
鄂尔多斯盆地西缘奥陶纪生物礁基本特征、分布规律及成礁模式*
2014-05-30 07:14赵俊兴李凤杰朱广社苏中堂邹敏王玉萍周俊烈
岩石学报 2014年3期
赵俊兴 李凤杰 朱广社,3 苏中堂 邹敏 王玉萍 周俊烈
1.油气藏地质及开发国家重点实验室成都理工大学,成都 610059
2.成都理工大学沉积地质研究院,成都 610059
3.中国石油长庆油田分公司第三采油厂,银川 750006
1 引言
鄂尔多斯盆地奥陶系海相碳酸盐岩成为近年来盆地油气勘探热点,具有丰富的油气资源与广阔的勘探前景(席胜利等,2006;夏明军等,2008;杨华等,2011;赵俊兴等,2012)。生物礁是指由原地生长的造礁生物营造的碳酸盐岩有机建造,具有抗浪格架,呈现凸镜状或丘状的外部形态,并突出于四周同期沉积物(Camoin and Montaggioni,1994;Camoinet al.,1999;范嘉松,1996;Riding,2002;Stanley,2003)。显生宙各地质历史时期皆有生物礁发育,受地史时期古板块纬度、古气候变化、海相地层发育差异的影响而导致生物礁分布在时代和区域上的极度不均匀性(Wilson,1974;Montaggioni and Camoin,1993;Flügel and Kiessling,2002)。生物礁是一种非常好的油气储集体,生物礁和相关的碳酸盐岩中蕴含着大约一半的油气储量(范嘉松和张维,1995;范嘉松,1996;付金华等,2012);生物礁沉积建造在形成过程中会记录一系列的古环境、古沉积条件、古生物等方面的重要信息(钟建华等,2005;王瑞等,2009)。我国自20世纪中后期开始开展了生物礁的调查与研究,发现了从震旦纪到新近纪发育分布范围很广的生物礁(刘春燕等,2007);叶俭等(1995)在20世纪90年代初对鄂尔多斯盆地南部地区的生物礁进行了研究,发现了多处生物礁,并对这些生物礁的形成、发展和演化进行了较为系统的总结,但目前对于盆地西缘生物礁研究相对薄弱。本次油气地质调查主要位于鄂尔多斯盆地西缘地区,北起内蒙古乌海、南至宁夏固原彭阳,主要涉及盆地西缘冲断带及天环坳陷(图1)。笔者在新一轮碳酸盐岩油气地质调查的基础上,对鄂尔多斯盆地西缘碳酸盐岩野外剖面及钻井岩心进行了系统调查,在调查过程中发现了露头较好的生物礁,并对各个典型生物礁特征进行了实际测量及鉴定分析和对比,揭示了盆地西缘生物礁分布规律,建立了成礁模式,探讨了礁消亡原因,这对鄂尔多斯盆地奥陶系碳酸盐岩沉积环境分析和油气勘探都具有重要的意义。
2 西缘生物礁的基本特征
通过本次对盆地西缘碳酸盐岩油气地质调查,认为鄂尔多斯盆地西缘奥陶纪发育了生物礁,在平面上主要分布于桌子山、棋探1井和石节子沟等地,成南北向带状展布,生物礁发育典型,证实了盆地西缘生物礁的存在(图1)。生物礁发育时代主要为奥陶纪中晚期,该时期盆地周缘碳酸盐岩台地具备“清、浅、暖”的沉积环境,这种台地适宜生物的生长,尤其是在碳酸盐岩台地的陡缓变化带上,因不断承受广海海水的冲击,海水新鲜,营养丰富,适宜造礁生物的生长。
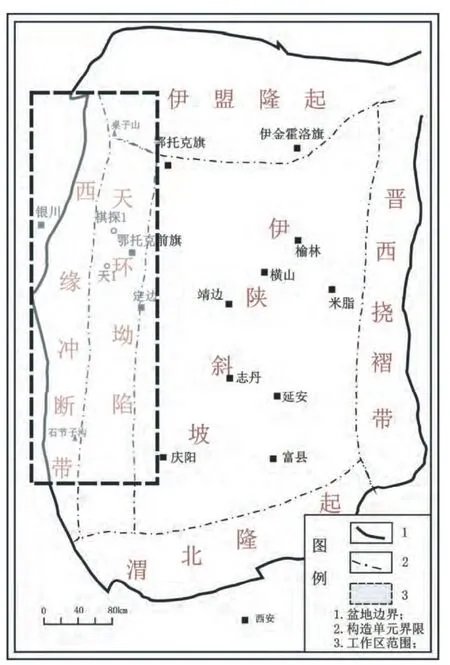
图1 鄂尔多斯盆地西缘构造分区与研究区位置Fig.1 Structural division on the western margin of the Ordos Basin and location of research area
2.1 西缘北段生物礁-桌子山生物礁特征
奥陶系生物礁剖面位于内蒙古自治区乌海市海南区老石旦镇东山,礁体发育层位为下奥陶统桌子山组顶部,为一套开阔台地、台地边缘浅滩及陆棚沉积组合,上覆地层为克里摩里组。通过本次实际测量将此生物礁发育段共分成7个小层(图2)。礁体主要出现在第1层、第5层和第6层。礁核为块状碳酸盐堆积体,呈丘状、“似层状”。主要造礁生物为横板珊瑚(郑昭昌和李玉珍,1991),其次为层孔虫;附礁生物种类繁多,含有角石、腹足、腕足、海百合等;共见有3期礁体生长。

图2 内蒙古桌子山生物礁剖面及照片Fig.2 Profile of organic reef and pictures in the Zhuozishan area,Inner-Mongolia
第一期礁体生长:礁核(第1层)厚5.69m,含生物碎屑珊瑚灰岩,上部有明显的岩溶现象,裂缝非常发育,礁体形态明显,呈丘状;礁基(第0层)为薄层状泥灰岩;礁盖(第2层)厚2.30m,为深灰色泥岩,页岩夹瘤状灰岩,中间夹薄层灰泥透镜体、生物碎屑,水平层理,为浅水陆棚到深水陆棚环境。其后发育了含生物碎屑的滩相沉积(第3层、第4层)厚约6m,上下层整齐,里面发育小型韵律层,颗粒主要为鲕粒、生物碎屑等。
