
泥蚶血红蛋白的制备及其抗菌活性研究
2014-06-01 12:30王素芳包永波施淼江郑丹妮杨婷婷林志华
海洋学报 2014年12期
王素芳,包永波,施淼江,郑丹妮,杨婷婷,林志华
(1.宁波大学海洋学院,浙江宁波 315211;2.浙江万里学院浙江省水产种质资源高效利用技术研究重点实验室,浙江宁波 315100)
泥蚶血红蛋白的制备及其抗菌活性研究
王素芳1,2,包永波2*,施淼江2,郑丹妮2,杨婷婷1,2,林志华2
(1.宁波大学海洋学院,浙江宁波 315211;2.浙江万里学院浙江省水产种质资源高效利用技术研究重点实验室,浙江宁波 315100)
利用凝胶层析技术从泥蚶血细胞裂解物中分离纯化得到两种血红蛋白,回收率为74.3%,并通过Tricine-SDS-PAGE和质谱技术鉴定为Tg-HbⅠ(同源二聚体)和Tg-HbⅡ(异源四聚体)。分别通过琼脂扩散法和分光光度法测其抗菌活性和过氧化物酶活性,结果发现,两种Tg-Hb对大肠杆菌和恶臭假单胞杆菌均有抗菌活性,Tg-HbⅡ对于恶臭假单胞菌和大肠杆菌的MIC值分别为0.063 mg/mL和0.13 mg/m L,Tg-HbⅠ对两种菌的MIC均为0.048 mg/m L;两种Tg-Hb均具有过氧化物酶活性,能催化愈创木酚、邻苯二酚、对苯二酚、苯酚、左旋多巴等多种酚类物质。在Tg-Hb中添加GSH后,两种Tg-Hb均不再对恶臭假单胞杆菌具有抗菌活性。可见,两种Tg-Hb对恶臭假单胞杆菌等的抗菌活性均是通过其过氧化物酶活性发挥作用的。
泥蚶;血红蛋白;抗菌活性;过氧化物酶活性
1 引言
血红蛋白(hemoglobin,Hb)是生物界分布最广的一类呼吸蛋白,主要在脊椎动物和部分无脊椎动物中存在[1]。近十多年的研究发现,Hb除了具有运输和储存氧气的功能外,还具有调节p H值、控制体内一氧化氮水平、参与免疫反应、运输硫化物、类卵黄蛋白原以及调节浮力等多种生物学功能[2—3]。尽管Hb是研究最多的一类呼吸蛋白,但以往的研究主要集中在其结构与携氧功能方面。直到近几年,受血蓝蛋白抗菌活性研究的启发,一些科学家才开始关注Hb的抗菌活性,Neajar-Arroume等[4—6]几位科学家先后报道了脊椎动物Hb及其多肽片段的抗菌活性,关于无脊椎动物Hb的抗菌活性的研究就更少。
泥蚶Tegillarca granosa隶属于软体动物门瓣鳃纲,是一种具有重要经济价值的海洋无脊椎动物,同时又是一类较特殊的含有Hb的双壳贝类。因其没有T细胞,缺乏真正意义上的抗体、没有类似脊椎动物的特异性免疫系统,因而只能依靠血细胞及体液因子的先天免疫反应来抵御外界环境中病原微生物的入侵。因此,进行泥蚶血红蛋白(Tg-Hb)的抗菌活性研究不仅有利于提高其自身的抗病能力,对于无脊椎动物免疫学的发展也具有重要的理论与实践意义。
