
pH对鱼腥藻和普通小球藻生长竞争的影响
2014-07-16 05:40陈家长王菁裘丽萍孟顺龙范立民宋超
生态环境学报 2014年2期
陈家长 ,王菁,裘丽萍,孟顺龙,范立民,宋超
1. 中国水产科学研究院淡水渔业研究中心//中国水产科学研究院内陆渔业生态环境与资源重点开放实验室,江苏 无锡 214081;
2. 南京农业大学渔业学院,江苏 无锡 214081
水体富营养化和有害藻类爆发性生长已经成为制约我国水产养殖业健康发展的关键因素之一。随着我国水产养殖集约化程度的提高以及人工配合饲料的普遍使用,生产能力与养殖效益有了大幅度的提高,同时也带来了较为严重的负面影响:养殖区域水体的污染、病原生物大量繁殖、生态环境恶化、生物多样性遭到破坏(Habersack等,2014),甚至会影响到周边地区的环境(苏雅玲和邓一荣,2013;赵风斌,2012;Lei等,2014)。
浮游植物是水生态系统的初级生产者,其种群变动和群落结构直接影响水生态系统的结构和功能(孟顺龙,2013)。浮游植物间也存在着竞争现象,各种环境条件,如光照、温度、营养盐等,对竞争的结果具有重要影响(Flöder等,2006;Zhang等,2013)。
普通小球藻和鱼腥藻是池塘养殖水体中常见的藻类,其中普通小球藻能为好氧菌提供氧气,使之将复杂的有机物质分解为简单的无机物,达到对富含养分的废水进行资源化处理的目的(胡开辉等,2006);同时小球藻还是鱼类的优良饵料,且对水体中的氮磷有一定的去除作用(余云龙等,2012;刘淑坡和李飞,2012);鱼腥藻是富营养化水体和养殖水体中主要水华藻之一。尽管目前有关小球藻和鱼腥藻的研究较多(郑晓宇等,2012;Zhai等,2009;Zhu等,2013),但这些研究大多是探讨环境因素对单一藻种生长特性的影响,而有关普通小球藻和鱼腥藻种间竞争的报道并不多见。为此,本研究选用浅水湖泊(太湖等)以及养殖池塘中常见有益藻——普通小球藻和常见有害藻——鱼腥藻为试验藻种,探究不同pH下,普通小球藻和鱼腥藻的种间竞争,以期揭示水体中常见藻类的生长过程及其与pH的相互关系,为水产养殖过程控制(曹煜成等,2012)和精准培水技术的研究,以及控制养殖水体富营养化提供研究基础。
1 材料与方法
1.1 藻种与培养
实验用普通小球藻(Chlorella vulga)、鱼腥藻(Anabaenasp.strain PCC)购自中国科学院水生生物研究所。藻种扩大培养采用BG11培养基。实验中使用的玻璃仪器均经清水冲洗后,在质量浓度为1%的稀盐酸中浸泡30 min,再用无菌水冲洗,高温灭菌,烘干备用。试验期间的培养条件与扩大培养期间的培养时的条件一致,光照强度约为 2.5×103lx,光暗比12 h︰12 h,温度为25 ℃。光照期间,每隔2 h手工摇匀锥形瓶1次,暗期则静置。