第二期礁体生长:礁基(第4层顶)为薄层状含生物碎屑灰岩,为碎屑滩沉积,顶部为泥灰岩薄层条带;礁核(第5层)厚6.0m,中、厚层状珊瑚灰岩,“似层状”块体,无层理、无成层性的块状碳酸盐堆积,礁体形态明显,呈丘状;礁盖、礁坪(第5层顶部)厚约0.5m,为生物碎屑灰岩,见鲕粒,见有波浪打碎的角砾状礁岩结构,顶部夹薄层灰泥透镜体。
第三期生物礁:礁核(第6层)厚6.89m,是在底部经压实呈瘤状的泥质条带上发育起来的。中、上部为灰色厚层块状灰岩,单层厚度30~50cm,造礁生物为横板珊瑚和层孔虫,附礁生物化石主要为头足类,腹足类,以及海百合,含颗粒、内碎屑、鲕粒、砂屑,局部可见硅质结核及生物化石硅化,下部泥岩颜色浅,氧化后泛红,上部层孔虫较多。

图3 棋探1井生物礁剖面及照片Fig.3 Profile and pictures of organic reefs in Qitan1 Well
在桌子山组生物礁生长之后,紧接着沉积了克里摩里组深灰色薄层瘤状灰岩与深灰色泥岩互层沉积,韵律层发育,为一套典型的深水陆棚沉积,有生物遗迹构造,含笔石化石。在此笔者认为桌子山组顶部生物礁死亡系克里摩里组沉积海平面快速上升事件导致礁体被淹死所致,也进一步推测在桌子山组-克里摩里组之间存在一次海平面快速上升事件。桌子山生物礁处在有利于礁滩储层发育的台地边缘相带,与上覆克里摩里组、乌拉力克组深水沉积的烃源岩相邻,气源充足,生、储、盖组合配套条件好,是盆地西缘有利的油气勘探对象。
2.2 西缘中段生物礁-棋探1井生物礁特征
钻井岩心观测过程中,棋探1井在克里摩里组钻遇生物礁(取心回次为8~9次),是盆地西缘中段仅有的生物礁体发现(图3)。棋探1井主要造礁生物为层孔虫、海绵及钙藻等;附礁生物有介形虫、三叶虫、腹足、棘皮类、腕足等;礁体主要的岩相组合有生物屑碎屑灰岩、层孔虫-海绵礁灰岩。取芯中发现的海绵骨针,形态分散,不构成生物骨架,与丝状蓝细菌、介形虫、腹足类、棘皮类等共生,说明该段地层为浅海环境,海绵骨针与浅海生物共生时,判断沉积相应当以浅海中水体较深环境。
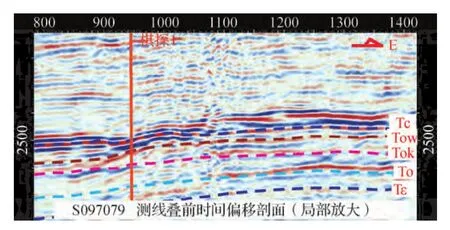
图4 S097079测线叠前时间偏移剖面的礁滩相特征Fig.4 Reef-bank facies characteristics of line S097079 in prestack time migration seismic section
该生物礁在地震剖面上地震相特征明显,总体上表现为较强振幅不连续反射和丘形反射特征(图4),与厚层块状白云岩或灰岩的中弱反射特征低频弱振幅弱连续性有较大差异,是识别礁滩体的主要依据(曾令帮等,2011)。
棋探1井生物礁发育在深水沉积的黑灰色云灰岩之上,随着水体变浅,适宜造礁生物生长,岩心观测中可见明显的藻纹层构造,藻类生物生长迅速,形成了层孔虫-海绵生物礁;此后,水动力作用加强,水体加深,生物礁骨架受到破坏,形成生物碎屑灰岩,随着水体再次变浅,水动力条件变缓,适宜造礁生物生长,形成了层孔虫-海绵礁灰岩。

图5 彭阳县石节子沟上奥陶统生物礁剖面Fig.5 Profile of Upper Ordovician organic reefs in Shijiezigou,Pengyang County
在克里摩里组生物礁生长之后,紧接着沉积了乌拉力克组灰黑色泥页岩,为一套典型的深水陆棚沉积,有机质含量高,富含笔石化石,是一套较好的烃源岩。同样认为克里摩里组生物礁死亡系沉积海平面快速上升事件导致礁体被淹死所致。该生物礁与上覆乌拉力克组深水沉积的烃源岩(笔石页岩)相邻,气源相对充足,生、储、盖组合配套条件好,但生物礁发育规模不大,也是盆地西缘较为有利的油气勘探对象。
另外,在盆地西缘中段的那1井中的第11次取芯发现有生物灰岩,虽然有海绵等生物,藻类含量高,但笔者认为那1井为藻粘结的生物丘灰岩,不具备礁体的典型特征,在此不做讨论。
2.3 西缘南段生物礁-彭阳石节子沟生物礁特征
该生物礁出露与石节子沟庠水坝下,向两侧延伸的可见范围有限,上部被黄土覆盖,与其紧邻下伏地层平凉组为灰色泥岩夹灰黄色砂岩,笔者认为是一套碎屑潮坪环境沉积。
该生物礁造礁生物为珊瑚,附礁生物有腹足、腕足及介形虫等。珊瑚有单管珊瑚、四射珊瑚及床板珊瑚(Plasmopotellasp.,Agetilitesgrandis,Cateniporasp.,Favistella grandis Lin)等(安太庠和郑昭昌,1990),腹足、腕足见到的大多生物碎屑,郑昭昌和李玉珍(1991)还识别出牙形石Panderodussp.,Panderiodussp.,Subcordylodiform等,最初将其定为生物灰岩,而没有作为生物礁予以点名。礁体可见总厚度89.5m,从造礁生物繁育情况可大致将该生物礁分成四层,13个小层(图5)。
