
早期游泳运动对过度喂养肥胖大鼠内脏脂肪积累和食欲肽表达的影响
2014-08-08 01:01何玉秀牛凌坤
中国体育科技 2014年2期
高 峰,何玉秀,郝 强,牛凌坤
早期游泳运动对过度喂养肥胖大鼠内脏脂肪积累和食欲肽表达的影响
高 峰1,何玉秀1,郝 强1,牛凌坤2
目的:探究早期游泳运动对哺乳期过度喂养肥胖大鼠(OF大鼠)内脏脂肪积累和食欲相关蛋白表达的长期影响,为运动防治哺乳期过度喂养型肥胖提供理论依据。方法:80只SD大鼠妊娠后第3天,随机选取22只母鼠孕期低强度游泳运动至分娩结束(10 min/day,6 days/week),其余安静至分娩。全部母鼠分娩后第2天随机调整每窝子鼠喂养数目为4只或10只,断乳后按体重配比随机抽取8只OF大鼠进行7周递增游泳运动,其余大鼠7周安静至取材。最终获得实验分组为,孕期安静+断乳后7周安静正常喂养对照组(CNC2,n=8)、孕期安静+断乳后7周安静OF组(CSC2,n=8)、孕期游泳运动+断乳后7周安静OF组(ESC2,n=8)和孕期游泳运动+断乳后7周游泳运动OF组(ESE1,n=8)。禁食后次日取材,采用免疫组化检测下丘脑弓状核NPY和POMC蛋白表达,放射免疫法测定血清insulin、leptin;酶化学法检测血糖和血脂;流式细胞术检测腹后壁脂肪细胞凋亡指数和脂肪细胞数目。结果:1)体成分和摄食量:整个幼龄期ESC2组体重、内脏脂肪垫重和摄食量与CSC2组相比差异不具显著性(P>0.05);ESE1组的体重、内脏脂肪垫重均显著低于CSC2组(P<0.05)。2)血清指标: ESC2组的血清学指标与CSC2组相比差异均不具显著性(P>0.05);ESE1组FINS、IGR、leptin和TG指标均显著低于CSC2组OF大鼠(P<0.05)。3)食欲肽:ESE1组下丘脑弓状核NPY和POMC蛋白表达量显著高于CSC2组(P<0.01);ESC2组和CSC2组NPY和POMC蛋白表达量相比差异不具显著性(P>0.05)。4)脂肪细胞指标:与CSC2组相比,ESE1组腹后壁脂肪细胞凋亡指数显著升高(P<0.01),脂肪细胞数目则显著降低(P<0.01);ESC2组腹后壁脂肪细胞凋亡指数和细胞数目与CSC2组相比差异均无显著性(P>0.05)。结论:单纯孕期低强度游泳运动对哺乳期过度喂养型肥胖大鼠没有长期改善效果,孕期结合断乳后7周的早期游泳运动具有改善哺乳期过度喂养型大鼠肥胖的效果。早期游泳运动是否能引发中枢肽-循环激素-脂肪细胞三者协同变化是有效改善哺乳期大鼠过度喂养型肥胖的关键。
早期游泳运动;过度喂养大鼠;神经肽Y;阿片促黑素细胞皮质素原;内脏脂肪凋亡指数
编程作用假说(Programming Hypothesis)认为在生长发育的关键期如胚胎期和新生早期,发育环境的变化会对子代在基因、细胞和组织的结构和功能上产生类似计算机编程样的持久影响[18]。有关早期营养变化的编程作用机制研究已充分证实,机体对营养性应激或刺激的早期适应将持续改变机体的生理和代谢活动,同时,对生命早期食欲和能量调控系统发育起持久性作用[20]。早期营养暴露的时程性是至关重要的,哺乳期过度喂养可引发下丘脑食欲中枢功能异常、血瘦素和胰岛素升高,促进脂肪细胞的分化和增殖,对机体正常的神经-内分泌-免疫调控网络和脂肪细胞的发育产生持久影响,诱发儿童和青少年肥胖,并对成年后肥胖的发生产生类似计算机编程样的作用,即体重编程作用[18]。自McCance 等证实,在哺乳期通过调整每窝喂养仔鼠数目可诱发子代大鼠肥胖和糖、脂代谢异常并持续至成年期的现象后,过度喂养大鼠(the overfed rat,OF 大鼠)已成为研究哺乳期营养过剩诱发子代体重编程作用的常用动物模型[20]。