1.2 试验设置
本研究设置4个pH梯度,分别为6,7,8和9。每个pH均设置3个试验组,分别为普通小球藻单独培养组(简称C组)、鱼腥藻单独培养组(简称A组)、普通小球藻和鱼腥藻共同培养组(简称CA组)。每组试验设置3个平行。
实验时,将达到接种浓度的普通小球藻、鱼腥藻在5000 r·min-1转速下离心8 min,去掉上清液,用BG11培养基稀释到实验所需浓度。各组普通小球藻、鱼腥藻的初始接种密度均设置为 5×105cell·mL-1。在容积为250 mL锥形瓶中加入对应pH的BG11培养液200 mL,然后置于智能光照培养箱(沃信仪器,无锡)内,在不同pH条件下进行一次性培养(中间不更换培养液),每24 h用0.1 mol/L的HCl和0.1 mol/L的NaOH对pH进行调节。
1.3 细胞计数
自实验开始后每24 h计数藻类数量。计数方法参照《水和废水监测分析方法(第四版)》。当所有藻类生物量均出现负增长时,试验结束,藻类出现负增长前 1 d的生物量即为该种藻类的最大现存量。
1.4 数据整理
1.4.1 比生长速率
根据藻液细胞密度计算比生长速率。比生长速率的计算方法为(Chu等,2007;孟顺龙等,2012):
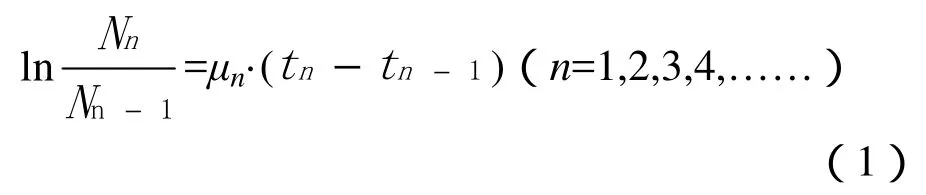
式中:μn为第n天的比生长速率;Nn为第n天的细胞密度(cells·L-1);Nn-1为第n-1天的细胞密度(cells·L-1);tn为对应于Nn的培养天数;tn-1为对应于Nn-1的培养天数。同时,将藻类从试验开始至生物量达最大现存量这一时间段内的比生长速率的平均值定义为藻类的平均比生长速率(μ),用于比较藻类生长速率的大小。
1.4.2 生长曲线拟合
以逻辑斯谛方程拟合藻类的增长过程。首先进行参数估计,每个处理组的最大生物量(Nmax)作为各自的K估计值。应用逻辑斯谛方程的对数形式(式 2),以最小二乘法进行回归分析,获得该方程的斜率和截距作为a和r的估计值。

式中:N为藻类生物量;K为最大生物量;r为内禀增长率;t为培养时间。
1.4.3 竞争抑制参数的计算
利用Lotka-Volterra竞争模型的差分形式(式3、式4)计算竞争抑制参数(孟顺龙等,2012)。

式中:Nc和Na分别为共同培养中的普通小球藻和鱼腥藻在时间tn时的数量(×104cells·mL-1);Ncn-1和Nan-1分别为共同培养中普通小球藻和鱼腥藻在时间tn-1时的数量(×104cells·mL-1);rc和ra分别为普通小球藻和鱼腥藻的内禀增长率(由单种培养经回归计算获得);Kc和Ka分别为普通小球藻和鱼腥藻的最大环境容量(由单种培养获得);α和β分别为共同培养中鱼腥藻对普通小球藻和普通小球藻对鱼腥藻的竞争抑制参数。
