
春、秋季崇明东滩盐沼潮沟大型浮游动物群落分布
2014-05-27 08:00:38秦海明汤臣栋马强陈家宽吴纪华
水生生物学报 2014年2期
秦海明 汤臣栋 马强 陈家宽, 吴纪华
春、秋季崇明东滩盐沼潮沟大型浮游动物群落分布
秦海明1汤臣栋2马强2陈家宽1, 3吴纪华3
(1. 南昌大学生命科学研究院流域生态学研究所, 南昌 330031; 2. 上海崇明东滩鸟类国家级自然保护区管理处, 上海 202183; 3. 复旦大学生物多样性与生态工程教育部重点实验室, 上海 200433)
大型浮游动物; 类群组成; 时空分布; 盐沼; 潮沟; 长江口
大型浮游动物(Macrozooplankton)是指体长介于1 mm和1 cm之间的浮游动物[1]。它们主要以碎屑、细菌、藻类、原生动物和中小型浮游动物等为食, 同时本身又是鱼类、虾蟹类和水鸟等动物的饵料[2, 3], 在生态系统食物网中处于中间环节, 通过能量传递联系着生产者和次级消费者。
盐沼(Salt marsh)是温带与亚热带地区海岸带及河口受海洋潮汐周期性或间歇性影响, 覆盖有草本植物的咸水或咸淡水淤泥质滩涂[4]。潮沟广泛分布于盐沼生境, 并影响着盐沼和河流/海洋生态系统之间的物质联系[5]。糠虾、桡足类、蟹类幼体等大型浮游动物主要通过潮沟系统进入盐沼, 它们一方面摄食盐沼中的碎屑等食物, 另一方面被进入盐沼的鱼类等游泳动物所食[6]。此外, 大型浮游动物的捕食对中小型浮游动物的生物量具有明显调控作用[7, 8]。因此, 大型浮游动物的种群动态变化对盐沼生态系统的结构、物质能量输移、渔业资源等均具有重要的影响[9, 10]。
我国是西北太平洋地区海岸线最长的国家, 分布着大面积的盐沼湿地, 其中长江口盐沼湿地面积约为3000 km2[11]。已有报道指出, 长江口盐沼湿地有大量浮游动物随潮汐进出, 数量可高达226.13×103ind/m3[12—14]。但这些已有的研究主要聚焦中小型浮游动物类群(体长0.06—1 mm), 虽然也涉及糠虾和仔稚鱼等少量较大体型的浮游动物类群, 但尚未有关于大型浮游动物群落结构和分布的专门报道。
本研究以长江口崇明东滩盐沼为研究地点, 选择两条不同类型潮沟开展大型浮游动物物种组成和时空分布的初步研究。主要回答以下三个问题: (1)崇明东滩盐沼潮沟大型浮游动物由哪些种类组成?(2)大型浮游动物群落在不同盐度的潮沟间是否有区别?在不同季节之间、日夜潮之间是否有差异?(3)哪些潮沟水体理化因子对大型浮游动物的分布具有主要影响?
1 地点、材料与方法
1.1 研究地点
崇明东滩盐沼湿地(121°50'—122°05'E, 31°25'—31°38'N) 位于长江河口, (图1), 年平均气温15.3℃, 年平均降水量1117.1 mm[15], 属于中等强度潮汐海区, 平均潮差2.7 m, 潮差范围2.45—4.96 m; 潮汐属非正规浅海半日潮, 一昼夜有两次高潮和两次低潮的变化, 平均高潮位为3.29 m, 平均低潮位为0.6 m[13, 15]。在潮汐动力作用下, 崇明东滩形成了20多条发育完善的潮沟系统[16]。本研究选择2条发育良好的潮间带潮沟——团结沙潮沟和小南港潮沟进行采样(图1)。团结沙潮沟处于冲刷岸段, 年平均侵蚀速率为 20.77 m/a, 底质平均粒径为22.65 μm (布乃顺未发表数据), 盐度相对较低; 小南港潮沟处于淤涨岸段, 年平均淤涨速率为 126. 85 m/a, 底质平均粒径为6.28 μm, 盐度相对较高[17]。

图1 崇明东滩盐沼潮沟大型浮游动物采样点及采样网具设置示意图
1.2 采样方案
2010年5月14日和10月8日, 新月大潮第一天, 分别在团结沙潮沟和小南港潮沟下游距离潮沟口约500 m处, 于潮沟中架设2顶插网(网口0.5 m×0.5 m, 网目0.5 mm× 0.5 mm)作为2个重复, 分别采集一次日潮和一次夜潮退潮过程中的大型浮游动物, 共计16个样品。采样期间, 日潮和夜潮的时间大致相当, 均为9—10h。插网网口中央距离潮沟底部0.5 m, 插网前方3 m处架设一面挡网, 防止大块碎屑进入网袋, 挡网尺寸为5 m×2 m, 网目4 cm× 4 cm。设置插网时, 在网口处固定一个1000 mL广口塑料标本瓶, 收集水样测定水温、盐度、pH、溶氧和浊度等水体理化因子。
1.3 样品处理
大型浮游动物样品现场用10%甲醛海水固定保存, 带回实验室挑选、鉴定、分类计数。挑拣时手工去除体长大于1 cm的浮游动物个体, 使用0.6 mm孔径的不锈钢筛网过滤去除样品中的中小型浮游动物。大型浮游动物个体数统计采用分亚样的方法, 抽样时保证每个亚样中大型浮游动物个体数不少于200个。若个体数少于1000个, 则计数样品中的全部大型浮游动物个体, 并进行物种鉴定, 尽可能地鉴定到最低分类级别。