控制摄食和增加运动是临床医学和运动医学非药物治疗过度喂养所致儿童单纯性肥胖的主要手段,其中控制摄食量是治疗的关键。但是,由于儿童正处在生长发育的高峰期,控制摄食量在实际操作中很难操作和实现,这也是目前儿童单纯性过度喂养型肥胖治疗的瓶颈。因此,本实验主要从运动与内分泌、运动与下丘脑食欲肽、运动与内脏脂肪细胞凋亡3个角度,研究早期运动对10周龄过度喂养大鼠(OF大鼠)体重编程作用影响的效果和潜在机制,以期为运动防治哺乳期过度喂养型肥胖提供理论依据。
1 材料与方法
1.1 材料
1.1.1 实验动物与分组
饲养条件:SD纯系大鼠,雌性大鼠80只(275±5 g),雄鼠20只(350±5 g),购自河北省动物实验中心;室温23℃±2℃,湿度50%±5%,控制光周期,标准饲料饲养。
OF大鼠模型复制:雌雄大鼠按 2∶1 比例合笼,次日晨起以精栓出现或阴道涂片确定妊娠。母鼠分娩后次日,选取每窝产仔数目10~13只的母鼠及其仔鼠,然后依据仔鼠生殖突与肛门间距离甄别性别,调整每窝喂养仔鼠数目。过度喂养为每窝 4 只雌鼠和正常喂养为每窝 10 只雌鼠,直至哺乳期21天后断乳[15,16]。在整个妊娠期和哺乳期,母鼠均为标准饲料喂养,断乳后7周全部OF大鼠和正常喂养大鼠均为标准饲料喂养。
实验分组:80只SD大鼠妊娠后第3天,随机选取22只母鼠孕期低强度游泳运动至分娩结束,其余安静至分娩。母鼠分娩后次日随机调整每窝子鼠喂养数目为4只或10只,进行21天哺乳期安静喂养。断乳后按体重配比随机抽取8只OF大鼠进行7周递增游泳运动,其余大鼠7周安静至取材。最终获得实验分组为,孕期安静+断乳后7周安静正常喂养对照组(CNC2,n=8)、孕期安静+断乳后7周安静OF组(CSC2,n=8)、孕期游泳运动+断乳后7周安静OF组(ESC2,n=8)和孕期游泳运动+断乳后7周游泳运动OF组(ESE1,n=8)。
1.1.2 早期游泳运动方案
母鼠孕期运动方案:10 min/day,6 days/week[19,21]。游泳设备为150 cm×50 cm×90 cm游泳池,水温34℃±2℃。
断乳后7周运动方案:每周运动6天,每天1次;第1周为30 min/day,每周递增15 min,至第3周为60 min, 每周递增5 min,至第 7周为90 min/day[24],总计运动7周。
1.1.3 标本采集
取材:称重后麻醉量身长和尾长,腹主动脉取血5 ml制备血清;取子宫旁脂肪垫(parametrial fat pad,PARA)、肾周脂肪垫(perirenal fat pad,PERI)和腹后壁脂肪垫(retroperitoneal fat pad,RP)称重;取右侧RP脂肪700 mg,制备单细胞悬液。升主动脉多聚甲醛灌流液后,取自视交叉至乳头体区域4%甲醛溶液固定。
1.2 方法
1.2.1 摄食量、体重检测和Lee指数计算
每日下午称量体重和余食物量。每日摄量=前天投食量-次日余食量。