应用上述公式计算共同培养藻类的增长曲线在拐点以后至达到最大环境容量时间段内每一单位时间的所有竞争抑制参数,取其平均值作为该种藻类对另一种藻类的竞争抑制参数估计值。
参照Logistic方程,藻类的增长有一个从正加速到负加速的过程,即在增长曲线的拐点之后,藻类的增长过程减缓并逐渐趋于稳定在一个水平上。生物生长拐点的生物学意义是生物个体从自由的快速增长阶段转入相互抑制的生长阶段,也即密度制约的起始点(setting point)或抑制起始点(陈德辉等,1999)。因此,藻类增长曲线的拐点就是抑制起始点,即逻辑斯谛方程二阶导数等于零的时间tp值,这时N=K/2,tp=α/r(茅华等,2008),tp对α/r取整数(陈德辉等,1999)。
1.5 统计分析
采用单因素方差分析对数据进行统计处理,并用t检验方法对回归方程进行回归显著性检验;P<0.05时,差异显著。
2 结果与分析
2.1 不同pH下普通小球藻和鱼腥藻的生长情况
不同pH下,普通小球藻、鱼腥藻的生长情况如图1所示。方差分析表明,无论是在单种培养还是在共同培养体系中,pH对两种藻类的最大生物量均产生显著影响。单种培养体系中,普通小球藻和鱼腥藻的最大生物量随着pH的增加而增加,pH 9时的最大生物量显著高于其他组(p<0.05)。在相同培养时间下,普通小球藻的生物量基本表现为pH9>pH8>pH7>pH6;鱼腥藻的生物量均表现为pH9>pH8>pH7>pH6。共同培养体系中,普通小球藻的最大生物量表现为 pH9>pH8>pH6>pH7,且 4种pH间最大生物量差异显著(p<0.05),在相同培养时间下,普通小球藻的生物量在1~8 d中基本表现的为:pH9>pH7>pH8>pH6。共同培养体系中,鱼腥藻的最大生物量随pH的升高而增加,且4种pH间最大生物量差异显著(p<0.05),同时,在相同培养时间下的鱼腥藻生物量也基本表现为pH9>pH8>pH7>pH6,这与单种培养体系相同。
由表1可见,pH能够对两种藻的平均比生长速率产生影响。无论在单种培养还是共同培养下,鱼腥藻的平均比生长速率均表现为:pH9>pH8>pH7>pH6;而普通小球藻的平均比生长速率在单种培养条件下表现为:pH9>pH7>pH8>pH6,在共同培养条件下表现为:pH9> pH8>pH6>pH7。单种培养体系中,普通小球藻和鱼腥藻在不同pH下的生长曲线基本符合S型生长曲线(图1),说明不同pH下,单种培养藻类的生长曲线均可用Logistic模型拟合,并可以根据Logistic方程计算拐点出现时间(表2)。同时,为计算拐点出现时间,共同培养体系中的普通小球藻、鱼腥藻的生长也用Logistic方程进行了拟合,并由此得到各生长曲线的拐点出现时间(表2)。

