地下水位条件下枯枝比的变化Fig.3 Variation of ratio of died branches to full branches with five classifications of groundwater level
的样圆多分布在最适宜的生境中,枯枝比一般小于10%。比叶面积在8—10 m2/kg之间,树木生长在较好的生境中,枯枝比一般小于20%。比叶面积在6.5—8 m2/kg,树木长势较差,枯枝比较大,多处于20%—50%之间,比叶面积小于6.5 m2/kg的样圆中林木生长差、林相残败、立木多枯顶,枯枝比一般大于50%。由此可以看出,比叶面积与枯枝比呈反比例关系(图4)。

图4 不同样圆中比叶面积与枯枝比的变化比较Fig.4 Comparison of variation between special leaf area and ratio of died branches to full branches in 42 plots
2.4 气孔密度的变化
气孔密度(SD)是单位叶面积上气孔的数目。在42个不同生境的样圆中,胡杨叶气孔密度变化很大,胡杨叶气孔密度的最小值为105(个/mm2)出现在34号样圆中,气孔密度的最大值为218(个/mm2),出现在22号样圆中,平均的气孔密度为158.4(个/mm2)。它与胡杨生长状态的关系比较复杂,较大值出现在枯枝比<4%和20%—33%的范围内。即随着水分胁迫的增加,气孔密度逐渐减小,在枯枝比达到10%左右达到低谷,随后又逐渐增加,在枯枝比30%左右达到峰值,之后又随着枯枝比的增加而下降(图5)。
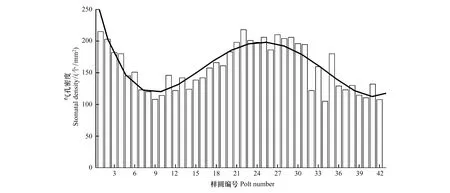
图5 气孔密度在不同的样圆中的变化Fig.5 Variation of stomatal density in 42 plots
3 讨论
3.1 胡杨生长态势与生境的关系
胡杨生长态势与生境的关系从林相上可以得到明显的反应。如果地下水位适宜,林木生长较好,林相整齐,树冠饱满;如果地下水位不适宜,林木生长较差,林相残败,立木多枯顶,树干中空,林木濒临死亡。地下水位介于适宜和不适宜之间,林木生长不良,林相不整齐,有枯梢和枯枝现象。前人对胡杨地下水位的适宜性进行了大量的研究,樊自立等[14]根据潜水蒸发与土壤盐渍化与荒漠化的关系,提出适宜生态水位(2—4 m)、生态胁迫水位(4—6 m)和荒漠化水位(>6 m)。根据地下水位与胡杨频度的关系模型确定的最佳地下水位为2.51 m,平均地下水位为4.52 m[2],利用地下水位与胡杨盖度的关系模型确定的胡杨生长最佳地下水位为2.6 m[15]。但是反映林相的关键参数-枯枝比定量化研究不多。枯枝是胡杨林相最直观的表征,能够反映胡杨生长态势并能够迅速指示胡杨分布区地下水位的变动,因此枯枝比的量化研究值得重视。
3.2 比叶面积变化与其环境解释
比叶面积(SLA)是植物叶片的重要性状之一,其大小主要取决于叶片组织密度和叶片厚度。已有研究表明,SLA可以反映植物获取资源的能力,低SLA的植物能更好的适应资源贫瘠和干旱的环境,高SLA的植物保持体内营养的能力较强[16- 17]。Ellsworth和Reich[18]研究得出,对于同一树种,植株冠层上部的SLA通常低于下部,具体表现在植物叶片厚度的增加和叶肉细胞密度的增大,在一定程度上反映了叶片截获光的能力和在强光下的自我保护能力[19]。因而SLA成为植物比较生态学研究中的首选指标[20]。本研究结果表明,随着胡杨生长态势的差别,SLA有明显的反映,比叶面积处于5.35—11.84 m2/kg之间。高值出现在胡杨生长状况良好样圆内,说明环境提供给胡杨生长的资源充足;低值出现在胡杨生长状况较差的样圆内,说明供植物利用的资源相对较少,是胡杨适应贫瘠环境的结果。吴晓成等研究了额尔齐斯河天然杨树林的SLA,得出不同径级不同物种SLA范围在10.41—79.49 m2/kg[16],但是胡杨SLA的研究没有报道。随着图像处理与扫描技术的发展,室外精确获取大量SLA数据已不再是难事,再者,胡杨SLA对环境变化有明显的指征,能够明显体现水分胁迫程度,是胡杨叶生态研究的重要指标。
3.3 气孔密度的变化与其环境解释
气孔是植物叶片与外界进行气体交换的通道,叶片内气孔数目的多少直接影响着蒸腾和光合作用的强弱。气孔密度大小受水分、温度、地形条件等影响[21]水分条件与叶片气孔密度的关系研究较多[22- 25],研究结果显示随着水分胁迫程度的加剧,气孔密度呈现上升趋势,主要的原因是干旱胁迫,叶面积减小造成气孔密度增加。但是本研究的结果与该规律并不吻合,胡杨随着干旱胁迫加强,气孔密度表现下降上升再下降的趋势。即在轻度胁迫条件下,气孔密度减少,在中度胁迫条件下气孔密度增加,在重度胁迫条件下,气孔密度显著减小。目前对胡杨气孔密度的研究主要限于对不同叶形气孔密度的研究[26],但对于成年胡杨在不同生境条件下气孔密度的变化没有报道。是否随着干旱胁迫程度的加强,气孔密度都呈现增加趋势?刘世鹏等对5个枣树品种叶片气孔密度进行研究,得出随水分胁迫程度的增加,气孔密度表现为先升后降的趋势[27],在重度水分胁迫下,气孔密度呈现下降趋势,与本研究的结果一致。王学臣等[28]认为重度水分胁迫时,光合作用受到严重影响,减少了植株的能量供应,抑制了气孔细胞的生长发育,细胞分裂、伸长和分化受到影响,气孔数目显著减少,表现为气孔密度下降。在中度水分胁迫下,光合作用的降低又会影响细胞的伸长,造成叶面积减小,所以气孔密度上升[29]。在轻度水分胁迫下,会导致气孔关闭,从而降低光合速率,保持水分,而气孔数目并无明显变化,但是本研究中气孔密度在轻度胁迫下有明显下降趋势,其机理过程有待研究。