2 材料与方法
2.1 材料
2.1.1 实验材料
泥蚶购自宁波市鄞州区东裕菜市场。金黄色葡萄球菌ATCC 29213、大肠杆菌ATCC35218、恶臭假单胞菌CGMCC1.0593、副溶血弧菌和哈维氏弧菌为浙江万里学院微生物与环境工程重点实验室提供。还原型谷胱甘肽(GSH)为Sigma公司产品,血红蛋白浓度测定试剂盒为北京瑞尔达生物科技有限公司,其他试剂均为国产分析纯。
2.1.2 主要仪器
AKTA蛋白层析系统(Explore100,GE公司),均质机(SH-2A,IKA公司),紫外分光光度计(3300 pro,美国Amershaw),二维纳升级超高效液相质谱串联仪(Nano Acquity UPLC/Synapt G2 MS,Waters)凝胶排阻色谱柱Sephacryl S-100 HR(HiPrep26/60,GE公司),CM-Sepharose Fast Flow(GE公司),立式压力蒸汽灭菌器(YXQ-LS-100SⅡ,上海迅博实业有限公司),恒温培养箱(DHP-9162,上海慧泰仪器制造有限公司)。
2.2 泥蚶血红蛋白的分离纯化
将新鲜的泥蚶洗净后,在洁净的海水中吐沙4 h。撬开贝壳后,采集血液,置于含有柠檬酸抗凝剂的离心管中,2 000 r/min离心10 min,得血细胞。用0.9%NaCl溶液多次洗涤至上层溶液无色透明,用PBS缓冲液悬浮,匀浆破碎,以12 000 r/min的转速4℃离心30 min,上清为泥蚶血红蛋白粗品。
将上述的血红蛋白粗品上Sephacryl S-100 HR凝胶层析柱,用PBS缓冲液以0.3 m L/min的流速洗脱,以2 mL/管收集洗脱液,用280 nm和415 nm两种波长检测洗脱液。
2.3 泥蚶血红蛋白浓度测定
采用血红蛋白测定试剂盒测定。测浓度分别为0、25、50、75、100 g Hb/L的氰化高铁血红蛋白标准液540 nm波长处的吸光度(A),以A为横坐标Hb浓度为纵坐标做标准曲线,求斜率K值,

取5μL样品液加入含1 mL B1溶液的比色皿中,混匀用B1做空白,在595 nm下测吸光度(C),

2.4 Tricine-SDS-PAGE
Tricine-SDS-PAGE参照文献[7]进行,其中浓缩胶聚丙烯酰胺浓度为4%,分离胶浓度为16%。电泳结束后,以考马斯亮蓝染色。
2.5 琼脂扩散法测抑菌活性
将对数生长期的细菌,用无菌生理盐水配制成105cfu/m L的菌悬液,取200μL均匀涂布在制好的培养基上,放置牛津杯后依次加入200μL生理盐水、HbⅠ(1.5 mg/mL)和HbⅡ(2 mg/m L)。金黄色葡萄球菌、大肠杆菌和恶臭假单孢杆菌置37℃生化箱培养24 h,哈维式弧菌和副溶血弧菌则置28℃生化箱培养24 h,取出观察拍照。