图1 不同pH普通小球藻和鱼腥藻单种培养组及共同培养组的生长曲线Fig. 1 Growth curves of Chlorella vulga and Anabaenasp.strain PCC in pure culture and mixed culture system s at different pH

表1 不同pH下普通小球藻和鱼腥藻的平均比生长速率Table 1 Mean μ of Chlorella vulga and Anabaenasp.strain PCC at different pH (μ)
2.2 普通小球藻和鱼腥藻两两间竞争抑制参数
以单种培养体系中拟合得到的K、r值和共同培养体系中普通小球藻和鱼腥藻的细胞数带入式(3)、(4),计算共同培养体系中普通小球藻对鱼腥藻以及鱼腥藻对普通小球藻竞争抑制参数(表3)。
如表3所示,在4个pH条件下普通小球藻对鱼腥藻的竞争抑制参数(β)均大于鱼腥藻对普通小球藻的竞争抑制参数(α)。pH7.0时普通小球藻对鱼腥藻的竞争抑制参数(β)最大;鱼腥藻对普通小球藻的竞争抑制参数(α)则是pH6.0时最大。同时,由表2可见,单种培养体系中,4个 pH条件下,鱼腥藻出现拐点的时间都比普通小球藻晚;在共同培养体系中,在pH6.0、7.0、9.0时,普通小球藻的拐点出现时间均早于鱼腥藻,而在pH8.0时,普通小球藻和鱼腥藻出现拐点的时间基本一致。
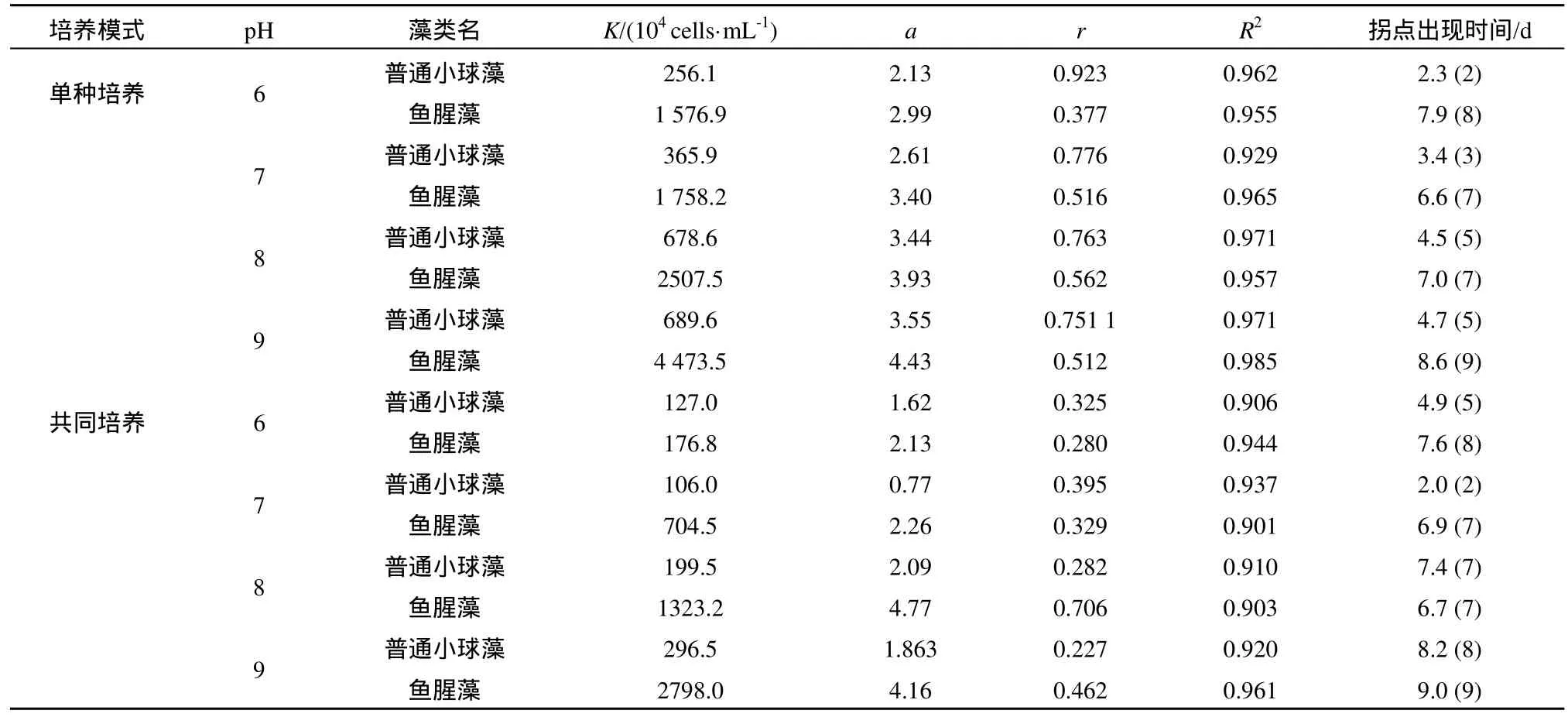
表2 不同pH下普通小球藻和鱼腥藻的逻辑斯谛模型拟合参数及拐点出现的时间Table 2 Parameters of Chlorella vulga and Anabaenasp.strain PCC regression coefficients of Logistic model and the inflection points at different pH
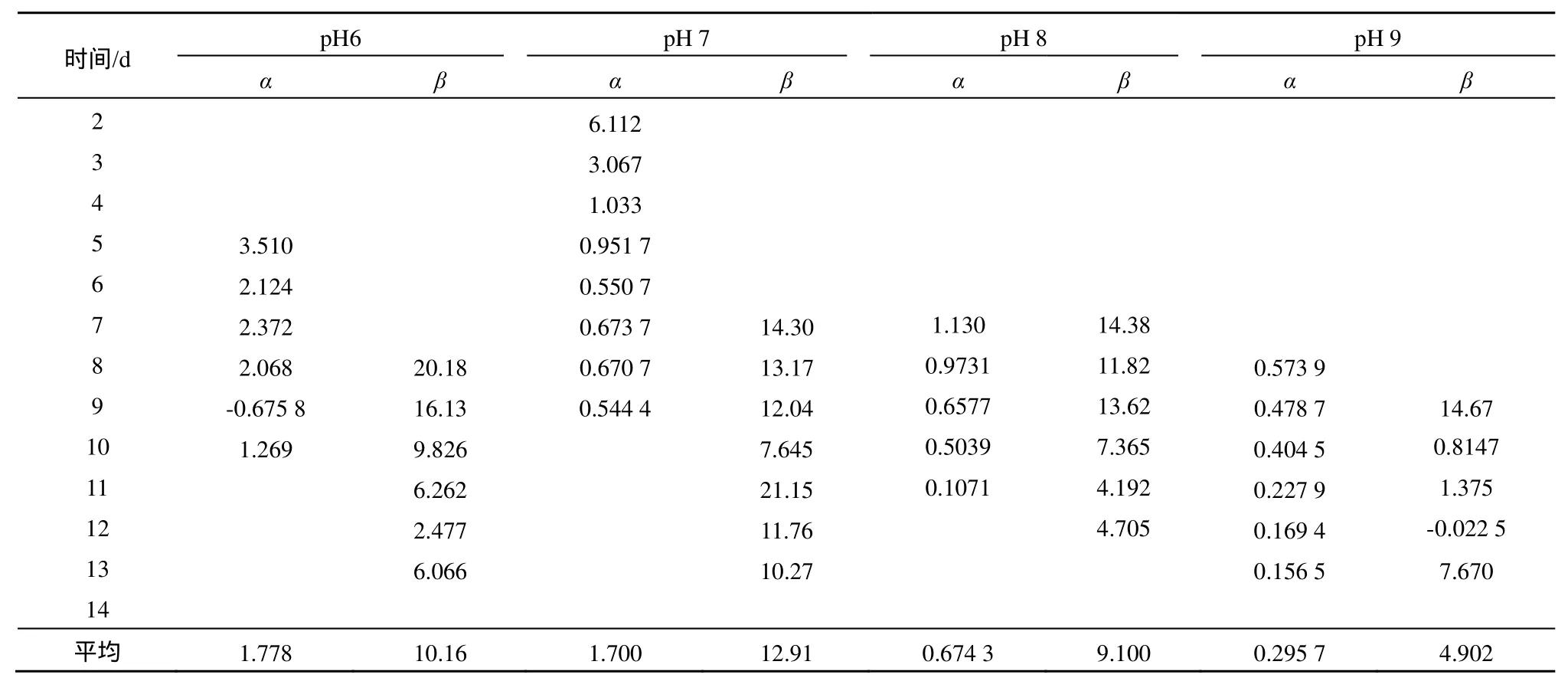
表3 不同pH下普通小球藻和鱼腥藻的竞争抑制参数Table 3 Inhibition parameters of Chlorella vulga and Anabaenasp.strain PCC after inflection point at different pH
3 讨论
3.1 pH对单种培养体系中普通小球藻和鱼腥藻生长的影响
水体pH主要从两方面对微藻产生影响,一方面改变环境酸碱度,酸性太强(H+浓度高)或碱性太强(OH-浓度高)都会对藻细胞产生伤害,只有在适宜的酸碱度范围内,藻细胞才能正常生长繁殖;另一方面影响碳酸盐平衡系统及不同形态无机碳分配关系来影响藻类的生长(王菁等,2013)。目前实验条件下还没有办法区分藻类对pH的反应究竟是由于受到酸碱伤害还是由于改变了碳酸盐平衡系统。