取材日测量大鼠鼻尖至肛门的距离即身长和体重。

1.2.2 血清学指标检测
血糖和血脂采用酶化学法在全自动分析仪上测定;血清胰岛素和瘦素采用放射免疫法测定。胰岛素敏感性评价指标为胰岛素与血糖比(the ratio of insulin to glucose,IGR)= FINS值/FBS[10](FINS为空腹胰岛素值,FBS为空腹血糖值)。
1.2.3 免疫组化检测
常规石蜡切片(Leica电动切片仪),脑冠状切片(5 μm)经脱蜡和梯度酒精复水后,室温孵育20~30 min后,柠檬酸缓冲溶液水浴抗原修复。正常山羊血清封闭液,分别滴加兔抗大鼠的神经肽Y(NPY,1∶500,武汉博士德)、阿片促黑素细胞皮质素原抗体(POMC,1:800,美国Phoenix公司),湿盒内4℃,孵育48h。阴性对照组用0.01MPBS替代一抗。生物素化第二抗体(SP-9000,中杉金桥),37℃孵育30min,滴加辣根酶标记链霉卵白素工作液(S-A/HRP),37℃孵育30 min,DAB( ZLI-9032,中杉金桥)显色,苏木素复染,1%盐酸酒精分化、1%氨水反蓝、脱水透明后中性树脂封片。
1.2.4 脂肪细胞单细胞悬液制备和脂肪细胞数目及凋亡指数的检测
单细胞悬液制备[25,28]:将精确称量的700 mg腹后壁脂肪垫,剪为1 mm3大小的碎块,加入I型胶原蛋白酶(美国Sigma公司),37℃水浴90 min;加入等体积的DMEM培养液终止消化后,用240 μm尼龙沙网过滤后;200 g离心5 min,取上清液后;用吸管反复吹打,离心取上清,反复3次后获得的单细胞悬液,75%乙醇固定后待测。
脂肪细胞数目和凋亡指数的检测:在单细胞悬液中加入碘化丙啶(PI)(PI:50 mg/L,triton-×100 1.0%,美国Sigma公司),在4℃冰箱染色30 min。洗去染液,以200目铜网过滤后,采用Epics-XL II型流式细胞仪(检测前以鸡血红细胞作为标准样品调整仪器CV值在5%之内,激发光源为15 mW氩离子激光器,激发波长为488 nm)检测,然后Muticycle AV专用软件分析,通过计数凋亡区细胞数量得出凋亡指数。
1.2.5 统计学处理
2 实验结果
2.1 哺乳期过度喂养对大鼠的体重编程作用
由图1和图2显示,断乳后OF大鼠从3~10周龄的体重和摄食量、均显著高于对照组;10周龄OF大鼠体重、Lee指数、血清瘦素、胰岛素(FINS)、胰岛素与血糖比(IGR)和血清甘油三酯(TG)均显著高于对照组。
2.2 早期游泳运动对OF大鼠体重和摄食量的影响
图3显示,7至10周龄期间体重主效应显著(F7 wk=9.38,P<0.05;F8 wk=8.42,P<0.01;F9 wk=31.79,P<0.01;F10 wk=18.89,P<0.01),而4~10周龄期间摄食主效应均不显著。OF大鼠体重主效应显著的post hoc比较结果显示:7~8周龄ESE1组显著低于CSC2组和ESC2组(P<0.01)。

图 1 本研究不同喂养方式对幼龄期大鼠体重和摄食量的影响图
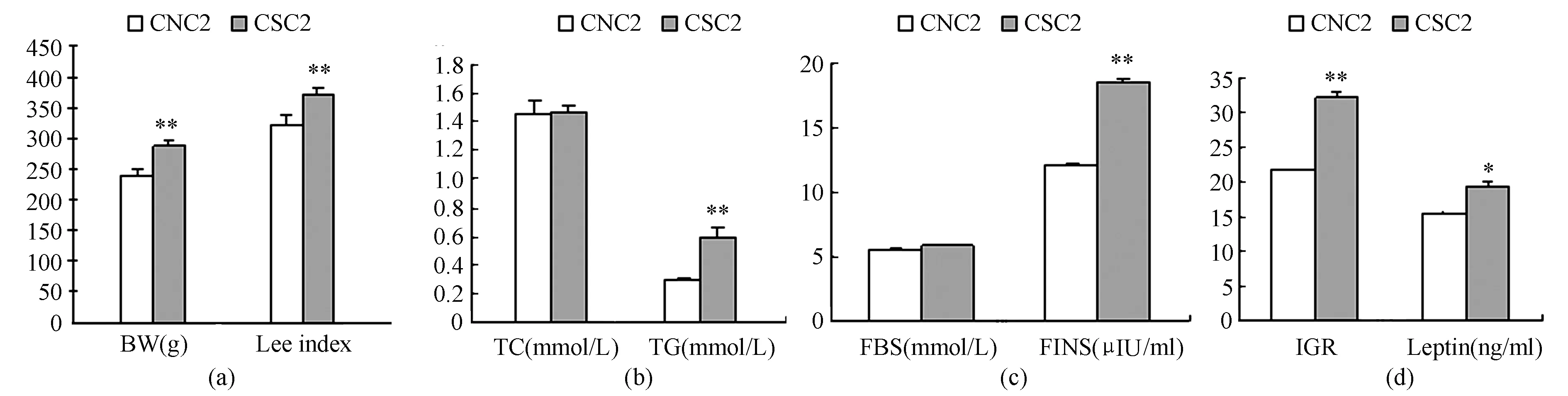
图 2 本研究不同喂养方式对10周龄OF大鼠体成分和血清学指标的比较图