2.6 最低抑菌浓度(MIC)测定
采用微量肉汤稀释法。将倍比稀释后不同浓度的抗菌药物溶液分别加到灭菌的96孔聚苯乙烯板中,第1至第11孔加Tg-Hb,每孔100μL,第12孔不加药作为生长对照。将浓度相当于0.5麦氏比浊标准的菌悬液,经MH肉汤1∶1 000稀释后,向每孔中加20μL,密封后置35℃普通空气孵箱中,孵育16~20 h判断结果。
2.7 过氧化物酶活性测定
取1 mL 50 mmol/L磷酸盐缓冲液(4 mmol/L愈创木酚2 mmol/L H2O2)于一次性比色皿中,加入血红蛋白,在25℃470 nm波长处,用动力学模式,测其30 s内的A470的变化。
2.8 米氏常数的测定
分别用邻苯二酚、对苯二酚、苯酚、3-羟基-L-酪氨酸(L-DOPA)L-酪氨酸和对甲酚等代替愈创木酚反应,在25℃测其30 s内的A470的变化。以1/ΔA为纵坐标,以1/[S]横坐标做双倒数图,求各酚类物质的米氏常数。
2.9 泥蚶血红蛋白的质谱鉴定
将收集的峰2、峰3样品,胰蛋白酶消化后用二维纳升级超高效液相质谱串联仪分析。将得到的数据用Mascot检索引擎在Swiss Prot数据库中进行检索鉴定。
3 结果与分析
3.1 泥蚶血红蛋白的分离纯化
血红蛋白粗品经凝胶层析后,洗脱图谱见图1。根据280 nm和415 nm(Tg-Hb在415 nm处有特征性的高吸光度)两种波长下的检测图谱,可初步判断峰1主要为杂蛋白,峰2、峰3为Tg-Hb的洗脱峰。已有文献报道[8],贝类血红蛋白含有同源二聚体(HbⅠ)和异源四聚体(HbⅡ)两种形式,其同源二聚体含有两个相同或相似的亚基,每个亚基的分子质量约为15~16 kD;而异源四聚体则具有α2β2结构,分子质量约为60~70 k D。可见,峰2为泥蚶血红蛋白Ⅱ(Tg-HbⅡ),峰3为泥蚶血红蛋白Ⅰ(Tg-HbⅠ)。Tricine-SDS-PAGE结果显示,峰1成分非常复杂,含多种大分子量的蛋白条带,峰2样品有两个蛋白条带,峰3样品为单一条带。Tricine-SDSPAGE中,峰2、峰3样品均未出现杂蛋白条带,说明均达到了较高的纯度(图2),由于分离过程简单,Tg-Hb的回收率比较高,达74.3%(表1)。
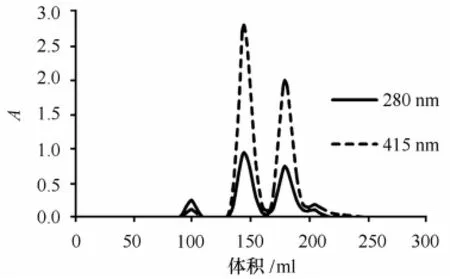
图1 泥蚶血红蛋白的凝胶层析图Fig.1 Gel chromatography of Tg-Hb
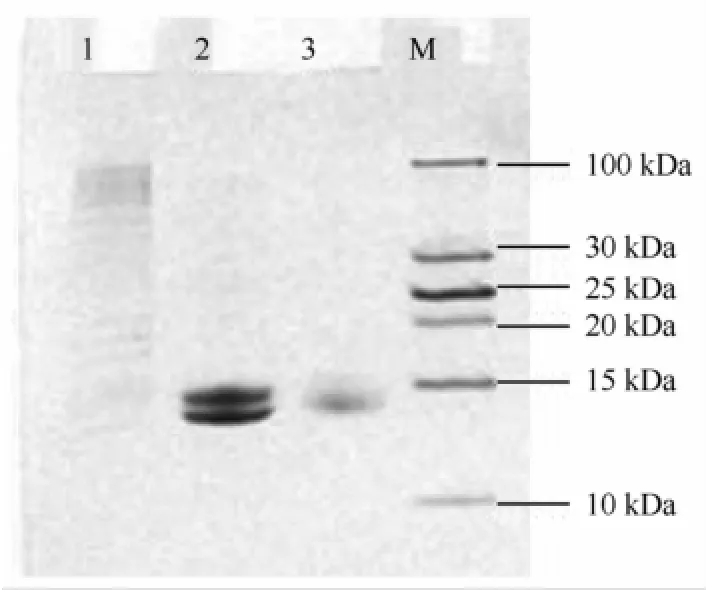