1.4 数据分析
使用STATISTICA 6.0软件, 运用两因子方差分析检验不同月份间和两条潮沟间的水体理化因子、大型浮游动物个体数、分类群数的差异, 采用最小显著差异法(LSD)进行多重比较,<0.05为差异显著。大型浮游动物群落相似性分析(ANOSIM)使用软件Primer 5.0, 使用软件Canoco for Windows 4.5进行群落和环境因子典型相关分析(CCA)。分析前对大型浮游动物个体数进行log (+1)转换。
2 结果
2.1 大型浮游动物种类组成
2010年5月和10月, 共采集到大型浮游动物3819.89×103个, 包括 3 个动物门, 28 个分类群(表1)。节肢动物门端足目和十足目各有6 个分类群, 是分类群数最多的 2 个目。中华拟亮钩虾Ren以及仿美钩虾科Paracalliopiidae和巨颚水虱科Gnathiidae的种类为长江口首次发现。在个体数量上占优势的是哲水蚤目、十足目和糠虾目, 分别占总捕获数量的47.94%、44.08%和6.17%。此外, 仔稚鱼、等足目、端足目和沙蚕目的捕获数量也较多, 个体数均在10000 个以上。在能鉴定到物种水平的种类中, 采样捕获个体数超过4×103个的有短额刺糠虾Chen et Wang、雷伊著名团水虱HoestlandtJo、日本刺沙蚕Izuka、尖叶大狐钩虾、日本旋卷蜾蠃蜚Hirayama和圆锯齿吻沙蚕Hartman。
2.2 大型浮游动物的时空分布
5月份进入盐沼潮沟的大型浮游动物个体数量显著高于10月(图2、表2), 大型浮游动物最高捕获量出现在5月的小南港潮沟, 为1487.44×103个/(网·日)。小南港潮沟大型浮游动物总个体数和分类群数均显著高于团结沙潮沟, 大型浮游动物个体数量为团结沙潮沟中的10.25倍(5月)和10.68倍(10月)。

表1 崇明东滩盐沼潮沟大型浮游动物名录及缩写
注: 捕获总个体数为1—500个用“+”表示, 501—5000个用“++”表示, 5000个以上用“+++”表示
Note: “+” represents the total individual numbers between 1—500; “++” represents the total individual numbers between 500—5000; “+++” represents the total individual numbers above 5000
除仔稚鱼和等足目外, 其他5个优势大型浮游动物类群(哲水蚤目、十足目、糠虾目、端足目和沙蚕目动物)在小南港潮沟中的个体数量均显著高于团结沙潮沟。除端足类个体数量在两个月份无显著差异外, 5月份哲水蚤目、十足目、沙蚕目动物和仔稚鱼的个体数量显著多于10月份, 而糠虾和等足类正相反(表2)。
除日本刺沙蚕个体数量在两个月份间无显著差异外, 5月份日本旋卷蜾蠃蜚和圆锯齿吻沙蚕的个体数量显著高于10月份, 而短额刺糠虾、雷伊著名团水虱和尖叶大狐钩虾则正相反(表2)。除雷伊著名团水虱和圆锯齿吻沙蚕外, 其他4个优势物种(短额刺糠虾、日本刺沙蚕、尖叶大狐钩虾、日本旋卷蜾蠃蜚)在小南港潮沟中的个体数量均显著高于团结沙潮沟。
从日潮和夜潮捕获的大型浮游动物分析, 除了仔稚鱼以外, 其他6个优势类群均表现为夜潮捕获数量多于日潮(表3)。5月份的十足目蟹类幼体主要是蚤状幼体形态, 最高捕获量出现在5月份的小南港潮沟, 单个夜潮的捕获数量可高达764.85×103个/网。10月份, 则以大眼幼体和幼蟹形态出现, 在小南港潮沟数量较多(表1)。
2.3 大型浮游动物群落结构
群落相似性分析(ANOSIM)显示大型浮游动物群落在春、秋季节间差异显著(= 0.931,= 0.001)。10月, 大型浮游动物群落结构在团结沙潮沟和小南港潮沟间的差异较大, 而5月份大型浮游动物群落结构在两条潮沟间的差异相对较小。
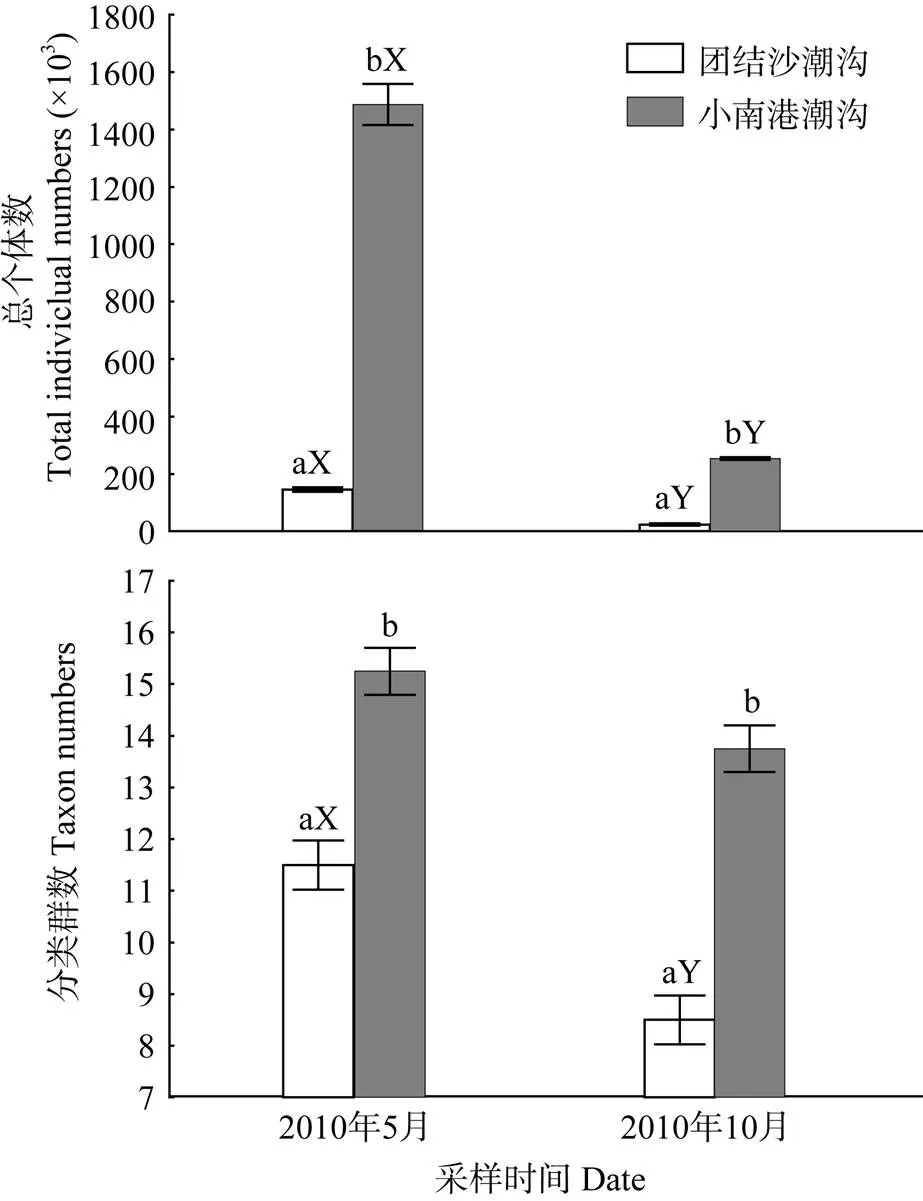
图2 崇明东滩盐沼潮沟大型浮游动物总个体数和分类群数
a, b表示同一月份在两条潮沟之间存在显著差异; X, Y表示同一条潮沟在不同月份之间存在显著差异
a and b indicate a significant difference between Tuanjiesha creek and Xiaonangang creek in the same month; X and Y indicate a significant difference between May and October in the same creek