本实验中,鱼腥藻和普通小球藻在pH6时生长均受到抑制,应该是受到酸害的作用。大部分藻类对pH的耐受范围较窄,贺春花等(2011)实验结果显示:颤藻在pH7.3~8.6的偏碱性条件下生长良好;茅华等(2007)实验结果显示旋链角毛藻的最适pH为8.3;许海等(2009)研究了水华鱼腥藻在pH6、7、8、9、10对3种淡水蓝藻和3种淡水绿藻生长的生长情况,结果表明,水华鱼腥藻最适生长pH在8.0~9.0,绿球藻为7.0~8.0;欧阳峥嵘等(2010)研究表明,小球藻适宜在中性偏碱的条件下生长,在pH6.0中,净光合放氧速率下降为零。从本文的研究结果看,普通小球藻和鱼腥藻的最大生长量随着pH的增大而增加,本试验中其最适生长的pH均为9.0。
3.2 pH对普通小球藻和鱼腥藻生长竞争的影响
薛凌展(2010)的研究表明铜绿微囊藻pH在8~11范围内生长良好,其最佳生长pH值为9,而普通小球藻则pH在7~9之间生长较好,其最佳pH值为 8。共同培养的试验结果显示碱性环境中铜绿微囊藻的竞争优势强于普通小球藻,pH为8~10时β值大于α值,而pH为7时普通小球藻占优势,α值大于β值。从本研究结果看,试验pH下,共同培养中普通小球藻和鱼腥藻的生长量都远小于单种培养中的生长量(P<0.05);pH7.0时普通小球藻对鱼腥藻的竞争抑制参数最大,分别为pH6.0、7.0、9.0下的1.27倍、1.42倍、4.33倍;鱼腥藻对普通小球藻的竞争抑制参数(α)则是pH 6.0时最大,分别是 pH 7.0、pH 8.0、pH 9.0时的 1.05、2.64、6.20倍。这种现象说明,pH对藻类的竞争抑制参数能够产生较大影响,且鱼腥藻对普通小球藻的竞争能力在pH6.0时环境下最强,而普通小球藻对鱼腥藻的竞争能力则在pH7.0时最强。
从共同培养体系中普通小球藻和鱼腥藻的拐点出现时间看,在pH 6.0、7.0、9.0时,普通小球藻的拐点出现时间均早于鱼腥藻,而在pH 8.0时,普通小球藻和鱼腥藻的拐点都出现第7天。生物生长拐点的生物学意义是生物个体从自由的快速增长阶段转入互相抑制的生长阶段,也即抑制的起始点(陈德辉,等,1999)。上述现象说明,在pH 6.0、7.0、9.0时,普通小球藻的生长首先被抑制;而在pH 8.0时,普通小球藻和鱼腥藻几乎同时对对方产生抑制。
藻类种间抑制主要是通过竞争光照、营养盐等资源而产生的抑制以及分泌克生物质而产生的抑制(郑朔方等, 2005)。从本试验中鱼腥藻和普通小球藻的竞争抑制参数研究结果看(表3),在4个pH条件下,虽然鱼腥藻对普通小球藻的抑制作用先于普通小球藻对鱼腥藻的抑制作用,但鱼腥藻对普通小球藻的竞争抑制参数(α)均小于普通小球藻对鱼腥藻的竞争抑制参数(β);说明在试验 pH下,鱼腥藻对普通小球藻的抑制能力均小于普通小球藻对鱼腥藻的抑制能力,说明普通小球藻在竞争中占优势。这种现象可能是因为普通小球藻分泌出的某种抑制物质的作用强于鱼腥藻分泌出的抑制物质。