图2 洗脱峰的Tricine-SDS-PAGE 1为峰1;2为峰2;3为峰3;M为低分子量标准蛋白Fig.2 Tricine-SDS-PAGE analysis of elution peak Lanes 1 to 3 indicate samples of peak 1,peak 2 and peak 3,M indicate marker

表1 泥蚶血红蛋白的回收率Tab.1 Recovery of Tg-Hb from the blood of Tegillarca granosa
3.2 泥蚶血红蛋白的质谱鉴定
为了进一步确定峰2、峰3组分是泥蚶血红蛋白,对其进行了质谱鉴定(图3、4)。结果显示,峰2的一条链与不等壳毛蚶Scapharca inaequivalvisHbⅡ-A最相似;一条链与梯形毛蚶Anadara trapezia血红蛋白的β链最相似;峰3与不等壳毛蚶的HbⅠ(同源二聚体)最相似。最终,峰2鉴定为Tg-HbⅡ(异源四聚体),峰3鉴定为Tg-HbⅠ(同源二聚体),与3.1中的推断一致。

图3 凝胶层析峰2的质谱图Fig.3 MSspectrum for peak 2 of gel chromatography

图4 凝胶层析峰3的质谱图Fig.4 MSspectrum for peak 3 of gel chromatography
3.3 泥蚶血红蛋白的抗菌活性
琼脂扩散法检测了Tg-Hb对金黄色葡萄球菌、大肠杆菌、恶臭假单胞菌、副溶血弧菌和哈维氏弧菌等菌的抗菌活性。结果表明,两种Tg-Hb对大肠杆菌、恶臭假单胞菌这两种革兰氏阴性菌均有抗菌效果,有明显的抑菌圈,而未检测到Tg-Hb对其他几种菌的抑菌圈(见表2),可见两种Tg-Hb的抗菌活性都有选择性。Tg-HbⅡ对大肠杆菌和恶臭假单胞菌的MIC分别为0.13 mg/mL和0.063 mg/mL(见表3),Tg-HbⅠ对两种菌的MIC均为0.048 mg/mL(见表4)。

表2 泥蚶血红蛋白对6种菌的抗菌活性Tab.2 Antimicrobial activities of Tg-Hb against bacteria
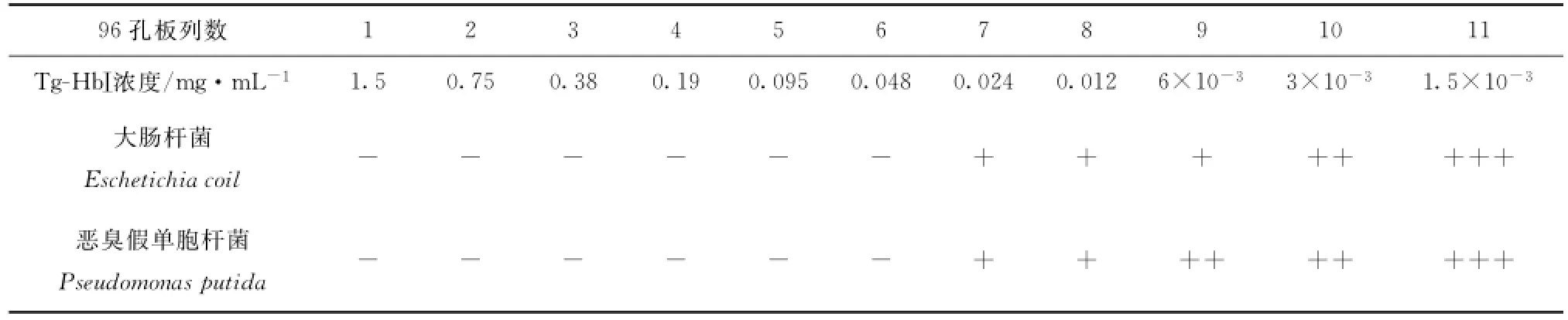
表3 不同浓度Tg-HbⅠ的抗菌效果Tab.3 Antibacterial activities of Tg-HbⅠat different concentration

表4 不同浓度Tg-HbⅡ的抗菌效果Tab.4 Antibacterial activities of Tg-HbⅡat different concentration
3.4 泥蚶血红蛋白的抗菌机制
