
南亚热带中幼龄针阔混交林碳储量及其分配格局
2014-07-16 05:40:32周丽张卫强唐洪辉陈伟光魏丹盘李军苏木荣
生态环境学报 2014年4期
周丽,张卫强*,唐洪辉,陈伟光,魏丹,盘李军,苏木荣
1. 广东省林业科学研究院, 广东 广州 5105201;2. 广东省佛山市云勇生态林养护中心, 广东 佛山 5285181
杉木(Cunninghamia lanceolata)是我国南方亚热带地区主要的造林树种之一,近几十年来,由于经济利益的驱动,大面积天然林被具有高经济价值的杉木人工林所取代(万晓华等,2013)。由于长期连栽杉木,导致杉木连栽地力下降(俞新妥等,1989;杨承栋等,1996),生物多样性减少(周霆等,2008)等生态问题。为了减少杉木人工纯林所带来的不利影响,乡土阔叶树种已经成为杉木林改造的主要造林树种,但对于乡土阔叶树种用于改建人工林后,它们所在生态系统各组分碳储量及分配格局的研究还比较缺乏(明安刚等,2012;王卫霞等,2013;刘恩等,2012;何斌等,2009),尤其是中幼龄针阔混交林生态系统各组分碳储量及分配特征研究还缺少系统的、详细的研究。碳是构成森林生态系统组分和影响森林生态功能发挥的重要元素,对生态系统的生产力、固碳潜力以及稳定性都具有关键的影响作用(Reich,2006)。本研究探讨了我国南亚热带杉木皆伐迹地阔叶树种造林对森林生态系统碳储量的影响,分析了中幼龄针阔混交林碳储量及其分配格局,旨在为该区人工林营建中的树种选择、人工林生态系统生产力和固碳潜力、人工碳汇林的经营管理研究提供科学参考。
1 研究区概况
研究区位于广东省佛山市高明区云勇林场,东经112°40′,北纬22°43′。属于南亚热带湿润季风气候。气候温和,年平均气温、最高气温和最低气温分别为22.0、34.5和3.5 ℃,雨量充沛,年降水量平均达2000 mm,集中在4-8月(薛立等,2008)。地势属丘陵地带,土壤为花岗岩发育的酸性赤红壤,土层深厚。从2002-2011年,采用皆伐造林方式对杉木林进行改造,造林树种大多选用乡土阔叶树种,杉木萌芽条只保留单株,经过多年改造,形成了针阔混交林,针阔混交林研究样地见表1。主要造林树种为红花荷(Rhodoleia championii)、火焰木(Spathodea campanulat)、火力楠(Michelia macclurei)、华南毛柃(Eurya ciliata)、米老排(Mytilaria laosensis)、阴香(Cinnamomum burmannii)、山杜英(Elaeocarpus sylvestris)、枫香(Liquidambar formosana)、香樟(Cinnamomum camphora)、格木(Erythrophleum fordii)、木荷(Schima superba)、红荷(Schima wallichii)、藜蒴(Castanopsis fissa)、楝叶吴茱萸(Evodia meliaefoli)、假苹婆(Sterculia lanceolata)、香椿(Toona sinensis)、黄樟(Cinnamomum porrectum)、乌桕(Sapium sebiferum)、小叶青冈(Cyclobalanopsis myrsinaefolia)、非洲桃花心(Khaya senegalensis)和灰木莲(Manglietia glauca)等。主要林下植被包括五指毛桃(Radix Fici)、鸭脚木(Schefflera heptaphylla)、三丫苦(Evodia lepta)、野牡丹(Melasroma candidum)、毛果算盘子(Glochidion eriocarpum)、猴耳环(Pithecellobium clypearia)、梅叶冬青(Ilex asprella)、山苍子(Litsea cubeba)、玉叶金花(Mussaenda pubescens)、黑面神(Breynia fruticosa)、鲫鱼胆(Maesa perlarius)、蔓生秀竹(Microstegium fasciculatum)、弓果黍(Cyrtococcum patens)、火炭母(Persicaria chinensis)、蒲公英(Taraxacum mongolicum)、乌毛蕨(Blechnum orientale)、半边旗(Pteris semipinnata)和铁线蕨(Adiantum flabellulatum)等。