表2 大型浮游动物总个体数、物种数、优势类群个体数和优势种类个体数双因子方差分析
注: 表示为值和值, 季节之间和采样潮沟之间差异显著时加粗表示
Note: Shown are-values with significance levels in parentheses, bold indicates significant difference

表3 大型浮游动物数量优势类群的个体数在日潮和夜潮间的差异
注: 个体数表示为×102个, 平均数±标准误; a, b表示在日潮和夜潮之间差异显著
Note: Individual numbers expressed as ×102mean numbers per net and SE. Superscripts a and b show a significant difference between day and night tides
典型相关分析(CCA)结果显示, 盐度、浊度、水温、pH和溶氧5个水体理化因子对大型浮游动物群落变化的总方差贡献率为97.8 %; 第一排序轴的特征值为0.217, 方差贡献率为47.5 %; 第二排序轴特征值为0.165, 方差贡献率为36.3 %。通过Monte Carlo检验发现, 盐度与大型浮游动物群落结构变化显著相关(=3.02,=0.014), 浊度对大型浮游动物群落结构有边缘显著影响(= 2.71,= 0.06)(图3)。

图3 大型浮游动物种类、采样点和潮沟水体理化性质典型对应分析
三角形、圆形和箭头分别表示大型浮游动物的种类、采样点和潮沟水体理化性质, 大型浮游动物的缩写见表2
Triangles, circles and arrows indicate macrozooplankton species, samples and water properties, respectively. For abbreviations of macrozooplankton, see Tab. 2
3 讨论
3.1 大型浮游动物种类组成
崇明东滩盐沼潮沟的大型浮游动物中, 哲水蚤目、十足目和糠虾目动物是数量优势类群, 分别占总捕获数量的47.94 %、44.08 %和6.17 %。Ceccherelli,.和Tan,.报道桡足类是河口中小型浮游动物中的优势类群[18, 19]。长江口水域浮游动物的研究[20, 21]及崇明东滩盐沼潮沟中小型浮游动物的研究[12, 14]结果表明桡足类是优势类群。而我们的研究表明, 在大型浮游动物这一生态类群中, 桡足类(主要是哲水蚤)在数量上仍然占很大比重。此外, 本研究显示十足目动物(主要是蟹类幼体)是大型浮游动物中的第二大数量优势类群。这反映了崇明东滩盐沼蟹类是植被区主要动物类群的特征, 同时也反映了盐沼与周边河口水体之间存在的重要物质联系[22]。
3.2 大型浮游动物群落在日夜潮和春、秋季节间的差异
本研究结果表明, 大型浮游动物个体数在昼夜潮汐间差异显著, 夜潮捕获的大型浮游动物个体数显著多于日潮(表3)。5月份夜潮捕获蟹类蚤状幼体数量可比日潮高 72.6 倍。这与盐沼蟹类主要在春季的夜间释放蚤状幼体有关[23]。10月份捕获蟹类幼体的形态为大眼幼体和幼蟹, 主要是从河口水体返回盐沼生境的个体[24], 而它们在日夜潮之间的数量没有显著差异。其他大型浮游动物类群(如哲水蚤目、等足目和沙蚕目等)也常出现夜潮的数量远超日潮的情况。团结沙和小南港潮沟夜潮的水温均高于日潮水温, 且差异显著, 因此温度可能是导致大型浮游动物数量日、夜间差异的原因之一。而大型浮游动物动物在日夜潮间的差异可能是导致夜潮进入盐沼捕食的鱼类数量多于日潮的原因之一[25]。
崇明东滩盐沼潮沟大型浮游动物群落结构在春、秋季之间差异显著, 而且5月份进入盐沼潮沟的大型浮游动物个体数量显著高于10月(图2、表2)。水体理化因子方差分析结果显示, 崇明东滩盐沼潮沟水体盐度在两个采样月份间差异显著, 水体温度的差异呈边缘显著。有报道指出盐度是影响浮游和底栖动物种类组成、多样性和分布格局的重要因素[26, 27]。而温度也会影响浮游动物种类和丰度的时空变化, 导致优势种的更替[12, 28]。因此, 盐度和水温可能是导致崇明东滩盐沼潮沟春、秋季间大型浮游动物群落差异显著的环境因素。
3.3 大型浮游动物群落的空间差异