图 3 早期游泳运动对幼龄OF大鼠体重和摄食量的影响图
2.3 早期游泳运动对10周龄OF大鼠体成分和血清学指标的影响
表1显示,ESC2组与CSC2组在体重、Lee指数、胰岛素、IGR、瘦素、总胆固醇(TC)和TG指标 差异均不具显著性(P>0.05);与CSC2组相比,ESE1组在体重、Lee指数、FINS、IGR、leptin和TG指标均显著降低(P<0.01,P<0.05,P<0.05,P<0.05,P<0.05,P<0.05)。
2.4 早期游泳运动对幼龄OF大鼠弓状核NPY和POMC蛋白表达的影响
图4显示,ESE1组下丘脑弓状核NPY蛋白表达量显著高于CSC2组(P<0.01);ESC2组和CSC2组NPY蛋白表达量相比差异不具显著性(P>0.05)。
图5显示,ESE1组下丘脑弓状核POMC蛋白表达量显著高于CSC2组(P<0.01);ESC2组和CSC2组POMC蛋白表达量相比差异不具显著性(P>0.05)。
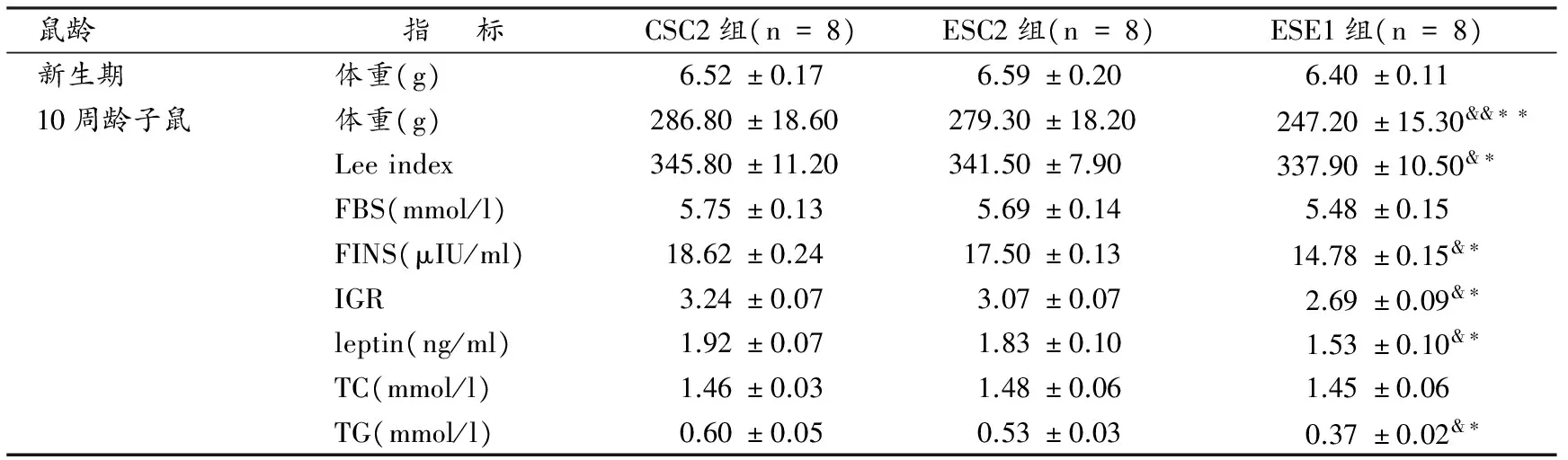
表 1 本研究早期游泳运动对10周龄OF大鼠体成分和血清学指标的影响一览表Table 1 Effects of Early Swimming on Body Composition and Serum Parameters in 10-week of Rats
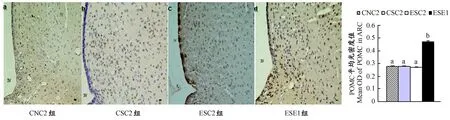
图 5 本研究下丘脑弓状核POMC蛋白表达的组间比较图
2.5 早期游泳运动对OF大鼠内脏脂肪积累的影响
早期运动对各组内脏脂肪垫重(肾周脂肪垫+腹后壁脂肪垫+子宫旁脂肪垫)和内脏脂肪垫重量百分比(内脏脂肪垫重/体重%)的差异比较显示(图6),ESC2组和CSC2组相比均无显著性差异(P>0.05);ESE1组则显著低于CSC2组(P<0.01);ESE1组与CNC2组相比,内脏脂肪积累无差异(P>0.05)。
2.6 早期游泳运动对OF大鼠腹后壁脂肪细胞凋亡和细胞数目的影响
图7显示,ESE1组腹后壁成熟脂肪细胞凋亡指数显著高于CSC2组(P<0.01),脂肪垫细胞数目显著低于CSC2组(P<0.01); ESC2组和CSC2组凋亡指数均显著低于CNC2组(P<0.01),而细胞数目显著高于CNC2组(P<0.01); ESC2组和CSC2组之间腹后壁成熟脂肪细胞凋亡指数和细胞数目差异均不具显著性(P>0.05)。
3 讨论与分析
哺乳期过度营养是婴幼儿超重肥胖、少年肥胖和成人肥胖发生的重要危险因素[22]。OF大鼠模型是研究早期过度喂养诱发持续肥胖的经典动物模型。OF大鼠是指通过在出生后当天或第2~3天调整每窝喂养仔鼠数目为3~4只直至哺乳期21天结束后获得的大鼠肥胖模型。借助该模型的研究显示,与正常喂养大鼠相比,OF大鼠体重和脂肪含量均升高10%~40%;OF大鼠出生后15天时体重显著高于正常喂养和大窝喂养大鼠并持续至成年,且其胰岛素水平也显著升高[23];OF大鼠在整个一生中均表现为过度摄食、糖耐量受损、TG水平升高、收缩压增高和超重与肥胖[14];雄性和雌性OF大鼠断乳后体重均显著高于正常喂养大鼠,而且,这种差异雄鼠比雌鼠更显著,出生后30天雄性和雌性OF大鼠均表现为脂肪细胞数目和体积的增加;但断乳后30~300天期间雄鼠主要表现为脂肪细胞体积增加,而雌鼠主要表现为脂肪细胞数目的增加,研究认为,脂肪组织形态的差异与脂肪细胞脂蛋白脂酶活性差异有关[12]。