表1 研究样地概况Table 1 Survey of sample plots
2 研究方法
2.1 乔木生物量测定
2013年10月,选择地形地貌、海拔、母岩、土壤类型等相同或相近,且彼此相连的3~5、7~9和10~11 a林龄段针阔混交林作为试验样地,系统布点,设立乔木、灌木、草本固定调查样方。在每个林龄段针阔混交林分内设计4个固定样方,样方规格为20 m×20 m,调查1.5 m(含1.5 m)以上乔木和灌木。在每木检尺的基础上,选择主要造林树种标准木3株,将标准木解析,现场测定干、枝、叶的鲜质量,地下部分采用“全挖法”测定根鲜质量,各器官取分析样品200 g,带回实验室在80 ℃下烘干至恒质量,将各器官鲜质量换算成生物量。各样地乔木的生物量测算采用已有生物量相对生长方程和样地调查数据相结合的方法(王晓丽等,2013)进行。大量研究表明,乔木各器官及森林总生物量与测树因子间普遍存在着相关关系,这种相关关系可以用数学模型进行拟合,如米老排(明安刚等,2012)、红椎(刘恩等,2012)、杉木(俞月凤等,2013)、樟树(姚迎九等,2003)、木荷(曾小平等,2008)。根据生物量预测模型直接估算林分叶、枝、干和根系等器官的生物量,其他常绿阔叶树种生物量参考文献计算求得(李静鹏等,2013;杨昆等,2007)。
2.2 林下植被生物量及凋落物现存量测定
按“梅花五点法”在每个固定样地布设5块2 m×2 m小样方,进行林下灌木和草本的常规调查,在样地外选择与小样方相似的地段,采用全部收获法收集灌木和草本,分地上、地下部分现场称鲜质量;在样地内2 m×2 m小样方收集全部凋落物;对灌木、草本以及凋落物进行取样,带回实验室在85 ℃的恒温箱中烘干至恒质量,由干/鲜质量比计算含水率,从而推算出林下植被生物量和凋落物现存量。
2.3 样品采集及碳含量测定
2.3.1 植物样品
乔木层树杆、树枝、树叶和树根分别取样。在距离地面1.3 m处取树主干样品;枝从粗到细按比例进行混合取样;叶按照叶片大小和树冠不同层次混合取样;根分粗(>5 mm)、中(2~5 mm)、细(<2 mm)混合取样;灌木分别取地上和地下部分的混合样品,而草本取地上和地下部分的混合样品;地表凋落物层按分解和半分解混合采样。采用重铬酸钾-水合加热法测定全碳含量。
2.3.2 土壤样品
森林土壤有机碳主要分布于土壤层1 m深度以内,是现阶段土壤碳库研究的重点(Dixon,1994)。在样地内有代表性的部位,分别挖取3个土壤剖面,按0 ~25、25 ~50、50 ~75和75~100 cm土层分层用100 cm3环刀取样测定土壤容重,同时取土壤样品,每层3个重复,同层土壤取混合样约200 g,分别装入自封袋中并做好标记。将土壤样品带回实验室风干,磨碎,过筛(孔径为0.25 mm),以重铬酸钾-水合加热法测定土壤有机碳含量。
2.4 森林生态系统各组分碳储量计算
乔木层各器官(干、枝、叶、根)、林下植被(灌木、草本)和凋落物层碳储量以其生物量现存量乘以相应的碳含量求得。
土壤层碳储量的计算采用如下公式:
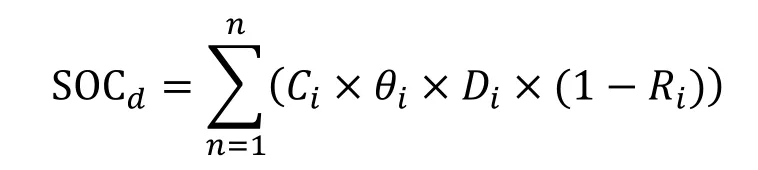
式中:SOCd为土壤有机碳密度(t·hm-2);n为土层数;Ci为第i层土壤的有机碳含量(%);θi为第i层土壤容重(g·cm-3);Di为第i层土壤厚度(m);Ri(%)为第i层土壤中直径大于2 mm石砾体积含量百分比。
2.5 数据处理