水动力条件(如淹水时间、潮汐强度)和盐度是影响盐沼潮沟浮游动物群落空间分布的主要因子[20, 29, 30], 曾有研究报道水温和盐度影响着长江口水域浮游动物优势种类哲水蚤的空间分布[31], 另外李共国等近期研究报道淡水排放、潮沟径流和潮汐决定了滩涂湿地浮游动物群落的时空格局[32]。本研究发现小南港潮沟大型浮游动物总捕获个体数约是团结沙潮沟的10倍, 物种/分类群数也显著高于团结沙潮沟。崇明东滩团结沙潮沟与北港相通, 受长江径流影响大[33], 盐度明显低于小南港潮沟, 大型浮游动物群落表现为河口半咸水种类(脊尾白虾、仔稚鱼)相对较多, 并且有少量淡水种类(崇西水虱、枝角类和秀丽白虾)分布。小南港潮沟主要受海水及半咸水团的控制, 盐度相对较高, 其群落以河口半咸水种(日本刺沙蚕、尖叶大狐钩虾)为主, 淡水种很少出现。此外, 团结沙潮沟位于潮间带冲刷区, 潮滩沉积物颗粒较粗, 而小南港潮沟位于淤积区, 潮滩沉积物颗粒较细, 因此小南港潮沟水体的浊度明显高于团结沙潮沟[34]。除了盐度以外, 浊度也是影响浮游动物种类组成、多样性和分布的重要环境因子[26, 27, 35]。而本研究通过CCA分析结果也证实盐度和浊度是影响崇明东滩大型浮游动物群落结构空间差异的重要因子。总体而言, 大型浮游动物数量和多样性在盐度较高的淤积区潮沟较高, 这个研究结果也可能反映了盐沼与周边河口水体之间的物质交换在滩涂淤积区相对更为频繁。
[1] Wimpenny R S. The Plankton of the Sea [M]. New York: Elsevier. 1966, 21—36
[2] Azovsky A I, Saburova M A, Chertoprood E S. Selective feeding of littoral harpacticoids on diatom algae: hungry gourmands [J]?, 2005, 148: 327—337
[3] Porri F, McQuaid C D, Froneman W P. Spatio-temporal variability of small copepods (especially) in shallow nearshore water off the south coast of South Africa [J].,, 2007, 72: 711—720
[4] Nybakken J W, Bertness M D. Marine Biology [M]. San Francisco: Benjamin Cummings. 2005, 11—43
[5] Mallin M A, Lewitus A J. The importance of tidal creek ecosystems [J]., 2004, 298: 145—149
[6] Hampel H, Cattrijsse A. Temporal variation in feeding rhythms in a tidal marsh population of the common goby(Kroyer, 1838) [J]., 2004, 66(3): 315—326
[7] Hutchings L, Pillar S C, Verheye H M. Estimates of standing stock, production and consumption of mesozooplankton and macrozooplankton in the Benguela ecosystem [J]., 1991, 11: 499—512
[8] Pakhomov E A, Perissinotto R, Froneman P W. Predation impact of carnivorous macrozooplankton and micronekton in the Atlantic sector of the Southern Ocean [J]., 1999, 19(1—3): 47—64
[9] Vargas C A, Martinez R A, Escribano R,. Seasonal relative influence of food quantity, quality, and feeding behavior on zooplankton growth regulation in coastal food webs [J]., 2010, 90(6): 1189—1201
[10] Winder M, Jassby A D. Shifts in zooplankton community structure: Implications for food web processes in the upper San Francisco Estuary [J]., 2011, 34(4): 675—690