本研究模型复制结果进一步证实(图1~图3,表1),雌性OF大鼠与正常喂养大鼠相比,在整个幼龄期均表现为过度摄食和体重超重/肥胖状态,及瘦素水平和血糖代谢异常的现象,并且OF大鼠内脏脂肪积累是由于内脏成熟脂肪细胞凋亡率降低和脂肪细胞数目增加引起(图6和7)。

图 6 本研究早期游泳运动对10周龄OF大鼠内脏脂肪积累的影响图
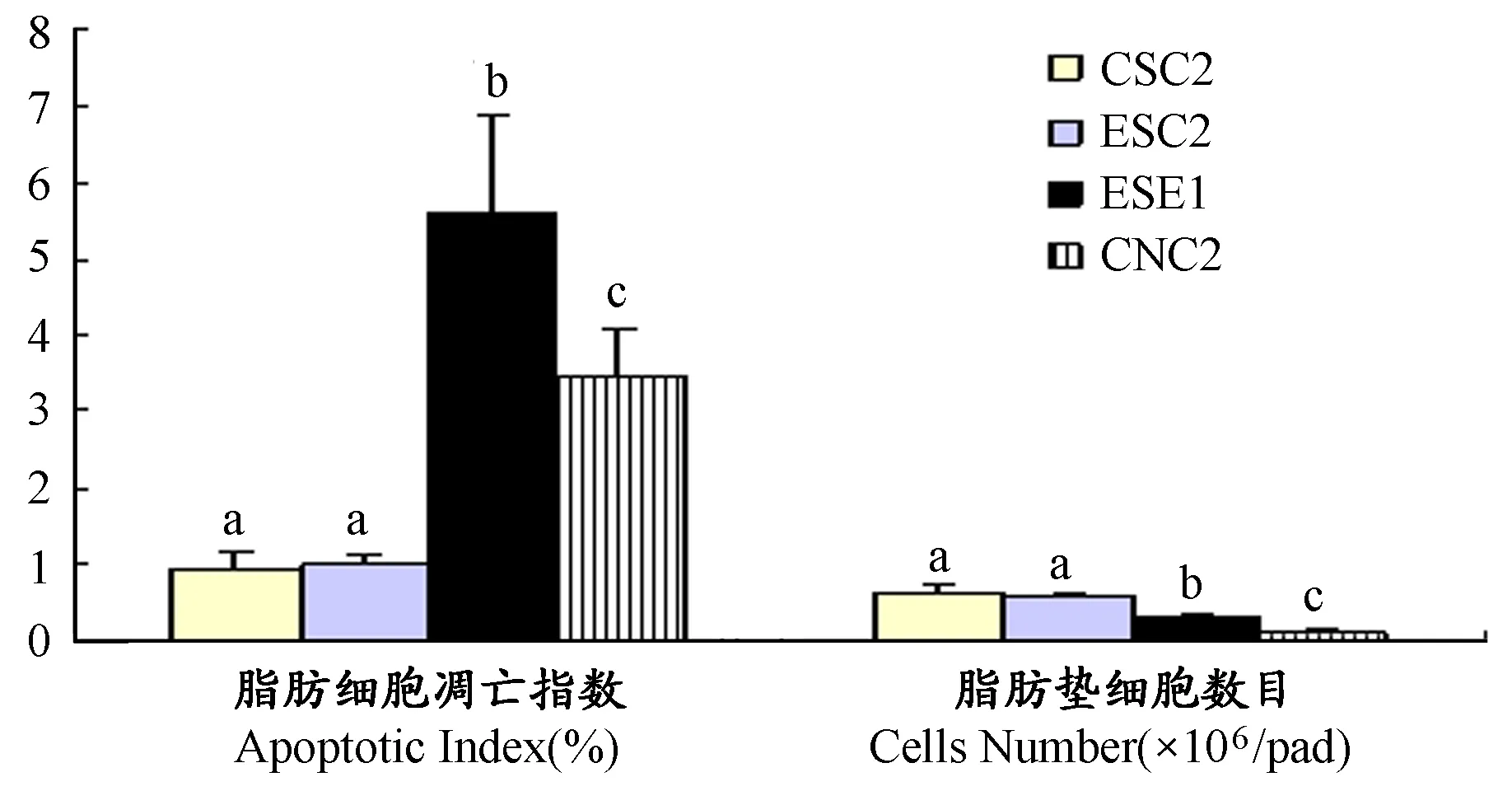
图 7 本研究早期游泳运动对腹后壁脂肪细胞凋亡指数和脂肪细胞数目的影响图
有关孕期运动对妊娠糖尿病大鼠的子代具有改善作用研究证实,孕期运动可通过改善母亲的胎盘机能,进而对子代胰脏功能产生改善作用[29]。孕期游泳运动是否会通过改善母亲胎盘机能,而优化子代发育的激素环境,使子代出生后面对营养过剩环境诱导具有预防作用呢?我们研究发现,孕期游泳运动对3周龄和10周龄OF大鼠的Lee指数、内脏脂肪积累和摄食量无显著改善作用(图3和表1)。究其原因,本研究孕期游泳运动方法为低强度短时间游泳运动,旨在增加孕鼠的身体活动水平,该运动方案对OF大鼠血瘦素和胰岛素水平异常无显著改善效果、对OF大鼠下丘脑食欲肽NPY表达无降低效果(图4),对内脏脂肪细胞数目和脂肪细胞凋亡率均无显著改善效果(图6,7),是孕期低强度游泳运动对出生后过度营养诱导肥胖无预防作用的潜在机制。可见,本研究所采用的经典孕期游泳运动方案[19,21],虽对断乳后OF大鼠神经反射、自主活动水平[1]和下丘脑POMC mRNA 表达和蛋白合成有显著改善效果[2],但是,对哺乳期过度喂养肥胖发生无显著的预防作用。可见,哺乳期3周的过度喂养可引发OF大鼠长达7周体重编程作用与早期过度营养导致的持久的摄食过度密切相关,经典孕期游泳运动方案虽然对子代中枢能量调控系统和学习记忆具有益的作用,但是单纯孕期低强度游泳运动不足以抵御哺乳期过度喂养诱发的大鼠持久地过度摄食行为。研究提示,体重正常的健康女性孕期适度运动是有益于子代的学习记忆和能量代谢中枢发育,但预防其子代幼龄期肥胖的关键是哺乳期科学喂养;能否引发“中枢肽-循环激素-脂肪细胞三者协同变化”是早期游泳运动防治哺乳期过度喂养型肥胖的关键。
肥胖的发生与脂肪细胞数量的极端异常增殖密切相关。与其他组织不同,脂肪组织具有体积无限增长的潜能,而且膳食诱导脂肪细胞数目的增加具有不可逆转性[16]。母亲妊娠期和哺乳期营养过剩可造成子代大鼠脂肪细胞肥大,还可永久改变脂肪细胞发育和脂解作用相关蛋白的表达,导致脂肪细胞增殖和肥大的永久性改变[9];相关研究显示,7周龄OF大鼠进行9周跑台运动可显著降低其体重和内脏脂肪垫重[13]。我们研究进一步说明,孕期结合断乳后7周的早期游泳锻炼可增加OF大鼠腹后壁脂肪细胞凋亡指数、降低腹后壁脂肪细胞数目,降低内脏脂肪垫重量。前脂肪细胞在生脂因子的刺激下增殖与分化实现脂肪细胞数目增加;其数目的减少则通过脂肪细胞、前脂肪细胞的凋亡及其他可能的机制,如脂肪细胞的去分化而实现。上述两个过程发生紊乱,均有可能导致肥胖发生[27]。机体可通过凋亡过程清除体内不需要或有害的细胞,或通过抗凋亡过程维持正常细胞功能。大量研究表明。脂肪细胞在体内外均可发生凋亡[26],但是,肥胖时脂肪细胞凋亡不足的确切机制以及在体内发生凋亡与中枢和内分调节之间的关系尚未阐明,有待于我们进一步的研究。