4 结论
从以上研究和分析中可得到如下结论:
(1) 胡杨受水分胁迫的程度不同,其生长态势有较大的差异,枯枝比是生长状况的重要指标,是生长状况与生境的直观表达参量。
(2) 随着生长状况的不同,胡杨叶在形态和生理上都有相应的反应。尤其是气孔密度和比叶面积响应明显。比叶面积随着胡杨生长状况的变差而减小,气孔密度的变化与水分胁迫程度关系比较复杂。
(3) 本次研究中,利用图像处理技术获得了枯枝比,使林相描述由定性到定量,是遥感技术在生态学中应用的又一体现。
[1] Wang S J, Chen B H, Li H Q.EuphratesPoplar. Beijing: Chinese Environmental Science Press, 1995: 3- 18.
[2] Zhang L, Dong Z C, Huang X L. Modeling on relation between major plants growth and groundwater depth in arid area. Journal of Desert Research, 2004, 24(1): 110- 113.
[3] Zhao C Y, Li S B, Feng Z D, Shen W H. Dynamics of groundwater level in the water table fluctuant belt at the lower reaches of Heihe River. Journal of Desert Research, 2009, 29(2): 365- 369.
[4] Wang L M, Zhang Q L, Yin J Y. Study on the growth pattern and bio-productivity of thePopuluseuphraticaforest stand in Ejina. Journal of Arid Land Resources and Environments, 2003, 17(2): 94- 99.
[5] Xu H L, Chen Y N, Ya G. Effect of translating water on vegetation at the lower reaches of Tarim River. Environmental Science, 2003, 24(4): 18- 22.
[6] Niu T, Li X, Zhang H F, Wang J G, Lin H J. Ecological characterization ofPopuluseuphratica′s response to non-deterministic water delivery. Resources Science, 2012, 34(5): 819- 826.
[7] Li W H, Zhuang L, Gong W C, Zhao W Q, Tian Z P. Ultramicroscopic structure ofPopuluseuphraticaleaves related to environmental heterogeneity in the lower reaches of Tarim river. Journal of Desert Research, 2009, 29(4): 680- 687.
[8] Yan C R, Han X G, Chen L Z. The relationship between the ecophysiological feature and leaf characteristics of some woody plants in Beijing mountain zone. Acta Ecologica Sinica, 2000, 20(1): 53- 60.
[9] He S M, Wang J H, Qin J S. Comparison of tissue structure characteristics and drought-resistant properties in some hippophae species leaves. China Forestry Science and Technology, 2009, 23(1): 16- 19.