运用Excel将数据进行整理和预处理,利用SPSS 16.0进行单因素方差(one-way ANOVA)和主成分分析。采用Duncan新复极差法比较各指标在各观测样地的差异性,极显著性水平设为α=0.01,显著性水平设为α=0.05。
3 结果与分析
3.1 树种各器官碳含量
从表2可知,主要造林树种树根、树杆、树枝和树叶碳含量变化范围分别为42.20%~49.88%、44.09%~50.21%、44.96%~49.11%、43.83%~50.44%,杉木各器官碳含量最高。除山杜英和盆架子外,其余树种叶片碳含量最高,从而显示出叶片作为光合作用的主要器官,在碳素同化中的重要性(张治等,2014),樟树、黄樟、红椎、火力楠、木荷、红花荷树叶碳含量显著高于其树根碳含量(P<0.05),而与其树杆、树枝碳含量差异不显著(P>0.05)。杉木、米老排、阴香、观光木和盆架子树根、树杆、树枝和树叶间碳含量不存在显著差异(P>0.05)。从表3可知,灌木枝叶、灌木根系和草本碳含量间差异不显著(P>0.05),但显著低于凋落物碳含量(P<0.05)。林下植被和凋落物碳含量表现为凋落物>灌木枝叶>草本层>灌木根系。

表2 乔木各器官碳含量Table 2 The concentration of C in different organs of tree layers

表3 林下植被与凋落物碳含量Table 3 The concentration of C of undergrowth vegetation and litter
3.2 造林树种碳储量
对试验样地内树种进行每木检尺,选择主要造林树种标准木3株,将标准木伐倒,测定树根、树杆、树枝、树叶的生物量,参考乔木各器官碳含量,计算主要造林树种碳储量(表4)。从表4可知,11年生米老排平均胸径为15.27 cm,碳储量达54.21kg,为杉木林改造的速生树种;红花荷和盆架子生长缓慢,胸径为7.37和8.93 cm,碳储量仅为7.03和4.73kg,生长效果较差,不适宜作为杉木林改造树种。9年生红荷生长状况良好,其碳储量为17.39kg;5年生的红椎、观光木、黄樟、木荷和山杜英生长良好,其中,山杜英表现最为良好,其胸径达6.70 cm,碳储量达5.33kg。从树种各器官碳储量所占比例可知,红花荷、盆架子、香樟、黄樟根系碳储量占树种总碳储量比例较大,介于28.57%~31.59%之间,而红荷、火力楠、木荷和山杜英根系碳储量所占比例不足20%。除黄樟外,其余树种树杆碳储量所占树种总碳储量比例最大,其中,火力楠树杆所占比例达64.57%。树叶碳储量最小,其所占比例范围仅为3.50%~13.74%。
3.3 植被碳储量
从表5可知,10~11年林龄针阔混交林植被总碳储量为33.23 t·hm-2,7~9年林龄针阔混交林为26.90 t·hm-2,而3~5年林龄仅为7.10 t·hm-2。不同林龄段乔木和灌木碳储量均值随林龄的增加呈上升趋势,与植被碳储量变化趋势一致,而草本碳储量呈下降趋势。造林初期,林下灌木碳储量增加幅度远低于乔木,这主要由于造林初期幼林抚育,对灌木和草本碳储量影响较大,进入7~9年和10~11年林龄后,乔木生长迅速,一定程度挤占了林下灌木营养空间和生长空间,致使灌木碳储量增幅缓慢,乔木碳储量成为植被碳储量的主体。不同林龄段乔木碳储量占植被总碳储量比例分别为94.08%(10~11年生)、93.75%(7~9年生)和63.38%(3~5年生),灌木碳储量占植被总碳储量比例介于3.55%~12.67%之间,草本碳储量所占比例范围为1.28%~23.95%。

表4 乔木各器官碳储量Table 4 The carbon storage in different organs of tree

表5 针阔混交林植被碳储量Table 5 The carbon storage of vegetation in conifer-broadleaf mixed forests
3.4 土壤碳储量