第一期生物礁发育在生物灰岩之上,包括1~6小层,厚度约43.5m,造礁生物为珊瑚,从生屑滩上发育而成,该礁体珊瑚个体大,质纯,形成的岩层向上逐渐变薄,表明水体逐渐加深不利于珊瑚生长,附生生物相对发育,形成生物碎屑灰岩和生物灰岩,该种环境经历了一段较长时期。
第二期生物礁的发育可能是生物碎屑逐渐生长使得水体变浅适宜珊瑚生长,从而在相对较长的一个时期内发育,形成生物礁灰岩。该层包括7、8小层,厚17m,造礁生物依旧是珊瑚,但较上层珊瑚个体变小,但数量有所增加,随后发育中层生物灰岩,说明水体再次变深,使得环境不利于珊瑚生长,从而结束了该层生物礁的生长。
第三期生物礁包括9、10两小层,厚10.5m,生物礁礁核由造礁生物珊瑚形成质纯的生物礁灰岩,该层内珊瑚发育跟第二层相似,个体小,数量相对多,但该层生物礁变化快,层厚薄,表明水体深浅变化频率加快,环境变化明显。

图6 鄂尔多斯盆地西缘典型生物礁时空分布对比图Fig.6 Contrast diagram of the typical reefs on the western margin of the Ordos Basin
第四期生物礁包括11~13层,厚19.5m,顶部发育一套岩溶角砾岩,基岩为中细晶灰岩,少见化石,可能是礁间沉积。而该层生物礁礁体本身厚仅14.5m,且生物礁灰岩较薄,生物灰岩相对发育。造礁生物为珊瑚,此层的珊瑚个体更小,但数量增多,且以单管珊瑚为主,表明此时环境非常适宜珊瑚生长,但环境变化快,不利于形成个体较大珊瑚。
生物礁顶部为第四纪黄土覆盖,上覆岩层特征不清。
3 盆地西缘生物礁时空分布规律
在前期对盆地西缘生物礁实际测量、特征分析的基础上,在此总结了盆地西缘生物礁发育特征及分布规律(图6),为认识盆地西缘奥陶系沉积环境、生物礁分布预测和礁滩型储层油气勘探具有十分重要的意义。对比研究认为有如下规律特征:
(1)平面上:盆地西缘(即华北版块西缘)发育礁滩相带,为点状生物礁沿盆地西缘天环拗陷呈南北向条带状分布;
(2)时间上:生物礁发育在奥陶纪。北部生物礁体发育层位早,主要位于早奥陶世晚期的桌子山组顶部,中部生物礁发育层位相对晚,主要在克里摩里组,南部生物礁发育最晚,位于上奥陶统背锅山组;
(3)造礁生物:造礁生物北部为珊瑚,中部为海绵-层孔虫,南部为珊瑚,且个体较大;
(4)成礁期次及发育规模:北段礁体生长发育2~3期,中段发育2期,南段发育可达4期;北端及南端生物礁生长的水体环境相对较浅,礁体规模较大,中部相对较深,礁体规模小。
前人研究证实,在早古生代早期至晚期,华北板块由赤道附近南部向北发生漂移,且发生了一定程度的右旋。鄂尔多斯盆地位于华北板块西缘,那么盆地西缘早古生代生物礁的这种分布变化规律与华北板块在早古生代处于赤道附近发生了北漂右旋的旋转漂移规律有关,认为是早期板块北段处于赤道附近有利于造礁,晚期板块南端处于赤道附近而利于成礁。
4 盆地西缘生物礁成礁模式及死亡原因探讨
4.1 生物礁成礁模式
鄂尔多斯盆地西缘生物礁的发育与演化过程各有特点,南、中、北段各有不同。北段桌子山生物礁体发育层位最老,于奥陶系下统的桌子山组顶部,发育层位最早;棋探1井生物礁发育层位主要在奥陶系下统顶部克里摩里组(桌子山上部地层);南端石节子沟生物礁发育于上统背锅山组,发育时代最晚。不同时期的生物礁出现在同一构造带不同地段,这种情况表明鄂尔多斯盆地西缘生物礁的明显发育受到活动性断裂、板块旋转、海平面变化等的控制,这些活动性断裂的多期活动使该区海平面时升时降,生物礁不能稳定生长发育,造成多期旋回的成礁序列。同时断裂构造的不均匀活动、华北板块间歇性北漂右旋使各个断块出现不同的情况,有些断块逐渐隆起,水体变浅,而有些断块则逐渐下降,使水体变深,导致生物礁最终消亡,不同地区生物礁发育特征也不尽相同。

图7 鄂尔多斯盆地西缘奥陶系生物礁成礁模式图Fig.7 Reef formation model of the Ordovician organic reef in the western margin of the Ordos Basin