[10] Tian Q, Cao Z Z, Wang G H. Response of leaf traits of 14 plant species in typical steppe to water gradient in Inner Mongolia. Grassland and Lawn, 2008, (5): 23- 26.
[11] Sun C X, Shen X Y. Advances in studies on identification indexes and methods of quantitative analyses for crop drought resistance. Chinese Agricultural Science Bulletin, 2002, 18(1): 49- 51.
[12] Wuri G F, Zhang S H, Cheng J Q, Zhang X L, Ban Z G. On drought-resistant and propagation mechanism of natural forest ofPopuluseuphraticaoliver in Ejina Banner. Inner Mongolia Forestry Investigation and Design, 2003, 26(4): 1- 5.
[13] Jian S Q, Zhao C Y, Zhao Y, Peng S Z, Peng H H. Based on image processing technology estimatingleaves stomatal density ofPopuluseuphraticaand analysis of its ecological significance. Acta Ecologica Sinica, 2011, 31(17): 4818- 4825.
[14] Fan Z L, Ma Y J, Zhang H, Wang R H, Zhao Y J, Zhou H F. Research of eco-water table and rational depth of groundwater of Tarim rriver drainage basin. Arid Land Geography, 2004, 27(1): 8- 13.
[15] Zhao C Y, Li S B, Jia Y H, Jiang Y C. Dynamic changes of groundwater level and vegetation in water table fluctuant belt in lower reaches of Heihe river: Coupling simulation. Chinese Journal of Applied Ecology, 2008, 19(12): 2687- 2692.
[16] Wu X C, Zhang Q L, Zang R G, Lei Q Z. Leaf area index and specific leaf area of natural poplars in Ergis Basin. Journal of Northwest Forestry University, 2009, 24(4): l0- 15.
[17] Wu D D, Zhou Y B, Yu D P, Dai G H. Physiological response ofBetulaermaniiat different altitudes in Changbai Mountain. Acta Ecologica Sinica, 2009, 29(5): 2279- 2285.
[18] Ellsworth D S, Reich P B. Canopy structure and vertical patterns of photosynthesis and related leaf traits in a deciduous forest. Oecologia, 1993, 96(2): 169- 178.
[19] Zhang L, Luo T X. Advances in ecological studies on leaf lifespan and associated leaf traits. Journal of Plant Ecology, 2004, 28(6): 844- 852.
[20] Garnier E, Shipley B, Roumet C, Laurent G. Standardized protocol for the determination of specific leaf area and leaf dry matter content. Functional Ecology, 2001, 15(5): 688- 695.
[21] Yang L M, Han M, Zhou G S, Li J D. The changes of water-use efficiency and stoma density of Leymus chinensis along Northeast China Transect. Acta Ecologica Sinica, 2007, 27(1): 16- 24.
[22] Wang G F, Li L G, Li X Y, Du S Y, Zhang L Y, Xie L F. Relationship between the morphological charateristics of leaf surface and drought resistance of sea buckthorn. Acta Horticulturae Sinica, 2006, 33(6): 1310- 1312.
[23] Wen G S, Zhang M R, Zhang G S, Wang L H. Ecophysiological strategy of Sabina vulgaris under drought stress. Acta Ecologica Sinica, 2006, 26(12): 4059- 4065.
[24] Yang H M, Wang G X. Leaf stomatal densities and distribution inTriticumaestivumunder drought and CO2enrichment. Acta Phytoecologica Sinica, 2001, 25(3): 312- 316.
[25] Yu H Q, Wu Z H, Shen X Y, Xu K Z. Changes of stomatal density, length, width and microstructure of maize leaves under water stress. Journal of Jilin Agricultural University, 2003, 25(3): 239- 242.
[26] Zheng C X, Qiu J, Jiang C N, Gao R F, Wang W F. Comparison of characteristics of stomas and photosynthesis ofPopuluseuphraticapolymorphic leaves. Scientia Silvae Sinicae, 2006, 42(8): 19- 24.
[27] Liu S P, Liu J M, Cao J Y, Bai C Y, Shi R. Stomatal distribution and character analysis of leaf epidermis ofJujubeunder drought stress. Journal of Anhui Agricultural Sciences, 2006, 34(7): 1315- 1318.
[28] Wang X C, Ren H Y, Lou C H. Root to shoot communication in the responses to drought. Plant Physiology Communications, 1992, 28(6): 397- 402.
[29] Li H B. Effects of NaCl stress and water stress on stomatal characters and other physiological characters of rice leaf. Shenyang: Shenyang Agricultural University, 2004.