4 结论
在试验所设pH条件下,无论在单种培养体系还是共同培养体系中,鱼腥藻的最大现存量都随着pH的升高而增加,说明碱性条件对鱼腥藻生长有利,同时也表明鱼腥藻是一种耐碱性藻类。单种培养条件下,普通小球藻的最大现存量随pH升高而增加;共同培养条件下,最大现存量随pH的变化表现为pH9>pH8>pH6>pH7。
pH对藻类的竞争抑制参数能够产生显著影响,pH 6.0时,鱼腥藻对普通小球藻的竞争抑制参数最大;而普通小球藻对鱼腥藻的竞争抑制参数则是在pH 7.0时最大。在4个pH条件下,鱼腥藻对普通小球藻的竞争抑制参数(α)均小于普通小球藻对鱼腥藻的竞争抑制参数(β);说明抑制作用与环境中藻种的相对藻量有关,在试验pH下,鱼腥藻对普通小球藻的抑制能力均小于普通小球藻对鱼腥藻的抑制能力,普通小球藻在竞争中处于优势。
CHU Z S,JIN X C,IWAMI N,et al. 2007.The effect of temperature on growth characteristics and competitions ofMicrocystis aeruginosaandOscillatoria mougeotiiin a shallow, eutrophic lake simulator system[J].Hydrobiologia, 581(1): 217-223.
FLÖDER S, COMBÜCHEN A, PASTERNAK A, et al. 2006.Competition between pelagic and benthic microalgae for phosphorus and light[J].Aquatic Sciences, 68(4):425-433.
HABERSACK H, HASPEL D, MUHAR S, et al. 2014. Preface: Impact of human activities on biodiversity of large rivers[J]. Hydrobiologia,729(1):1-2.
LEI L, PENG L, HUANG X, et al. 2014.Occurrence and dominance ofCylindrospermopsis raciborskiiand dissolved cylindrospermopsin in urban reservoirs used for drinking water supply, South China[J].Environmental Monitoring and Assessment, 186(5):3079-3090.