奥陶纪早期统一沉积的潮坪,在经历海平面上涨后,不仅海水分布面扩广,水体加深,同时由于构造运动,地层出现明显差异,有的地方隆起,有的地方凹陷。隆起处水浅,光照条件好,适合生物生长、繁殖,在此便有生物礁发育。但由于隆起处地形也有差异,因此各地造礁生物也不尽相同。凹陷处水深,沉积有薄层泥晶灰岩,靠近断裂处有碎屑流沉积,成礁模式为靠近贺兰坳拉槽东缘的陆缘海背景下的开阔台地边缘带点礁-滩体组合模式(图7)。礁滩型储层形成于台地边缘高能相带,其空间展布受控于台地边缘相带的展布,在该相带不同的微相内礁滩型储层储集性能不同,表现出一定的相控性,如生物礁孔隙发育但渗透性较差,颗粒滩间灰岩物性条件均差,颗粒滩孔隙度和渗透率相对均好(付金华等,2012;周俊烈等,2013)。在鄂尔多斯盆地西缘奥陶系的三个典型生物礁上覆地层岩性均为深灰色、灰黑色深水相的含笔石泥页岩沉积,韵律层发育,为一套典型的深水陆棚沉积,是礁滩相储层的良好盖层。
4.2 盆地西缘奥陶系生物礁死亡原因探讨
4.2.1 生物礁死亡原因综述
生物礁通常作为指相和古气候的标志,是一种正常条件下沉积产物。礁的研究表明,一个礁的生长与终止,往往受地壳运动、海水进退、气候变化等条件影响(Flügel and Kiessling,2002;张永刚等,2011)。导致生物礁死亡通常都与突变事件有关,通常有这么几种情况:一是礁盖层为深海相沉积物,无侵蚀面,浅海礁相与深海相直接接触的此种情况,显然是突变事件,是陆棚迅速沉降、海平面快速上升事件的结果;二是礁盖层为砂泥岩等陆源碎屑岩,礁停止生长后为大量陆源碎屑岩沉积,显然也是一种突变,表明海水变浅或有河流带来泥砂使盆地充填;三是礁盖层为蒸发岩,也是一种突变,表明气候突然变干燥,海水盐度增高,属气候突变事件引起;四是火山作用事件,由于火山灰影响阳光、氧气等原因而导致生物礁死亡。
4.2.2 生物礁相碳酸盐岩碳氧同位素地球化学特征
生物礁中δ13C值的变化与生物埋藏量以及埋藏速率有很密切的关系。造礁生物的繁盛及绝灭,反映在δ13C值的变化上,即氧化碳与还原碳的转化,在成礁过程中,造礁生物大量繁殖,对应海洋中δ13C值的升高(陈鹤等,2008)。
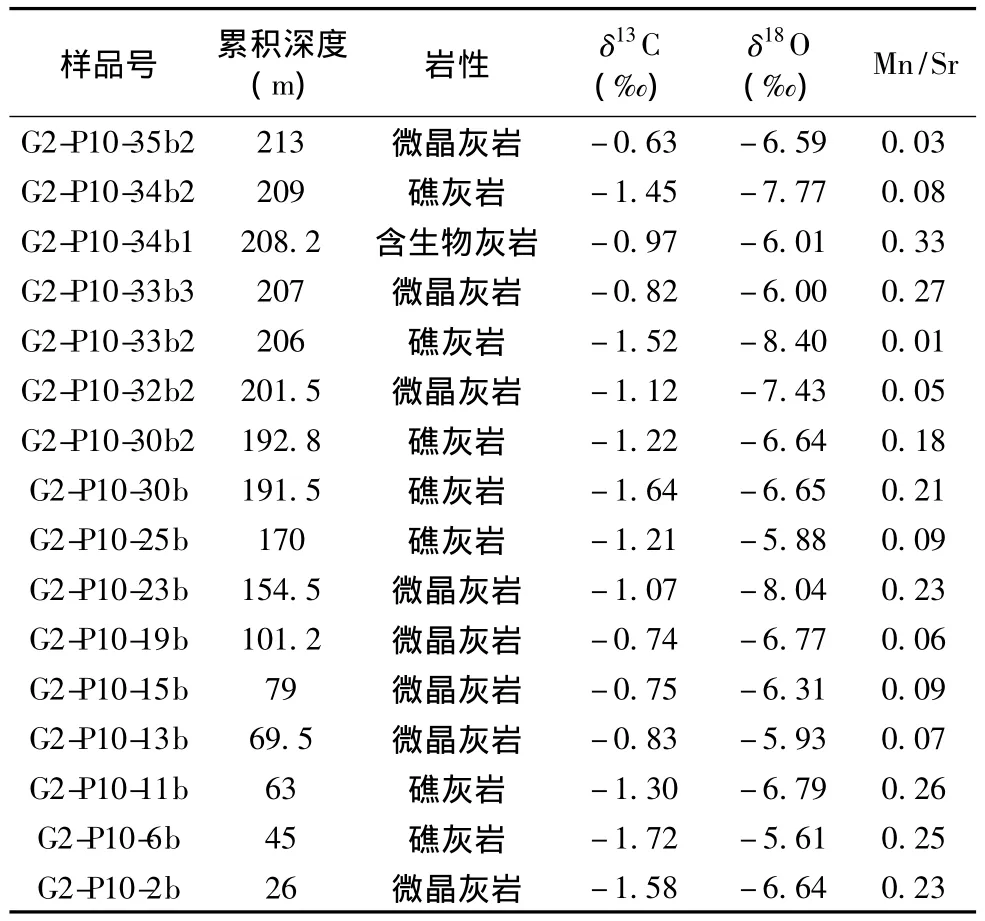
表1 乌海老石旦东山桌子山组生物礁碳氧同位素分析数据Table 1 Carbon and oxygen isotope data of organic reef of the Zhuozishan Formation Laoshidan east hill in Wuhai County,Ordos Basin
本次研究在桌子山剖面共采集新鲜岩石样品16块,所取样品主要桌子山组生物礁的岩性,碳氧同位素分析数据见表1。在样品采集过程中,避开了岩脉充填部位,并尽量排除结晶程度较高的岩石样品,以保证样品的可用性。样品送中国科学院地质与地球物理研究所兰州油气资源中心,碳氧同位素分析采用磷酸法,在Finnigan MAT252型稳定同位素比质谱仪上进行,δ13C和δ18O的标准偏差分别为0.05‰和0.07‰。为了判断样品的碳氧同位素组成是否遭受过沉积期后的改造,还进行了Mn、Sr含量的测定。
碳酸盐岩样品的碳氧同位素分析数据在应用之前,首先要判断其在成岩过程中是否发生了蚀变,通常判断标准有3个,包括:Mn/Sr、δ18O 组成、δ13C 和 δ18O 的相关性。Mn/Sr小于2~3,表示样品很好地保持了原始的碳、氧同位素组成(郝松立等,2011);δ18O<-10‰代表岩石已发生了强烈的蚀变,不能代表原始的碳、氧同位素组成,如果δ13C和δ18O数值不具有明显的相关性,则表明海相碳酸盐岩基本保持了原始的碳、氧同位素组成(黄思静等,2003;郝松立等,2011)。
对所取16件样品的Mn/Sr比值进行了计算,最大值仅为0.33,远小于2~3的范围(表1)。所测定的δ18O最小值为-8.40‰,大于-10.0‰,表明样品没有发生强烈的蚀变(表1);对样品的δ13C和δ18O相关性分析,可以得出其相关系数仅为0.21(图8),不具有明显的相关性。由此说明样品受后期成岩作用影响较小,基本保持了原始的碳、氧同位素组成。
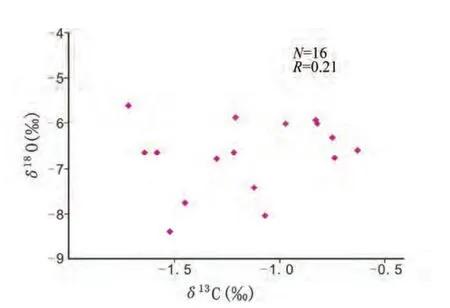
图8 乌海老石旦东山桌子山组碳酸盐岩δ13C和δ18O值相关图Fig.8 The relationship between δ13C and δ18O of carbonates from Loahsidan east hill in Wuhai County,Ordos Basin