酶活性研究结果表明,两种Tg-Hb均具有过氧化物酶活性(见图5),能催化多种酚类物质(见表5)。一般认为,过氧化物酶可催化酚类物质生成醌及其后续一系列的产物,在此过程中产生活性氧(ROS),从而起到抗菌作用。本研究证实了这一点,在添加40 mmol/ L还原型谷胱甘肽(GSH,活性氧清除剂)的实验组,Tg-Hb的抗菌能力丧失(见图6)。因此判断,Tg-Hb的抗菌活性与其过氧化物酶活性相关,通过过氧化物酶活性产生的活性氧发挥抗菌作用,当产生的ROS被GSH清除后,便不能发挥抗菌作用。但为什么Tg-Hb的抗菌作用有选择性,这需要进一步深入研究。

图5 Tg-Hb的过氧化物酶活性Fig.5 Peroxidase activity of Tg-Hb
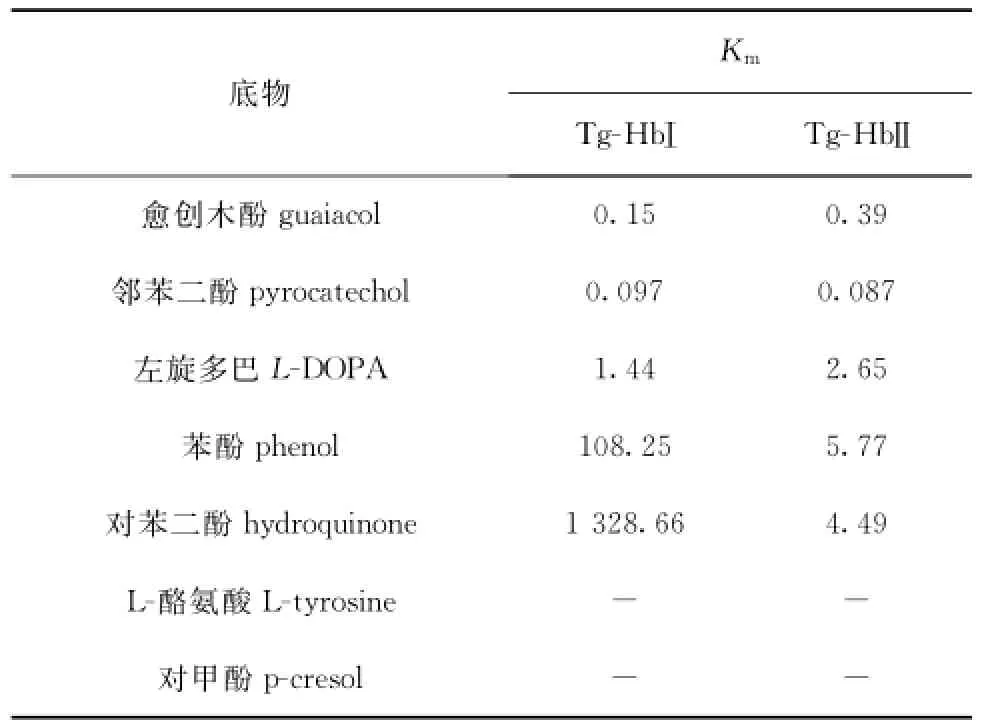
表5 Tg-Hb的底物特异性Tab.5 Substrate specificity of Tg-Hb

图6 GSH对泥蚶血红蛋白抗菌活性的影响Fig.6 Effect of GSH on the antibacterial activity of Tg-Hb
4 讨论
4.1 泥蚶血红蛋白的分离纯化
开展Tg-Hb免疫学研究的前提是得到纯化的Tg-Hb,但直到目前为止,仍未见到有关Tg-Hb高效分离纯化的研究报道。本实验室在既往的研究中,已经克隆到Tg-Hb 3个亚基的基因Tg-HbⅠ、Tg-HbⅡA和Tg-HbⅡB[9],并用软件预测其分子量分别为16.04 k Da、16.23 kDa和17.23 k Da,等电点分别为9.144、8.836和5.745,因此进一步推断Tg-HbⅠ、Tg-HbⅡ的分子量分别约为32 kDa和67 k Da,等电点可能分别在9和7.3附近。根据两种Tg-Hb等电点和分子量的差异,选择了阳离子交换层析和凝胶层析两种层析技术。结果发现,阳离子交换层析并不适合Tg-Hb的纯化。在上样缓冲液p H6.0条件下,阳离子交换层析仅能吸附Tg-HbⅠ,不吸附Tg-HbⅡ,因此不能实现两种Tg-Hb的同时纯化。此外,在研究过程中发现Tg-Hb在偏离p H7.2的环境下易解聚,在p H6.0的条件下过夜就有一部分Tg-HbⅠ解聚成单体,因此采用进一步降低上样缓冲液p H的方法更不可行。凝胶层析技术能实现两种高纯度Tg-Hb的同时分离,回收率高达74.3%(见图1~3),而且凝胶层析采用的是p H7.2的PBS缓冲液,两种Tg-Hb在p H7.2的PBS缓冲液中稳定性都较好。此外,本研究采用280 nm和415 nm双波长检测洗脱峰,实现了Tg-Hb峰方便快速判断,为其他物种血红蛋白的分离纯化提供了参考。
4.2 泥蚶血红蛋白的抗菌活性