参考文献:
[1] 王世绩, 陈炳浩, 李护群. 胡杨林. 北京: 中国环境科学出版社, 1995: 3- 18.
[2] 张丽, 董增川, 黄晓玲. 干旱区典型植物生长与地下水位关系的模型研究. 中国沙漠, 2004, 24(1): 110- 113.
[3] 赵传燕, 李守波, 冯兆东, 沈卫华. 黑河下游地下水波动带地下水位动态变化研究. 中国沙漠, 2009, 29(2): 365- 369.
[4] 王立明, 张秋良, 殷继艳. 额济纳胡杨林生长规律及生物生产力的研究. 干旱区资源与环境, 2003, 17(2): 94- 99.
[5] 徐海量, 陈亚宁, 杨戈. 塔里木河下游生态输水对植被和地下水位的影响. 环境科学, 2003, 24(4): 18- 22.
[6] 牛婷, 李霞, 张绘芳, 王建刚, 林海军. 胡杨对非确定性输水响应的生态表征. 资源科学, 2012, 34(5): 819- 826.
[7] 李卫红, 庄丽, 公维昌, 赵文勤, 田中平. 塔里木河下游胡杨叶片变化与环境异质性. 中国沙漠, 2009, 29(4): 680- 687.
[8] 严昌荣, 韩兴国, 陈灵芝. 北京山区落叶阔叶林优势种叶片特点及其生理生态特性. 生态学报, 2000, 20(1): 53- 60.
[9] 何士敏, 汪建华, 秦家顺. 几种沙棘叶片组织结构特点和抗旱性比较. 林业科技开发, 2009, 23(1): 16- 19.
[10] 田青, 曹致中, 王国宏. 内蒙古多伦典型草原14种植物比叶面积对水分梯度变化的响应. 草原与草坪, 2008, (5): 23- 26.
[11] 孙彩霞, 沈秀瑛. 作物抗旱性鉴定指标及数量分析方法的研究进展. 中国农学通报, 2002, 18(1): 49- 51.
[12] 乌日根夫, 战士宏, 程继全, 张晓丽, 班振国. 额济纳旗天然胡杨林生物学、生态学抗旱机理与繁殖机理研究. 内蒙古林业调查设计, 2003, 26(4): 1- 5.
[13] 荐圣淇, 赵传燕, 赵阳, 彭守璋, 彭焕华. 基于遥感图像处理技术胡杨叶气孔密度的估算及其生态意义. 生态学报, 2011, 31(17): 4818- 4825.
[14] 樊自立, 马英杰, 张宏, 王让会, 赵元杰, 周宏飞. 塔里木河流域生态地下水位及其合理深度确定. 干旱区地理, 2004, 27(1): 8- 13.
[15] 赵传燕, 李守波, 贾艳红, 姜云超. 黑河下游地下水波动带地下水与植被动态耦合模拟. 应用生态学报, 2008, 19(12): 2687- 2692.
[16] 吴晓成, 张秋良, 臧润国, 雷庆哲. 额尔齐斯河天然杨树林叶面积指数及比叶面积的研究. 西北林学院学报, 2009, 24(4): 10- 15.
[17] 吴栋栋, 周永斌, 于大炮, 戴冠华. 不同海拔长白山岳桦的生理变化. 生态学报, 2009, 29(5): 2279- 2285.
[19] 张林, 罗天祥. 植物叶寿命及其相关叶性状的生态学研究进展. 植物生态学报, 2004, 28(6): 844- 852.
[21] 杨利民, 韩梅, 周广胜, 李建东. 中国东北样带关键种羊草水分利用效率与气孔密度. 生态学报, 2007, 27(1): 16- 24.
[22] 王国富, 李连国, 李晓燕, 杜世勇, 张丽盈, 谢丽芬. 沙棘叶片表面形态特征与抗旱性的关系. 园艺学报, 2006, 33(6): 1310- 1312.
[23] 温国胜, 张明如, 张国盛, 王林和. 干旱条件下臭柏的生理生态对策. 生态学报, 2006, 26(12): 4059- 4065.
[24] 杨惠敏, 王根轩. 干旱和CO2浓度升高对干旱区春小麦气孔密度及分布的影响. 植物生态学报, 2001, 25(3): 312- 316.
[25] 于海秋, 武志海, 沈秀瑛, 徐克章. 水分胁迫下玉米叶片气孔密度、大小及显微结构的变化. 吉林农业大学学报, 2003, 25(3): 239- 242.