在鄂尔多斯西缘奥陶系桌子山组样品中,礁灰岩的δ13C的均值为-1.37‰,而普通灰岩的均值为-0.987‰,高于礁灰岩。这是因为在礁体的形成过程中,造礁生物大量繁殖,在比较合适的环境下,这些造礁生物会大量吸收轻同位素12C,使得重同位素13C在水体中含量升高(陈鹤等,2008;郝松立等,2011),在13C含量丰富的海水中形成的岩石其13C的含量也会相应升高(郝松立等,2011)。因此,生物含量丰富的礁灰岩因生物体本身吸收了大量的12C而呈现δ13C的低值;相应地,生物含量少的非礁灰岩则呈现δ13C的高值(郝松立等,2011)。
通过δ13C值的变化可以反映海平面的变化(Lee and Carpenter,2001;陈鹤等,2008;郝松立等,2011);其中的δ13C的高值对应着海平面的上升期,δ13C的低值对应着海平面的下降期(Lee and Carpenter,2001;郝松立等,2011)。从图9可以看出,鄂尔多斯盆地西缘桌子山组碳酸盐岩δ13C由较低的-1.58‰上升为-0.74‰,之后虽然呈现高低交替变化,但总体呈现出变高的趋势。反映了在桌子山组礁灰岩沉积后,海平面上升的变化趋势。
4.2.3 生物礁死亡原因探讨
鄂尔多斯盆地奥陶系西缘生物礁的根据礁体上覆地层、岩性等实际情况,尤其是从乌海老石旦东山桌子山组剖面上来看,该剖面从下而上生物含量逐渐增加,尤其在23层~33层内珊瑚丰度明显增加,可形成抗浪群落,而从C同位素曲线上也显示出该段处于海平面下降时期,时间上为桌子山组晚期,一方面说明在桌子山晚期珊瑚开始规模生长,可形成抗浪体,另一方面说明海平面下降利于珊瑚的生长,从而形成礁体。再向上至克里摩里期海平面上升,导致环境不利于珊瑚生长,在剖面上表现为生物丰度降低,层厚变薄。同时鄂尔多斯盆地西缘三个典型生物礁上覆地层岩性主要为深灰色、灰黑色深水相的含笔石泥页岩沉积,韵律层发育,为一套典型的深水陆棚沉积,说明桌子山地区早奥陶世中晚期生物礁是随海平面快速上升事件导致礁体被淹死所致,同时也进一步推测生物礁之上存在一次海平面快速上升事件。
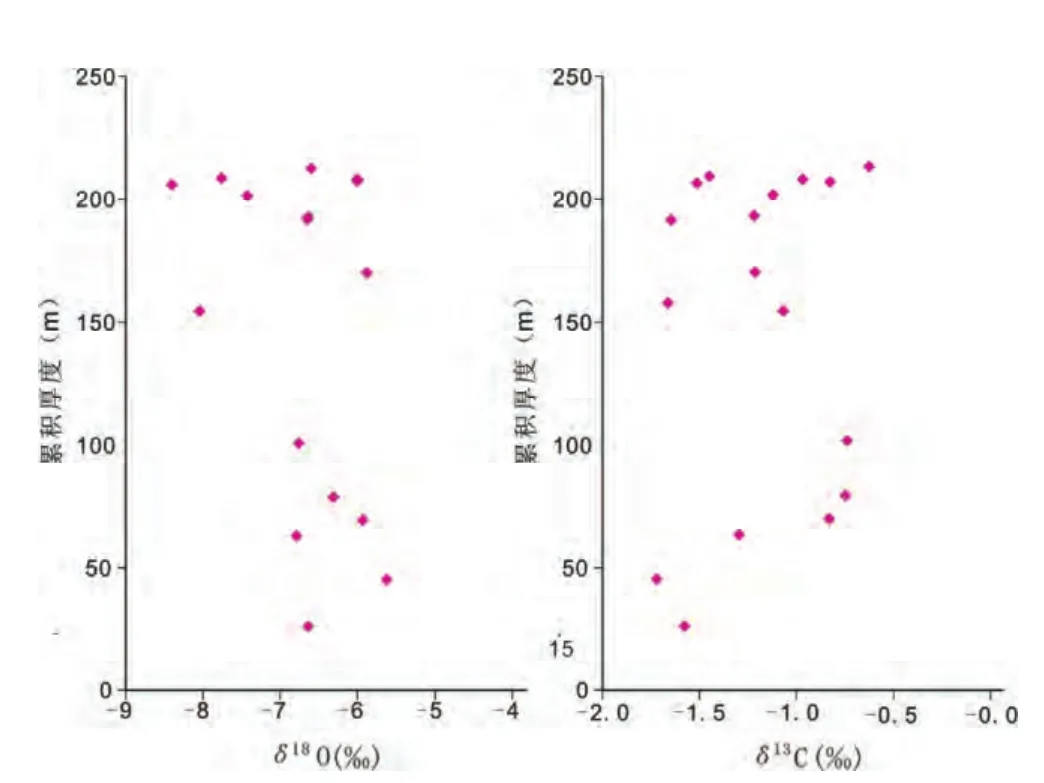
图9 乌海老石旦东山桌子山组碳酸盐岩δ13C和δ18O值分布图Fig.9 Distribution of δ13C and δ18O of carbonates from Laoshidan east hill in Wuhai County,Ordos Basin
5 结论
通过本次鄂尔多斯盆地西缘地区新一轮野外露头和钻井岩心碳酸盐岩油气地质调查过程,重点对盆地西缘典型的生物礁特征并进行了实际测量。通过本次对各个典型生物礁的发育特征进行了分析和分布规律研究,可以得出以下结论:
(1)认为鄂尔多斯盆地西缘地区发育有较为典型的生物礁,但发育数量少且分布局限。分别分布在西缘北段桌子山、中段的棋探1井和南段的石节子沟等地,西缘奥陶系生物礁与各种滩共生,发育了礁滩型储层。
(2)北段生物礁体发育层位早,中部生物礁发育层位相对晚,南部生物礁发育最晚。这种分布变化规律与华北板块在早古生代处于赤道附近发生了北漂右旋的旋转漂移规律有关,认为是早期板块北段处于赤道附近有利于造礁,晚期板块南端处于赤道附近而利于成礁。
(3)造礁生物主要为珊瑚、层孔虫和海绵,附礁生物为腹足类、腕足、角石、棘皮类等,三处生物礁发育特征有差异。北部为珊瑚、层孔虫造礁,中部为海绵-层孔虫造礁,南部为珊瑚造礁,且个体较大;北段礁体生长发育2~3期,南段发育可达4期;北端及南端生物礁生长的水体环境相对较浅,中部相对较深。
(4)建立了西缘生物礁成礁模式,认为盆地西缘生物礁成礁模式为中央古隆起西部陆缘海背景下的开阔台地边缘带点礁-滩体组合模式,生物礁分布局限。西缘生物礁油气成藏条有利,生、储、盖组合配套良好,是鄂尔多斯盆地的碳酸盐油气勘探重要目标。
(5)认为该地区生物礁死亡原因系海平面快速上升导致礁体被淹死所致,同时推测桌子山组顶部生物礁之上的克里摩里组存在一次海平面快速上升事件。
An TY and Zheng ZC.1990.The Molar-Shaped Rocks around the Ordos Basin.Beijing:Science Press,1-201(in Chinese)
Camoin GF and Montaggioni LF.1994.High energy coralgal-stromatolite frameworks from Holocene reefs(Tahiti, French Polynesia).Sedimentology,41(4):655-676
Camoin GF,Gautret P,Montaggioni LF and Cabioch G.1999.Structure and composition of organic reefs and carbonate mud mounds:Concepts and categories.Sedimentary Geology,126(1 - 2):271-304