近些年,呼吸蛋白的抗菌作用已成为国内外研究的热点,已有多位科学家证实了部分呼吸蛋白的抗菌作用。Fang等[5—6]发现人血红蛋白对金黄色葡萄球菌、铜绿假单胞菌等细菌的抗菌作用;荆昭[3]发现毛蚶Scapharca kagoshimensis血红蛋白对枯草杆菌和四联球菌等菌具有抗菌作用;Jiang等[6]的研究表明鲎血蓝蛋白就具有抗金黄色葡萄球菌等菌的作用;Deloffre等[10]发现沙蚕Nereis diversicolor血紫蛋白也具有抗细菌活性。本研究则证实Tg-Hb对大肠杆菌和恶臭芽孢杆菌等部分革兰氏阴性菌的抗菌作用。
呼吸蛋白的酶活性方面,关于血蓝蛋白的酚氧化酶活性的报道较多。多位科学家[11—15]已证明血蓝蛋白能被SDS、胰蛋白酶和入侵微生物等因子激活,具有酚氧化物酶活性,产生ROS,发挥抗菌作用。血蓝蛋白为什么具有酚氧化酶活性,一些科学家从结构方面找到了答案。血蓝蛋白与酚氧化物酶同属于Ⅲ型铜蛋白,都具有两个铜离子,在活性位点的基因序列与蛋白质结构均具有高度的相似性[16]。Hb虽然是研究最广泛和深入的呼吸蛋白,但长期以来,关于Hb结构与功能关系的研究都集中在它如何与O2结合、分离和运输O2等方面,Hb的类酶催化活性方面研究不多。2002年,Kawano等[17]发现人血红蛋白的过氧化物酶活性,但其后大家的目光均集中在人血红蛋白的过氧化物酶活性对机体的伤害[18]及作为过氧化物酶替代品的可行性研究方面[19—20],并没有意识到Hb具有抗菌活性,更没有意识到Hb可利用其过氧化物酶活性发挥抗菌作用。受血蓝蛋白抗菌及酚氧化酶研究的启发,一些科学家也开始关注Hb的抗菌作用及其与过氧化物酶活性的关系。Fang[5]的研究表明,当有细菌侵入机体时人血红细胞内的血红蛋白会释放出ROS,迅速杀死致病微生物。Jiang等[6]研究得出人血红蛋白在入侵病菌毒力因子的刺激下获得了一种过氧化物酶的活性,催化产生ROS,进而产生有毒衍生物,起到杀菌作用。
本研究进一步证实了Tg-Hb对大肠杆菌和恶臭芽孢杆菌等部分革兰氏阴性菌也有抗菌作用,也检测到了其过氧化物酶活性,并且证明Tg-Hb的抗菌活性与其过氧化物酶活性有关(见图5、6)。此外,Tg-Hb的抗菌作用具有选择性,如对金黄色葡萄球菌、副溶血弧菌等菌并没有抗菌作用,这又说明其本身的过氧化物酶活性可能偏低,不足以抗菌。分析其抗菌机理,很可能是敏感菌分泌物或组分进一步激活了它的过氧化物酶活性,产生了大量的ROS,从而发挥抗菌作用。这些发现,为进一步阐明Hb的抗菌机制提供了新的思路。
[1] Riggs A F.Aspects of the origin and evolution of non-vertebrate hemoglobins[J].American Zoologist,1991,31(3):353-545.