[11] Chen Y Q, Ye W J, Xu Z L. Yangtze Delta wetlands ecosystem characteristics and ecological restoration [A]. In: Shanghai wetland protection symposium [C]. Shanghai: Shanghai Science and Technology Press. 2003, 115—121 [陈亚瞿, 叶维均, 徐兆礼. 长江口滨海湿地生态特征及生态修复. 上海市湿地保护研讨会论文集. 上海: 上海科技出版社. 2003, 115—121]
[12] Zhou S C, Jin B S, Guo L,. Spatial distribution of zooplankton in the intertidal marsh creeks of the Yangtze River Estuary, China [J]., 2009, 85(3): 399—406
[13] Xu H F, Zhao Y L. Scientific survey on Chongming Dongtan migratory Birds Nature Reserve of Shanghai [C]. Beijing: China Forestry Publishing House. 2005, 2—24 [徐宏发, 赵云龙.上海市崇明东滩鸟类自然保护区科学考察集. 北京: 中国林业出版社. 2005, 2—24]
[14] Li Q, An C G, Xu L L,. Species composition and diversity of zooplankton in tidal creeks of the Chongming Dongtan intertidal flat [J]., 2010, 41: 214—222 [李强, 安传光, 徐霖林, 等.崇明东滩潮间带潮沟浮游动物的种类组成及多样性. 生物多样性, 2010, 41: 214—222]
[15] Chen J K. Scientific Investigations of the Shanghai Jiuduansha Wetland Nature Reserve [M]. Beijing: Science Press. 2003, 30—62 [陈家宽. 上海九段沙湿地自然保护区科学考察集. 北京: 科学出版社. 2003, 30—62]
[16] Xie D F, Fan D D, Gao S. Flat of Chongming Island and its impacts on the sediment distribution [J]., 2006, 26(2): 9—16 [谢东风, 范代读, 高抒. 崇明岛东滩潮沟体系及其沉积动力学. 海洋地质与第四纪底质, 2006, 26(2): 9—16]
[17] Han Z, Yun C X, Dai Z J. Remote sensing quantitative inversion research of the silt tidal flat elevation and deposition and erosion - Chongming Dongtan as the example [J]., 2009, 1: 12—18 [韩震, 恽才兴, 戴志军. 淤泥质潮滩高程及冲淤变化遥感定量反演方法研究——以长江口崇明东滩为例.海洋湖沼通报, 2009, (1): 12—18]
[18] Ceccherelli V U, Mistri M, Franzoi P. Predation impact on the meiobenthic harpacticoidin a lagoon of the Po River Delta, Italy [J]., 1994, 17(1): 283—287
[19] Tan Y H, Huang L M, Chen Q C,. Seasonal variation in zooplankton composition and grazing impact on phytoplankton standing stock in the Pearl River Estuary, China [J]., 2004, 24(16): 1949—1968
[20] Xu Z L. Character of zooplankton community and its variation in the water near the Yangtze River estuary [J]., 2005, 24(7): 780—784 [徐兆礼, 长江口邻近水域浮游动物群落特征及变动趋势. 生态学杂志, 2005, 24(7): 780—784]