Chen H,Gong EP,Guan CQ,Sun BL and Zhang YL.2008.Application of constitution of carbon,oxygen and strontiumisotos and trace elements in study of paleoenvironment of reefs.Global Geology,27(2)130-136(in Chinese with English abstract)
Fan JS and Zhang W.1985.On the basic concept and classification of organic reefs and their main identifying criteria.Acta Petrologica Sinica,1(3):45-49(in Chinese with English abstract)
Fan JS.1996.Reefs and Petroleum of China.Beijing:Ocean Press,1 -210(in Chinese)
Flügel E and Kiessling W.2002.A new look at ancient reefs.In:Kiessing W,Flügel E and Golonka J(eds.).Phanerozoic Reef Patterns.SEPM Special Publication,72:1-50
Fu JH,Bai HF,Song LY and Ma ZR.2012.Types and characteristics of Ordovician carbonate reservoir in Ordos Basin,China.Acta Petrolei Sinica,33(Suppl.2):110 - 117(in Chinese with English abstract)
Hao SL,Li WH,Liu JP,Chen Q,Li Y,Zhu J,Tan CX and Zhang HY.2011.Characteristics of carbon and oxygen Isotopes geochemistry of organic reef facies carbonates of Ordovician in southern margin of Ordos.Acta Geological Science and Technology Information,30(2):52-56(in Chinese with English abstract)
Hang SJ,Shi H,Mao XD,Zhang M,Shen LC and Wu WH.2003.Palaeozoic marine carbonation and preservation for the infaomation of seawater.Journal of Chengdu University of Technology(Science&Technology Edition),30(1):9-17(in Chinese with English abstract)
Lee D and Carpenter SJ.2001. Isotopic disequilibrium in marine calcareous algae.Chemical Geology,172(3-4):307-329
Liu CY,Liu CS,Wu MBet al.2007.Characteristics of spatiotemporal distributions of reefs in China and their geological significance.Global Geology,26(1):44 -51(in Chinese with English abstract)
Montaggioni LF and Camoin GF.1993.Stromatolites associated with coralgal communities in Holocene high-energy reefs.Geology,21(2):149-152