从表6可知,不同林龄针阔混交林土壤碳储量均值间差异不显著(P>0.05),但随林龄的增加呈现出先降低后升高的趋势,这可能由于造林后的频繁抚育加速了土壤有机质的分解,但有机肥料投入量大,弥补了土壤有机碳损失,土壤有机碳含量维持在较高的水平,进入7~9年林龄后,植被层的生物量和土壤表层凋落物量都较小,凋落物与死亡根系的输入量不足以维持3~5年林龄土壤有机碳水平,土壤碳储量持续下降,随着乔木快速生长和林下植被的恢复,林地凋落物输入量明显增加,造林后第10~11年土壤有机碳储量升高,但仅为与3~5年林龄针阔混交林土壤碳储量的92.09 %。不同林龄针阔混交林土壤碳储量随土层深度的增加呈逐渐下降的趋势,在0-25 cm土壤表层内土壤碳储量显著大于其他土层(P<0.05),分别占整个土壤碳储量的40.39%、36.62%和37.14%,这主要由于枯枝落叶分解后输入土壤的有机碳都聚集在土壤表层,土壤有机碳的表聚性较明显(马和平等,2013)。随土层厚度的增加,差异减弱,50-75和75-100 cm土层内碳储量差异已不显著(P>0.05)。
3.5 针阔混交林碳储量及分配格局
针阔混交林碳储量及分配格局见表7。从表7可知,不同林龄针阔混交林总碳储量均值介于134.79~162.60 t·hm-2之间,针阔混交林碳库主要分为3个部分:植被层、凋落物层和土壤层,其碳储量分配格局表现为土壤层>植被层>凋落物层。研究发现,不同林龄针阔混交林植被层和凋落物层碳储量均值随林龄的增加呈上升的趋势;受土壤碳储量的影响,不同林龄针阔混交林总碳储量随林龄的增加表现为先降低后升高。土壤层0-100 cm 碳储量均值介于106.73~136.61 t·hm-2之间,占总储量的78.34%~94.45%,土壤层碳储量所占总碳储量比例随林龄的增加呈下降趋势,但中幼龄针阔混交林土壤层碳储量贡献最大。植被层的碳储量介于7.10~33.21 t·hm-2之间,占总碳储量的4.84 %~20.16%,其固碳能力远小于土壤层。凋落物层碳储量介于0.99~2.50 t·hm-2之间,仅0.71%~1.50%,凋落物层的碳储量虽较小,但它是土壤-植物系统碳循环的连接库,对森林生态系统的碳循环起到重要的作用(程小琴等,2012)。
4 讨论与结论
4.1 讨论
1)主要造林树种树根、树杆、树枝和树叶碳含量变化范围分别为42.20%~49.88%、44.09%~50.21%、44.96%~49.11%和43.83%~50.44%,这与王卫霞等(2013)和宫超等(2011)研究结果基本一致。人工林生态系统植被层各层次碳含量有较明显的层次规律(王卫霞等,2013),乔木层、灌木层、草本层的平均碳含量表现出随层片所在高度降低而减小的趋势(张治等,2014),表现为乔木层>灌木层>草本层,表现出随植物个体高度或组织木质化程度的降低,其碳素含量相应减少的趋势(李铭红等,1996)。林下植被和凋落物碳含量表现为凋落物>灌木枝叶>草本>灌木根系。乔木和灌木碳储量随林龄的增加呈上升趋势,而草本碳储量呈下降趋势,林下灌木碳储量增加幅度远低于乔木,这主要由于林下植被抚育措施在林分生长初期有利于造林树种的生长和生物量的积累,进而促进乔木有机碳的积累(吴亚丛等,2013)。

表6 针阔混交林土壤碳储量Table 6 The carbon storage of soil in conifer-broadleaf mixed forests t·hm-2

表7 针阔混交林碳储量及分配格局Table 7 The carbon storage and their allocation of conifer-broadleaf mixed forests