MENGYUAN ZHU, GUANGWEI ZHU, LINLIN ZHAO, et al.2013.Influence of algal bloom degradation on nutrient release at the sediment–water interface in Lake Taihu, China[J].Environmental Science and Pollution Research, 20(3):1803-1811.
SHUIJING ZHAI, LONGYUAN YANG, WEIPING HU. 2009.Observations of atmospheric nitrogen and phosphorus deposition during the period of algal bloom formation in northern lake Taihu,China[J].Environmental management, 44(3):542-551.
曹煜成, 王少沛, 李卓佳, 等. 2012.微绿球藻、隐藻和颤藻竞争氮磷营养的通径分析[J]. 渔业科学进展, (04):99-106
陈德辉,刘永定,袁俊峰,等. 1999.微囊藻和栅藻共培养实验及其竞争参数的计算[J].生态学报, 19(6):908-913.
贺春花,黄翔鹄,李长玲,等. 2011.温度、光照度、盐度和pH对颤藻生长的限制条件研究[J].渔业现代化, 38(6):20-25.
胡开辉,朱行,汪世华,等. 2006.小球藻对水体氮磷的去除效率[J].福建农林大学学报:自然科学版, 35(6):648-651.
刘淑坡,李飞. 2012.固定化核蛋白小球藻对人工废水中不同形态氮和磷的去除[J].山东理工大学学报:自然科学版, 26(4):43-47.
茅华,许海,刘兆普,等. 2008.不同起始细胞数量对旋链角毛藻和中肋骨条藻种群竞争的影响[J]. 海洋环境科学, 27(5):458-461.
茅华,许海,刘兆普. 2007.温度、光照、盐度及pH对旋链角毛藻生长的影响[J].生态科学, 26(5):432-436.
孟顺龙, 裘丽萍, 胡庚东,等. 2012. 氮磷比对两种蓝藻生长及竞争的影响[J]. 农业环境科学学报, (7):1438-1444.
孟顺龙, 瞿建宏, 裘丽萍,等. 2013. 富营养化水体降磷对浮游植物群落结构特征的影响[J]. 生态环境学报, (09):1578-1582.
欧阳峥嵘, 温小斌, 耿亚红 , 等. 2010.光照强度、温度、pH、盐度对小球藻(Chlorella)光合作用的影响[J]. 武汉植物学研究, (01):49-55.
苏雅玲, 邓一荣. 2013.富营养化湖泊中微囊藻毒素及其控制去除技术[J].环境科学与技术, (06):62-66.
王菁, 陈家长, 孟顺龙. 2013.环境因素对藻类生长竞争的影响[J]. 中国农学通报, 29(17):52-56.
许海,刘兆普,袁兰,等. 2009.pH对几种淡水藻类生长的影响[J].环境科学与技术, 32(1):27-30.
薛凌展,黄种特,林泽,等. 2010.铜绿微囊藻和普通小球藻在不同pH下生长特性及竞争参数计算[J].福建农业学报, 25(2):142-148.
余云龙,邹华,张强,等. 2012.黑暗条件下普通小球藻处理废水的研究[J].食品与生物技术学报, 31(9): 938-943.
赵风斌. 2012.富营养化水体中常见沉水植物恢复重建影响因子研究[D].上海:上海海洋大学:48.
郑朔方,杨苏文,金相灿. 2005.铜绿微囊藻生长的营养动力学[J].环境科学,(2):152-156.
郑晓宇,金妍,任翔宇,等. 2012.不同氮磷浓度对铜绿微囊藻生长特性的影响[J].华东师范大学学报:自然科学版, (1):11-18.
猜你喜欢
西南农业学报(2022年8期)2022-10-28
当代水产(2021年8期)2021-11-04
军事文摘(2020年20期)2020-11-16
陶瓷学报(2019年5期)2019-01-12
今日农业(2019年10期)2019-01-04
江苏农业科学(2017年24期)2018-01-12
环境科技(2017年1期)2017-09-05
农业与技术(2017年6期)2017-04-21
环境科技(2016年2期)2016-11-08
化工进展(2015年6期)2015-11-13