[21] Guo P Y, Shen H T, Liu A C,. The species composition, community structure and diversity of zooplankton in Changjiang estuary [J]., 2003, 23(5): 893—900 [郭沛涌, 沈焕庭, 刘阿成, 等. 长江河口浮游动物的种类组成、群落结构及多样性. 生态学报, 2003, 23(5): 893—900]
[22] Qin H M, Chu T J, Xu W,. Effects of invasive cordgrass on crab distributions and diets in a Chinese salt marsh [J]., 2010, 415: 177—187
[23] Dittel A I, Epifanio C D, Lizano O. Flux of crab larvae in a mangrove creek in the Gulf of Nicoya, Costa Rica [J].,, 1991, 32: 129—140
[24] Johnson G E, Gonor J J. The tidal exchange of(Crustacea, Decapoda) Larvae between the ocean and the Salmon river estuary, Oregon [J]., 1982, 14: 501—516
[25] Jin B S, Fu C Z, Zhong J S,. Fish utilization of a salt marsh intertidal creek in the Yangtze River estuary, China [J].,, 2007, 73: 844—852
[26] Li K Z, Yin J Q, Huang L M,. Spatial and temporal variations of mesozooplankton in the Pearl River estuary, China [J].,, 2006, 67: 543—552
[27] Echaniz S A, Vignatti A M. Seasonal variation and influence of turbidity and salinity on the zooplankton of a saline lake in central Argentina [J]., 2011, 39(2): 306—315
[28] Fulton R S. Interactive effects of temperature and predation on an estuarine zooplankton community [J]., 1983. 72: 67—81
[29] Quintana X D. Measuring the intensity of disturbance in zooplankton communities of Mediterranean salt marshes using multivariate analysis [J]., 2002, 24: 255—265
[30] Badosa A, Boix D, Brucet S,. Nutrients and zooplankton composition and dynamics in relation to the hydrological pattern in a confined Mediterranean salt marsh (NE Iberian Peninsula) [J].,, 2006, 66: 513—522
[31] Xu Z L, Shen X Q, Ma S W. Ecological characters of zooplankton dominant species in the waters near the Changjiang estuary in spring and summer [J]., 2005, 29(12): 13—19 [徐兆礼, 沈新强, 马胜伟. 春、夏季长江口邻近水域浮游动物优势种的生态特征. 海洋科学, 2005, 29(12): 13—19]
[32] Li G G, Tu X X, Wang P E,. Influence of freshwater discharge on zooplankt community distribution in the mudflat wetlands of the Hangzhou Bay [J]., 2013, 37(3): 397—404 [李共国, 屠霄霞, 王佩儿. 等. 淡水排放对杭州湾湿地浮游动物群落分布的影响. 水生生物学报, 2013, 37(3): 397—404]