Riding R.2002.Structure and composition of organic reefs and carbonate mud mounds:Concepts and categories.Earth Science Reviews,58(1-2):163-231(in Chinese with English abstract)
Stanley GD Jr.2003.The evolution of modern corals and their early history.Earth-Science Reviews,60(3-4):195-225
Wang R,Jiao YQ,Wu LQet al.2009.Reef-forming model of Honghua reefs of the Upper Permian Changxing Formarion in Kaixian,Chongqing.Journal of Palaeogeography,11(2):157 - 166(in Chinese with English abstract)
Wilson JL. 1974. Carbonate Facies in Geologic History. Berlin,Heidelberg,New York:Springer-Verlag,96-102
Xi SL,Li ZH,Wang X and Zheng CB.2008.Distribution and exploration potential of Ordovician reservoir in Ordos basin.Oil and Gas Geology,27(3):406-412(in Chinese with English abstract)
Xia MJ,Zheng CB,Bi JX,Zeng ZQ,Guo HX,Wei LX,Song L,Miao Q,Teng YC and Miao B.2008.Ordovician reefs and the prospect of gas,Ordos Basin.Natural Gas Geoscience,19(2):178 -182,271(in Chinese with English abstract)
Yang H,Fu JH,Wei XS and Ren JF.2011.Natural gas exploration domains in Ordovician marine carbonates, Ordos Basin. Acta Petrolei Sinica,32(5):733 -739(in Chinese with English abstract)
Ye J,Yang YY and Xu AD.1995.The Organic Reef of Ordovician in Southwestern Margin of Ordos Basin.Beijing:Geological Publishing House,1-242(in Chinese)
Zeng LB,Duan YS,Bi MB,Zhu WM and He ZG.2011.Seismic identification and exploration effect of reef-bank facies reservoir in northwestern Ordos Basin.Lithologic Reservoirs,23(2):75-79
Zhang YG,He ZH,Chen HD and Liu SG.2011.Typical Marine Reef Reservoirs in China.Beijing:Science Press,1 -100(in Chinese)
Zhao JX,Li J,Lu T,Feng QH,Wang Y,Li FJ and Shen ZJ.2012.Reservoir controlling factors and accumulation regularity of Member 5 of Majiagou Formation in Sulige region of Ordos basin,China.Journal of Chengdu University of Technology(Science&Technology Edition),39(4):388-394(in Chinese with English abstract)
Zheng ZC and Li YZ.1991.The new advances in the study of the Ordovican System in Helanshan area.Geoscience,5(2):119-137(in Chinese with English abstract)