2)针阔混交林生态系统碳储量空间分布序列为土壤层>植被层>凋落物层,这种分配格局与王卫霞等(2013)和Finér等(2003)的研究结果基本一致。本研究中不同林龄针阔混交林地下部分与地上部分碳储量分别为3.55:1、3.80:1和16.87:1,远高于22年生人工林-杉木纯林1.53:1和杉木-火力楠混交林1.55:1研究结果(黄宇等,2005),这主要是由于中幼龄针阔混交林碳储量主要以土壤碳储量为主。中幼龄针阔混交林总碳储量均值介于134.79~162.60 t·hm-2之间,低于我国森林生态系统平均碳储量258.82 t·hm-2(周玉荣等,2000),土壤层碳储量均值介于106.73~136.61 t·hm-2之间,与广东鹤山马占相思林土壤层碳储量114.62 t·hm-2(李跃林等,2004)基本一致,但低于我国森林土壤平均碳储量201.76 t·hm-2(周玉荣等,2000),这主要由于土壤有机C的积累有影响南亚热带地区, 全年热量丰富, 雨量充沛, 生物循环旺盛, 有机物质代谢快, 不利于土壤有机质的积累(黄宇等,2005)。
4.2 结论
1)主要造林树种树根、树杆、树枝和树叶碳含量变化范围分别为42.20%~49.88%、44.09%~50.21%、44.96%~49.11%、43.83%~50.44%,植物层次平均碳含量表现为乔木>灌木>草本。乔木碳储量为植被碳储量的主体,占植被总碳储量比例介于63.38%~94.08%,灌木碳储量所占比例介于3.55%~12.67%之间,而草本碳储量仅介于1.28%~23.95%。
2)中幼龄针阔混交林土壤碳储量介于106.73~136.61 t·hm-2之间,土壤碳储量随土层深度的增加呈逐渐下降的趋势,土壤碳储量主要集中在0-50 cm土层内,分别占整个土壤碳储量的65.40%、64.02%和62.55%,而在0-25 cm土层内碳储量显著大于其他土层(P<0.05),随土层厚度的增加,差异减弱。
3)中幼龄针阔混交林总碳储量均值介于134.79~162.60 t·hm-2之间,其碳储量分配格局表现为土壤层>植被层>凋落物层。不同林龄针阔混交林植被层和凋落物层碳储量均值随林龄的增加呈上升的趋势;受土壤碳储量的影响,不同林龄针阔混交林总碳储量随林龄的增加表现为先降低后升高。土壤层碳储量贡献最大,占总储量的78.34%~94.45%。植被层的碳储量介于7.10~33.21 t·hm-2之间,占总碳储量的4.84%~20.16%,其固碳能力远小于土壤层。凋落物层碳储量介于0.99~2.50 t·hm-2之间,仅占0.71%~1.50%。
Dixon R K, Solomon A M, Brown S, et al. 1994. Carbon pools and flux of global forest ecosystems[J]. Science, 263: 185-190.
Finér L, Mannerkoski H, Piirainen S, et al. 2003. Carbon and nitrogen pools in an old-growth, Norway spruce mixed forest in eastern Finland andchanges associated with clear-cutting[J]. Forest Ecology and Management, 174(1/3) : 51-63.
Reich P B, Hobbie S E, Lee T, et al. 2006. Nitrogen limitation constrains sustainability of ecosystem response to CO2[J]. Nature, 440(7086):922-925.
曾小平,蔡锡安,赵平,等. 2008. 南亚热带丘陵 3 种人工林群落的生物量及净初级生产力[J]. 北京林业大学学报, 30(6):148-152.
程小琴,韩海荣,康峰峰. 2012. 山西油松人工林生态系统生物量、碳积累及其分布[J]. 生态学杂志, 31(10): 2455-2460.
宫超,汪思龙,曾掌权,等. 2011. 中亚热带常绿阔叶林不同演替阶段碳储量与格局特征[J]. 生态学杂志, 30( 9) : 1935-1941.
何斌,刘运华,余浩光,等. 2009. 南宁马占相思人工林生态系统碳素密度与贮量[J]. 林业科学, 45(2): 6-11.