[26] 郑彩霞, 邱箭, 姜春宁, 高荣孚, 汪万福. 胡杨多形叶气孔特征及光合特性的比较. 林业科学, 2006, 42(8): 19- 24.
[27] 刘世鹏, 刘济明, 曹娟云, 白重炎, 师荣. 干旱胁迫下枣树叶片表皮气孔分布及特征分析. 安徽农业科学, 2006, 34(7): 1315- 1318.
[28] 王学臣, 任海云, 娄程后. 干旱胁迫下植物根与地上部间的信息传递. 植物生理学通讯, 1992, 28(6): 397- 402.
[29] 李海波. 水分亏缺和盐胁迫对水稻叶片气孔及其他生理性状的影响. 沈阳: 沈阳农业大学, 2004.
The growth state ofPopuluseuphraticaOliv. and its leaf ecological characteristics in the lower reaches of Heihe River
ZHAO Chuanyan*, ZHAO Yang, PENG Shouzhang, WANG Yao, LI Wenjuan
StateKeyLaboratoryofGrasslandAgro-ecosystems,LanzhouUniversity,Lanzhou730000,China
The study was conducted in the lower reach of Heihe River, which is located in an extremely arid desert in northwest China. We pay a particular attention onPopuluseuphraticaOliv that adapts to continental arid climate and is a dominant component of the riparian ecosystem. Groundwater levels in the riparian ecosystem may have influences on states of plant growth. In turn, plant leaves may reflect different growth states. To reveal the growth state and leaf-related physiological responses ofPopuluseuphraticaOliv forest to groundwater table variations, 42 sampling plots ofPopuluseuphraticaOliv under different DG were selected to investigate the relationship betweenPopuluseuphraticaOliv and its inhabit. We investigated the ratio of died branches to total branches, the specific leaf area (SLA), and the stomatal density (SD) ofPopuluseuphraticaOliv, and DG in 2009 and 2010. The result showed that the ratio of died branches to total branches increased continually with the increasing DG, ranging from 2.45% to 81.00%, whereas the SLA decreased with the increasing DG from 11.84 m2/kg to 5.35 m2/kg. In contrast, the SLA decreased with the increasing DG. Additionally, the SD has a complex change trend with the increasing DG. The SD first decreased, then increased, and at last decreased again with the increasing DG. The minimum of SD is 105 (pore/mm2), the maximum is 218 (pore/mm2),and the mean value is 158.40 (pore/mm2). We can draw conclusions thatPopuluseuphraticaOliv suffers from various degrees of water stress due to changes in the DG. The ratio of died branches to total branches can obviously be used to indicate the variation of groundwater level.PopuluseuphraticaOliv has low productivity under severe water stress due to SLA and SD decline. Therefore it is necessary to balance water requirement between humans and nature and protectPopuluseuphraticaOliv from extinction in this area. SD and SLA are important parameters to measure the response of the growth state ofPopuluseuphraticaOliv. These findings should be helpful in monitoring the growth and development ofPopuluseuphraticaOliv forests as well as in assessing the efficiency of ecological water delivery in the study areas.
the lower reaches of Heihe River;PopuluseuphraticaOliv.; the growth state; ratio of died branches to total branches; specific leaf area; stomatal density
国家自然科学基金项目(91025015)
2012- 12- 25; 网络出版日期:2014- 03- 04*通讯作者Corresponding author.E-mail: nanzhr@lzb.ac.cn
10.5846/stxb201212251867
赵传燕,赵阳,彭守璋,王瑶,李文娟.黑河下游绿洲胡杨生长状况与叶生态特征.生态学报,2014,34(16):4518- 4525.
Zhao C Y, Zhao Y, Peng S Z, Wang Y, Li W J.The growth state ofPopuluseuphraticaOliv. and its leaf ecological characteristics in the lower reaches of Heihe River.Acta Ecologica Sinica,2014,34(16):4518- 4525.
猜你喜欢
思维与智慧·上半月(2022年4期)2022-04-08
金桥(2021年11期)2021-11-20
湖南农业大学学报(自然科学版)(2021年3期)2021-07-02
北广人物(2020年47期)2020-12-09
青年文学家(2020年13期)2020-05-26
新疆农业科学(2020年1期)2020-02-14
创新作文(1-2年级)(2018年6期)2018-01-22
甘肃林业(2016年4期)2016-11-07
中学生(2016年25期)2016-03-01
中国麻业科学(2015年5期)2015-12-28