瘦素是脂肪组织与中枢能量稳态系统联系的外周信号之一。血瘦素水平通常与体重,尤其是脂肪组织的变化呈正相关。但临床上儿童肥胖症患者多表现为瘦素抵抗[8]。本研究进一步证实,哺乳期过度喂养诱导的肥胖OF大鼠为瘦素抵抗(图2)。相关研究表明,运动是改善机体血瘦水平的有效手段。长期有氧运动具有降低内脏脂肪垫含量,增加脂肪组织瘦素表达、提高血瘦素水平,降低NPY合成,使机体产热和能量消耗增加[3,6,7]; 8周中等强度游泳运动可显著改善糖尿病大鼠体脂和血瘦素含量[5]。本研究显示,单纯孕期低强度游泳运动对子代OF大鼠瘦素抵抗现象的产生无预防效果,但是,孕期结合断乳后7周的早期游泳锻炼可显著降低血瘦素水平,还具有降低OF大鼠内脏脂肪垫重和成熟脂肪细胞数目,提高脂肪细胞凋亡率,以及增加下丘脑NPY表达的作用 (表1,图4,图6,图7)。以上结果提示,血leptin水平随OF大鼠体脂积累减少也相应下降,血leptin水平的下降,经过leptin信号转导作用于下丘脑弓状核NPY神经元,增加了NPY蛋白表达,该信号从下丘脑弓状核“一级”食欲和能量调控靶点向“下一级”神经元传递,可以起到增加食物摄入,减少产热,降低能量消耗的反馈调节作用,这与成年后运动和单纯幼龄期运动对下丘脑NPY蛋白表达的影响相一致。但是,早期游泳运动对OF大鼠下丘脑弓状核NPY蛋白表达升高的影响,并没有引起OF大鼠摄食量增加和内脏脂肪积累上升(图3,图6),说明OF大鼠断乳后7周运动,不但对leptin-下丘脑NPY轴有显著影响,还对下丘脑其他食欲与能量代谢调控通路产生影响。
在弓状核中,约30%的POMC神经元可分泌表达瘦素受体,且瘦素对POMC mRNA的表达具有正向调节作用[11]。高瘦素水平,可诱导弓状核中POMC表达升高,减少进食[4]。本研究发现,孕期结合断乳后7周的早期游泳锻炼,OF大鼠与安静对照OF大鼠相比,体重下降、内脏脂肪积累下降、血瘦素水平下降、血胰岛素水平下降,OF大鼠弓状核POMC和NPY蛋白表达均显著增加。早期游泳运动虽然增加了OF大鼠下丘脑NPY蛋白表达水平,但是,OF大鼠未通过增加摄食量以补充运动引发的能量消耗,这可能与POMC蛋白表达增加有关。提示,运动最终是通过何种途径对这两种互相拮抗的中枢神经肽系统(NPY系统和黑皮素系统)产生影响,进而起到减低体重和减少内脏脂肪积累还有待进一步探究。
4 小结
单纯孕期低强度游泳运动对哺乳期过度喂养型肥胖大鼠没有长期改善效果,孕期结合断乳后7周的早期游泳运动具有改善哺乳期过度喂养型大鼠肥胖的效果。早期游泳运动是否能引发中枢肽-循环激素-脂肪细胞三者协同变化是有效改善哺乳期过度喂养型大鼠肥胖的关键。
研究提示,体重正常的健康女性孕期适度运动是有益于子代的学习记忆和能量代谢中枢发育,但预防其子代幼龄期肥胖的关键是哺乳期科学喂养;早期游泳运动治疗哺乳期过度喂养型肥胖的关键是,科学制订能引发“中枢肽-循环激素-脂肪细胞三者协同变化”的运动方案。
[1]高峰,牛凌坤,冯鹏,等.孕期低强度游泳运动对哺乳期过度喂养子代肥胖大鼠生长发育和糖脂代谢的影响.中国运动医学杂志,2010,29(6):677-682.