[2] Hardison R C.A brief history of hemoglobins:plant,animal,protest,and bacteria[J].Proc Natl Acad Sci U S A,1996,93(12):5675-5679.
[3] 荆昭.毛蚶(Scapharca kagoshimensis)血红蛋白的类酚氧化酶性质及其抗菌活性研究[D].青岛:中国海洋大学,2011.
[4] Nedjar-Arroume N,Dubois-Delval V,Adje E Y,et al.Bovine hemoglobin:an attractive source of antibacterial peptides[J].Peptides,2008,29(6):969-977.
[5] Fang F C.Antimicrobial reactive oxygen and nitrogen species:concepts and controversies[J].Nat Rev Microbi,2004,2(10):820-832.
[6] Jiang N X,Tan N S,Ho B,et al.Respiratory protein-generated reactive oxygen species as an antimicrobial strategy[J].Nat Immunol,2007,8(10):1114-1122.
[7] Schägger H.Tricine-SDS-PAGE[J].Nat Protoc,2006,1(1):16-22.
[8] Chiancone E,Vecchini P,Verzili D,et al.Dimeric and tetrameric hemoglobins from the molluscScapharca inaequivalvis:structural and functional properties[J].J Mol Biol,1981,152(3):577-592.
[9] Bao Y B,Wang Q,Lin Z H.Hemoglobin of the bloody clamTegillarca granosa(Tg-HbⅠ)isinvolved in the immune responseagainst bacterial infection[J].Fish&Shellfish Immunology,2011,31(4):517-523.
[10] Deloffre L,Salzet B,Vieau D,et al.Antibacterial properties of hemerythrin of the sand wormNereis diversicolor[J].Neuro Endocrinol Lett,2003,24(1/2):39-45.
[11] Paul R J,Pirow R.The physiological significance of respiratory proteins in invertebrates[J].Zoology,1998,100:319-327.
[12] Fan T J,Zhang Y,Yang L L,et al.Identification and characterization of a hemocyanin-derived phenoloxidase from the crabCharybdis japonica[J].Comp Biochem Physiol,2009,152(2):144-149.
[13] Decker H,Tuczek E.Tyrosinase/catecholoxidase activity of hemocyanins:structural basis and molecular mechanism[J].Trends in Biochemical Sciences,2000,25(8):392-397.
[14] Siddiqui N I,Préaux G,Gielens C.Intrinsic and induced o-diphenoloxidase activity ofβ-hemocyanin ofHelix pomatia[J].Micron,2004,35(1/2):91-92.
[15] Zhang X B,Huang C H,Qin Q W.Antiviral properties of hemocyanin isolated from shrimpPenaeusmonodon[J].Antiviral Res,2004,61(2):93 -99.
[16] 章跃陵,罗芸,彭宣宪.血蓝蛋白功能研究新进展[J].海洋科学,2007,31(2):77-79.
[17] Kawano T,Pinontoan R,Hosoya H,et al.Monoamine-dependent production of reactive oxygen species catalyzed by pseudoperoxidase activity of human hemoglobin[J].Bioscience,Biotechnology,and Biochemistry,2002,66(6):1224-1232.