Zhong JH,Wen ZF,Li Yet al.2005.Organic reefs study:Concept,classification,characteristics,history and development.Geological Review,51(3):288-298(in Chinese with English abstract)
Zhou JL,Cheng HD,Zhao JX,Su ZT,Zhang CG,Wang YP and Hao ZM.2013.The Ordovician reef and reservoir performance evaluation in western margin of Ordos Basin.Journal of Chengdu University of Technology(Science& Technology Edition),40(2):193-199(in Chinese with English abstract)
附中文参考文献
安太庠,郑昭昌.1990.鄂尔多斯盆地周缘的牙形石.北京:科学出版社,1-201
陈鹤,巩恩普,关长庆,孙宝亮,张永利.2008.C、O、Sr同位素及微量元素组成在生物礁礁体环境研究中的应用.世界地质,27(2):130-136
范嘉松,张维.1985.生物礁的基本概念、分类及识别特征.岩石学报,1(3):45-49
范嘉松.1996.中国生物礁与油气.北京:海洋出版社,1-30
付金华,白海峰,孙六一,马占荣.2012.鄂尔多斯盆地奥陶系碳酸盐岩储集体类型及特征.石油学报,33(增刊2):110-117
郝松立,李文厚,刘建平,陈强,李莹,朱静,谭晨曦,张慧元.2011.鄂尔多斯南缘奥陶系生物礁相碳酸盐岩碳氧同位素地球化学特征.地质科技情报,30(2):52-56
黄思静,石和,毛晓冬,张萌,沈立成,武文慧.2003.早古生代海相碳酸盐的成岩蚀变性及其对海水信息的保存性.成都理工大学学报(自然科学版),30(1):9-17
刘春燕,林畅松,吴茂炳等.2007.中国生物礁时空分布特征及其地质意义.世界地质,26(1):44-51
王瑞,焦养泉,吴立群等.2009.重庆开县上二叠统长兴组红花生物礁成礁模式.古地理学报,11(2):157-166
席胜利,李振宏,王欣,郑聪斌.2006.鄂尔多斯盆地奥陶系储层展布及勘探潜力.石油与天气地质,27(3):405-412
夏明军,郑聪斌,毕建霞,曾正清,郭海霞,魏立新,孙利,苗菁,滕英翠,苗兵.2008.鄂尔多斯盆地奥陶系生物礁及其天然气勘探前景.天然气地球科学,19(2):178-182,271
杨华,付金华,魏新善,任军峰.2011.鄂尔多斯盆地奥陶系海相碳酸盐岩天然气勘探领域.石油学报,32(5):733-740
叶俭,杨友运,许安东.1995.鄂尔多斯盆地西南缘奥陶纪生物礁.北京:地质出版社,20-60
曾令帮,段玉顺,毕明波,朱望月,何争光.2011.鄂尔多斯盆地西北部礁滩相储层地震识别与勘探效果.岩性油气藏,23(2):75-79
张永刚,贺振华,陈洪德,刘树根.2011.中国典型海相礁滩储层.北京:科学出版社,1-50
赵俊兴,李婕,卢涛,冯强汉,王勇,李凤杰,申赵军.2012.苏里格地区马五段储层控制因素与成藏规律.成都理工大学学报(自然科学版),39(4):388-394
郑昭昌,李玉珍.1991.贺兰山奥陶系研究的新进展.现代地质,5(2):119-137
钟建华,温志峰,李勇等.2005.生物礁的研究现状与发展趋势.地质论评,51(3):288-298
周俊烈,陈洪德,赵俊兴,苏中堂,张成弓,王玉萍,郝哲敏.2013.鄂尔多斯盆地西缘奥陶纪生物礁及其储集性能评价.成都理工大学学报(自然科学版),40(2):193-199
猜你喜欢
山东冶金(2022年2期)2022-08-08
作文小学中年级(2020年10期)2020-12-29
动漫星空(兴趣百科)(2020年12期)2020-12-12
中国海上油气(2020年5期)2020-10-20
科学导报·学术(2019年21期)2019-09-10
城市道桥与防洪(2019年5期)2019-06-26
科普童话·神秘大侦探(2018年9期)2018-10-25
儿童故事画报·发现号趣味百科(2017年4期)2017-06-30
中国卫生(2016年10期)2016-11-13
西夏学(2016年2期)2016-10-26