[2]高峰,何玉秀,郝强,等.孕期低强度游泳运动对哺乳期过度喂养子代肥胖大鼠下丘脑弓状核食欲相关基因表达的影响.中国体育科技,2012,48(6):123-127.
[3]何玉秀,白文忠,姚玉霞,等.运动对腹部脂肪积累及肥胖基因表达的影响.体育科学,1998,18(4):69-70.
[4]刘国庆,章孝荣,赵有璋.侧脑室注射Leptin对大鼠下丘脑前阿黑皮素mRNA表达的影响.中国兽医学报,2002,22(3) :265 -266.
[5]孙莉敏,吴毅,胡永善,等.耐力运动增加糖尿病大鼠血清瘦素水平的研究.中华物理医学与康复杂志,2003,18(2):72-74.
[6]张缨,冯美云,吴昊,等.有氧运动训练对大鼠下丘脑食欲调节肽的影响.中国运动医学杂志,2002,21(1):19-22.
[7]张缨,冯美云,吴昊.有氧运动训练对糖尿病大鼠Leptin-下丘脑NPY轴的影响.中国运动医学杂志 ,2002,21(3):239-243.
[8]AKANISHI T,U R,LIU ZJ,etal.Sexual dimorphism um leptin and relative weight for the standard in norm overweight children as well as adolescents.Eur J Clin Nutr,2001,55(11):899-993.
[9]BAYOL S A,SIMBI B H,STICKLAND N C.A maternal cafeteria diet during gestation and lactation promotes adiposity and impairs skeletal muscle development and metabolism in rat offspring at weaning .J Physiol,2005,567(3):951-961.
[10]CHENG T O.Fat kids grow up to be fat adults.A lesson to be learned from China .Int J Cardiol,2007,117(1):133-135.
[11]CHEUNG C C,CLIFTON D K,STEINER R A,etal.Proopiomelanocortin neurons are direct targets for leptin in the hypothalamus .Endocrinol,1997,138(4):4489-4492.
[12]CRYER A,JONES H M.The development of white adipose tissue.Effect of litter size on the lipoprotein lipase activity of four adipose-tissue depots,serum immunoreactive insulin and tissue cellularity during the first year of life in male and female rats .Biochem J,1980,186(3):805-815.
[13]DANTAS E M,PIMENTEL E B,GONCALVES C P,etal.Effects of chronic treadmill training on body mass gain and visceral fat accumulation in overfed rats.Braz J Med Biol Res,2010,43(5):515-521.
[14]DAVIDOWA H,Li Y ,PLAGEMANNN A.Altered responses to orexigenic (AGRP,MCH) and anorexigenic (alpha-MSH,CART) neuropeptides of paraventricular hypothalamic neurons in early postnatally overfed rats .Eur J Neurosci,2003,18(3):613-621.
[15]EPSTEIN H T.The effect of litter size on weight gain in mice.J Nutr,1978(1):120-123.
[16]FAUST I M,JOHNSON P R, HIRSCH J.Long-term effects of early nutritional experience on the development of obesity in the rat .J Nutr,1980,110(10):2027-2034.
[17]FAUST I M,JOHNSON P R,STERN J S,etal.Diet-induced adipocyte number increase in adult rats:a new model of obesity .Am J Physiol Endocrinol Metab,1978,235(G279-G286):E279-E286.
[18]GARDER D S,RHODES P.Developmental origins of obesity:programming of food intake or physical activity? Adv Exp Med Biol,2009,646(1):83-93.
[19]HEE-HYUK L,HONG K,LEE J W,etal.Maternal swimming during pregnancy enhances short-term and neurogenesis in the hippocampus of rat pups.Brai Deve,2006,28(3):147-154.
[20]KIANIL A,NIELSEN M O.Metabolic programming:Origin of non-communicable diseases in early life nutrition .Int J Endocrinol Metab,2011,9(3):409-415.
[21]LOKEY E A,TRAN Z V,WELLS C L,etal.Effects of physical exercise on pregnancy outcomes:a meta-analytic review .Med Sci Sports Exe,1991(11),23:1234-1239.
[22]MARTORELL R,STEIN A D ,SCHROEDER D G.Early nutrition and later adiposity .J Nutr,2001,131(3):874S- 880S.
[23]PLAGEMANN A,HARDER T,RAKE A,etal.Increased number of galanin-neurons in the paraventricular hypothalamic nucleus of neonatally overfed weanling rats .Brain Res,1999,818(1):160-163.
[24]PLOUG T,STALLKENCHT B M,PEDERSEN O,etal.Effect of endurance training on glucose transport capacity and glucose transporter expression in rat skeletal muscle .Am J Physiol,1990,259(6Pt1):E778-E786.
[25]POULOOS S P,MATTHEW S.Pre-and postnatal dietary conjugated linoleic acid alters adipose development,body weight gain and body composition in sprague-dawley rats .J Nutr,2001,131(10):2722-2731.