[33] Feng G P, Zhuang P, Liu J,. Community diversity and growth characteristic of fish in Tuanjiesha along the eastern beach of Chongming Island [J]., 2007, 29(1): 38—43 [冯广朋, 庄平, 刘健, 等. 祟明东滩团结沙鱼类群落多样性与生长特性. 海洋渔业, 2007, 29(1): 38—43]
[34] Liu Q Y, Dai X R, He X Q. The spatial distribution characteristics of sediment grain size on the east tidal flat, Chongming Island [J]., 2003, (4): 5—10 [刘清玉, 戴雪荣, 何小勤. 崇明东滩表层沉积物的粒度空间分布特征. 上海地质, 2003, (4): 5—10]
[35] Lin X, Li C Y, Lu K H. The effect of temperature and salinity on the survival of[J].(NSEE), 2001, 14(1): 43—46. [林霞, 李春月, 陆开宏. 温度和盐度对细巧华哲水蚤存活率的影响. 宁波大学学报(理工版), 2001, 14(1): 43—46]
Distribution of macrozooplankton community in Chongming Dongtan salt marsh creeks during summer and autumn
QIN Hai-Ming1, TANG Chen-Dong2, MA Qiang2, CHEN Jia-Kuan1, 3and WU Ji-Hua3
(1. Center of Watershed Ecology, Institute of Life Science, Nanchang University, Nanchang 330031, China; 2. Shanghai Chongming Dongtan National Nature Reserve, Shanghai 202183, China; 3. Ministry of Education Key Laboratory for Biodiversity Science and Ecological Engineering, Fudan University, Shanghai 200433, China)
Macrozooplankton; Taxon composition; Temporal-spatial distribution; Salt marsh; Creek; Yangtze River Estuary
2013-01-17;
2013-12-10
国家重点基础研究发展计划项目(2013CB430404); 上海市科学技术委员会(12231204700, 13231203503)资助
秦海明(1978—), 男, 山东昌乐人; 博士; 主要从事浮游动物生态与资源保护研究。E-mail: qinhaiming@ncu.edu.cn
吴纪华(1973—), 女, 教授; 主要从事海滨和河口湿地水生生物多样性、食物网动态研究。E-mail: jihuawu@fudan.edu.cn
Q178.1+4
A
1000-3207(2014)02-0375-07
10.7541/2014.53
猜你喜欢
小主人报(2022年18期)2022-11-17 02:19:52
煤气与热力(2022年4期)2022-05-23 12:44:56
天津化工(2021年3期)2021-06-11 08:03:12
山东煤炭科技(2020年4期)2020-05-12 05:34:28
环球时报(2019-11-14)2019-11-14 05:16:17
百科探秘·航空航天(2017年10期)2017-11-08 08:14:52
中国工程咨询(2017年4期)2017-01-31 03:05:28
中国工程咨询(2015年5期)2015-02-16 05:35:06
自然资源遥感(2014年3期)2014-02-27 11:56:42
中国摄影家(2013年8期)2013-04-29 00:44:03