[18] 王学琦,范启修,王者福,等.血红蛋白氧化生成的自由基对自身红细胞老化作用的初步研究[J].中国病理生理杂志,1991,7(1):42-45.
[19] 李得加,蔡小强,王利玲,等.锰取代血红蛋白的制备及其类酶活性的研究[J].武汉大学学报(理学版),2004,50(2):239-242.
[20] 王全林,刘志洪,蔡汝秀,等.血红蛋白的过氧化物酶催化特性研究[J].化学学报,2003,61(1):34-39.
Purification and antibacterial activity of hemoglobin from Tegillarca granosa
Wang Sufang1,2,Bao Yongbo2,Shi Miaojiang2,Zheng Danni2,Yang Tingting1,2,Lin Zhihua2
(1.School of Marine Sciences,Ningbo University,Ningbo 315211,China;2.Zhejiang Key Laboratory of Aquatic Germplasm Resources,Zhejiang Wanli University,Ningbo 315100,China)
Two kinds of hemoglobins were purified by gel chromatography with a recovery 74.3%.They were identified as Tg-HbⅠ(homodimer)and Tg-HbⅡ(heterogeneous tetramer)by Tricine-SDS-PAGE and mass spectrometer.The agar diffusion method and spectrophotometric method were used to detect their antibacterial activity and peroxidase activity,respectively.Results showed that both hemoglobins exhibited antibacterial activities againstEschetichia coliandPseudomonas putida.The MIC values of Tg-HbⅡtowardsE.coliandP.putidawere 0.063 mg/mL and 0.13 mg/m L,while corresponding data of Tg-HbⅠwere 0.048 mg/m L.Both hemoglobins showed peroxidase activities,which could catalyze various of phenolic substances,such as guaiacol,pyrocatechol,hydroquinone,phenol,L-DOPA,etc.Both hemoglobins lost their antibacterial activities againstP.putidain the presence of GSH,indicating that they played the antibacterial abilities through their peroxidase activities.
Tegillarca granosa;hemoglobin;antibacterial activity;peroxidase activity
S944.4+4
A
0253-4193(2014)12-0067-07
王素芳,包永波,施淼江,等.泥蚶血红蛋白的制备及其抗菌活性研究[J].海洋学报,2014,36(12):67—73,
10.3969/j.issn. 0253-4193.2014.12.006
Wang Sufang,Bao Yongbo,Shi Miaojiang,et al.Purification and antibacterial activity of hemoglobin fromTegillarca granosa[J].Acta Oceanologica Sinica(in Chinese),2014,36(12):67—73,doi:10.3969/j.issn.0253-4193.2014.12.006
2014-01-15;
2014-03-27。
国家863计划项目(2012AA10A410);宁波市自然基金项目(2013A610161);浙江省教育厅高校科研项目(Y201225951);国家级大学生创新创业训练计划项目(201310876018);宁波市海洋贝类科技创新团队(2011B82017);水产科学浙江省重中之重学科开放基金项目。
王素芳(1977—),女,山东省单县人,博士,主要研究水产资源的开发与利用。E-mail:24274414@qq.com
*通信作者:包永波,主要从事贝类功能基因和蛋白组研究。E-mail:bobbao2001@gmail.com
猜你喜欢
河北科技师范学院学报(2022年2期)2022-08-26
纺织科技进展(2021年5期)2021-07-22
陶瓷学报(2021年1期)2021-04-13
军事文摘(2020年20期)2020-11-16
中学生数理化·八年级物理人教版(2020年12期)2020-01-01
今日农业(2019年10期)2019-06-26
中学生数理化·八年级物理人教版(2018年12期)2019-01-31
中国塑料(2016年3期)2016-06-15
中国卫生标准管理(2015年24期)2016-01-14
华东理工大学学报(自然科学版)(2015年4期)2015-12-01