[26]PRINS J B,NIESLER C U,WINTERFORD C M,etal.Tumor necrosis factor α induces apoptosis of human adipose cells .Diabetes,1997,46(12):1939-1944.
[27]PRINS J B,STEPHEN O R.Regulation of adipose cell number in man.Clin Sci,1997,92(1):3-11.
[28]SAMAD F,PANDEY M,LOSKUTOFF D J.Tissue factor gene expression in the adipose tissues of obese mice.Proc Natl Acad Sci,USA,1998,95(13):7591-7596.
[29]VANHEEST J C ,RODGERS C D.Effects of exercise in diabetic rats before and during gestation on maternal and neonatal outcomes.Am J Physiol Endocrinol Metab,1997,273(4 Pt 1):E727-E733.
EffectofEarlySwimmingExerciseontheExpressionsofAppetite-RelatedPeptidesandVisceralFatAccumulationinOverfedObeseRats
GAO Feng1,HE Yu-xiu1,HAO Qiang1,NIU Ling-kun2
Objective:To investigate whether early swimming exercise has long-term effects on the appetite peptides of hypothalamus and visceral fat accumulation in overfed rats.Methods:Total 80 SD pregnant rats were enrolled,among which 22 rats began swimming(10min per day,6days per week)from the 3rd day of pregnancy to delivery .The rest rats kept sedentary till delivery.The pups were distributed randomly and assigned to small litters (4 pups) and normal litters (10 pups) on the second day after birth.8 overfed pups were randomly selected and assigned to swim 7-week graded swimming training after weanling,the rest kept sedentary.Then we had 4 groups:the pregnancy sedentary+normal little control group(CNC2,n=8),the pregnancy sedentary+ OF control group(CSC2,n=8),the pregnancy swimming + OF group(ESC2,n=8)and pregnancy swimming + post weanling 7wks swimming OF group(ESE1,n=8).At the 10 weeks of age the animals were starved overnight and were decapitated in the next day.NPY and POMC in the arcuate nucleus of hypothalamus were analyzed with immunohistochemistrical technique.The fasting serum insulin and leptin were assayed by radioimmunoassay,the fasting serum glucose and lipids were assayed by enzymatic methods using commercial kits.The apoptotic index and the number of adipocyte in retroperitoneal (RP) fat pad were detected by flow cytometric analysis.Results:1)Body composition and food consumption:No significant (P>0.05) differences were found between the Group ESC2 and CSC2 (P>0.05) with regard to the body weight,internal fat mass and the food consumption per day at the 10 weeks of age.The body weight,internal fat mass and the food consumption per day were significantly lower in Group ESE1 than that was in Group CSC2(P<0.01).2)Serum parameters:No significant (P>0.05) differences were found between Group ESC2 and Group CSC2 with regard to the Serum parameters at the 10 weeks of age.The serum levels of FIN,IGR,leptin and TG were significantly lower in Group ESE1 than those in Group CSC2 (P<0.01).3)Appetite-Related peptides:The positive expressions of NPY and POMC in the arcuate nucleus of hypothalamus were obviously higher in Group ESE1 than those in Group CSC2 (P<0.01).The positive expressions of NPY and POMC were not significant differences in Group ESC2 than those in Group CSC2 (P<0.01).4)Adipocyte parameters:RP fat pad apoptotic index was obviously higher in Group ESE1 than that in Group CSC2 (P<0.01).While the number of RP adipocyte was obviously lower in Group ESE1 than that in Group CSC2 (P<0.01).Conclusion:Maternal swimming itself during pregnancy in order to increasing the level of physical activity has no long-term prevention effects on the interval fat accumulation and the body weight programming in the overfed rats;The early swimming during the pregnancy and juvenile could improve the overfed obesity.The synergistic regulation among the central peptides,circulation hormones and the adipocyte is the key for the early swimming lasting improving the overfed obesity.
earlyswimmingexercise;overfedrat;NPY;POMC;visceralfat;apoptoticindex
2013-06-03;
:2013-12-21
国家自然科学基金(30871215),河北省重点学科项目(20130227);河北师范大学博士基金资助。
高峰(1972-),女,湖北松滋人,高级实验师,博士,硕士研究生导师,研究方向为运动行为与健康促进,Tel:0311-80787735,Email:gaolindy@126.com;何玉秀(1964-),女,浙江临安人,教授,博士,博士研究生导师,研究方向为运动生理学,Tel:(0311)80787702,E-mail:heyuxiu@mail.hebtu.edu.cn。
1.河北师范大学 省重点实验室,河北 石家庄 050024;2.河北传媒学院 艺术体育系,河北 石家庄 050071 1.Hebei Normal University,Shijiazhuang,050024,China;2.Department of Sports and Art,Hebei Institute of Communication,Shijiazhuang, 050071,China.
1002-9826(2014)02-0038-08
G861.101.7
:A
猜你喜欢
癌症进展(2022年18期)2022-11-26
基层中医药(2022年7期)2022-11-17
实用老年医学(2021年10期)2021-12-05
现代畜牧科技(2021年4期)2021-07-21
健康之家(2021年19期)2021-05-23
科学与财富(2021年26期)2021-03-04
人人健康(2018年8期)2018-08-23
科学种养(2017年11期)2017-11-28
农家顾问(2016年6期)2016-05-14
商情(2009年17期)2009-09-23