黄宇,冯宗炜,汪思龙,等. 2005. 杉木、火力楠纯林及其混交林生态系统C、N 贮量[J]. 生态学报, 25(12) : 3146-3154.
李静鹏,徐明锋,苏志尧,等. 2013. 小尺度林分碳密度与碳储量研究[J].华南农业大学学报, 34(2):213-218.
李铭红,于明坚,陈启瑺,等. 1996. 青冈常绿阔叶林的碳素动态[J]. 生态学报, 16(6): 645-651.
李跃林,胡成志,张云,等. 2004. 几种人工林土壤碳储量研究[J]. 福建林业科技, 31(4) : 4-7.
刘恩,王晖,刘世荣. 2012. 南亚热带不同林龄红锥人工林碳贮量与碳固定特征[J]. 应用生态学报, 23(2): 335-340.
马和平,郭其强,刘合满,等. 2013. 藏东南色季拉山西坡土壤有机碳库研究[J]. 生态学报, 33(10): 3122-3128.
明安刚,贾宏炎,陶怡,等. 2012. 桂西南28年生米老排人工林生物量及其分配特征[J]. 生态学杂志, 31(5):1050-1056.
万晓华,黄志群,何宗明,等. 2013. 阔叶和杉木人工林对土壤碳氮库的影响比较[J]. 应用生态学报, 24(2): 345-350.
王卫霞,史作民,罗达,等. 2013. 我国南亚热带几种人工林生态系统碳氮储量[J]. 生态学报, 33( 3) : 0925-0933.
王晓丽,王媛,石洪华,等. 2013. 山东省长岛县南长山岛黑松和刺槐人工林的碳储量[J]. 应用生态学报, 24(5): 1263-1268.
吴亚丛,李正才,程彩芳,等. 2013. 林下植被抚育对樟人工林生态系统碳储量的影响[J]. 植物生态学报, 37(2): 142-149.
薛立,傅静丹,郑卫国, 等. 2008. 3种人工幼林的土壤微生物和酶活性研究[J]. 中南林业科技大学学报, 28(4):98-100.
杨承栋, 张小泉, 焦如珍, 等. 1996. 杉木连栽土壤组成, 结构, 性质变化及其对林木生长的影响[J]. 林业科学, 32(2): 175-181.
杨昆,管东生. 2007. 珠江三角洲地区森林生物量及其动态[J]. 应用生态学报, 18(4):707-712.
姚迎九,康文星,田大伦. 2003. 18 年生樟树人工林生物量的结构与分布[J]. 中南林学院学报, 23(1):1-5.
俞新妥, 张其水. 1989. 杉木连栽林地土壤生化特性及土壤肥力的研究[J]. 福建林学院学报, 9(3): 263-271.
俞月凤,宋同清,曾馥平,等. 2013. 杉木人工林生物量及其分配的动态变化[J]. 生态学杂志, 32(7) : 1660-1666.
张治,钟全林,程栋梁,等.2014. 闽西北地区不同龄组常绿阔叶混交林生态系统碳储量结构特征[J]. 生态环境学报, 23(2): 203-210.
周霆,盛炜彤. 2008. 关于我国人工林可持续问题[J]. 世界林业研究,21(3): 49-53.
周玉荣,于振良,赵士洞. 2000. 我国主要森林生态系统碳贮量和碳平衡[J]. 植物生态学报, 24(5): 518-522.
猜你喜欢
建筑与预算(2022年6期)2022-07-08 13:59:18
今日中国·西班牙文版(2021年4期)2021-04-09 11:22:46
防护林科技(2020年10期)2020-12-22 11:46:42
现代园艺(2018年2期)2018-03-15 08:01:03
防护林科技(2016年9期)2016-09-27 01:34:21
防护林科技(2016年5期)2016-09-05 01:19:43
广西林业科学(2016年1期)2016-03-20 05:33:02
广西林业科学(2016年4期)2016-03-16 05:44:58
亚热带资源与环境学报(2015年1期)2015-01-22 07:04:58
建筑与预算(2014年5期)2014-04-11 07:29:28
