
草原锦鸡儿直播造林模式的比较研究
2014-04-03 02:00:40陈玉萍黄俊华胡成志姚雨仙
中国水土保持 2014年11期
陈玉萍,黄俊华,胡成志,姚雨仙
(1.新疆农业大学 林学与园艺学院,新疆 乌鲁木齐 830052;2.乌鲁木齐县水电林业局,新疆 乌鲁木齐县 830036)
锦鸡儿属植物主要分布于亚洲干旱和半干旱地区,集中分布在草原和荒漠区,中国是该属的主要分布区[1]。锦鸡儿属植物具有繁殖性强、抗逆性强等特点[2],其根系发达、具根瘤,能通过根瘤固氮改善土壤肥力[3],在荒漠、草原地区是水土保持、防风固沙、改善环境、荒山绿化、植被恢复的先锋树种和优质牧草资源。国内许多学者对其植物生理、种子萌发特性、无性繁殖等多个方面[4-5]进行了研究。
草原锦鸡儿(CaraganapumilaPojark.)隶属于锦鸡儿属,在新疆地区主要分布于阿尔泰山、天山、准噶儿西部山地,常形成地带性植被。在西北山地、丘陵地区及新疆荒漠草原地区,直播造林技术简单易行,对大面积缺水地区造林具有重要意义,但迄今为止,仅见李晓梅等对锦鸡儿属植物有关种子萌发进行了研究[6],而关于其直播造林的技术未见报道。其他植物直播技术的研究表明,种子前期处理[5]、播种模式处理[7]和播种量处理[8]均对直播出苗率有一定的影响。探讨草原锦鸡儿的直播模式,可为新疆干旱半干旱区大面积无水源的瘠薄地区应用直播造林技术提供依据,为草原锦鸡儿在植被恢复及荒地造林中的应用提供技术指导。
1 试验材料与方法
1.1 试验材料
草原锦鸡儿种子于2012年9月采自新疆乌鲁木齐县的板房沟(43°31′10″N、87°41′49″E,海拔1 350 m),采种区草原锦鸡儿平均高度为(150±17.32)cm,平均冠幅为(146.83±26.65)cm,种子千粒重为(14.06±0.23)g。种子采收后置于通风条件良好的室内风干,选取颗粒饱满、大小均一、未受昆虫和病菌侵害的成熟种子室温下保存。
1.2 试验地概况
试验地位于乌鲁木齐县(水西沟)花卉基地,地理位置87°29′39″E、43°27′20″N,海拔1 776 m,地处南山脚下,属于大陆性北温带温和干旱气候,光照充足,风力较大。春季干旱多风,夏季炎热、雨量集中,秋季凉爽短促,冬季寒冷而漫长。年降水量不多,而且分布不均,年降水量的70%~80%集中在5—8月。年均气温2.5 ℃,极端最高气温39.5 ℃,极端最低气温-43.2 ℃。试验区土质为普通种植土。
1.3 试验方法
1.3.1 播种试验设计
穴播,行距20 cm,穴距15 cm,覆土厚2 cm[9]。秋播试验于2012年10月下旬进行,种子不做处理;春播于2013年5月下旬完成,播前一周对试验地喷洒除草剂、除虫剂及整地,种子用始温为70 ℃的温水浸泡24 h后播种。定期测量记录试验区地温、气温及土壤湿度情况。
试验采用双因素的随机试验设计,一为每穴播种粒数,分为5、10、20粒(对照),二为播种模式,分为先覆膜后播种模式、先播种后覆膜模式、梁峁播种模式、直播模式(对照),分别标记为XFB、XBF、LMN、ZB。双因素交叉共得到 12个处理,春秋播共24个处理。小区排列采用二因素随机区组排列。
1.3.2 出苗率、保存率、死亡率测定
播种后每2 d观测1次出苗情况,出苗以幼苗出土0.5 mm计数,出苗终止调查时间以苗木连续5 d不萌发为止,死亡率从开始出苗统计至6月22日截止。计算公式为
出苗率=(田间出苗数/播种粒数)×100%
死亡率=(死亡苗总数/出苗总数)×100%
保存率=[(出苗总数-死亡苗总数)/出苗总数]×100%
1.4 统计分析
本试验数据和图利用Excel 2010进行初步整理、绘图, 应用SPSS17.0及DPS统计软件进行单因素和双因素方差分析检验。不同季节、模式之间采用多重比较法,表、图中所有数据均为3个重复指标的平均值。
2 结果与分析
2.1 不同播种处理对出苗率的影响
2.1.1 不同播种季节对出苗率的影响
从春播、秋播试验对比分析(表1)可以看出,春播、秋播对出苗率差异影响显著,春播出苗率普遍高于秋播,其出苗率均值高于秋播20百分点以上。春播最高出苗率为92.7%,最低出苗率为39.1%;而秋播最高出苗率仅为74.7%,最低出苗率是36.1%;春播近一半的处理出苗率高于80%,而秋播均低于80%。总体而言,春季是草原锦鸡儿适宜播种的季节。
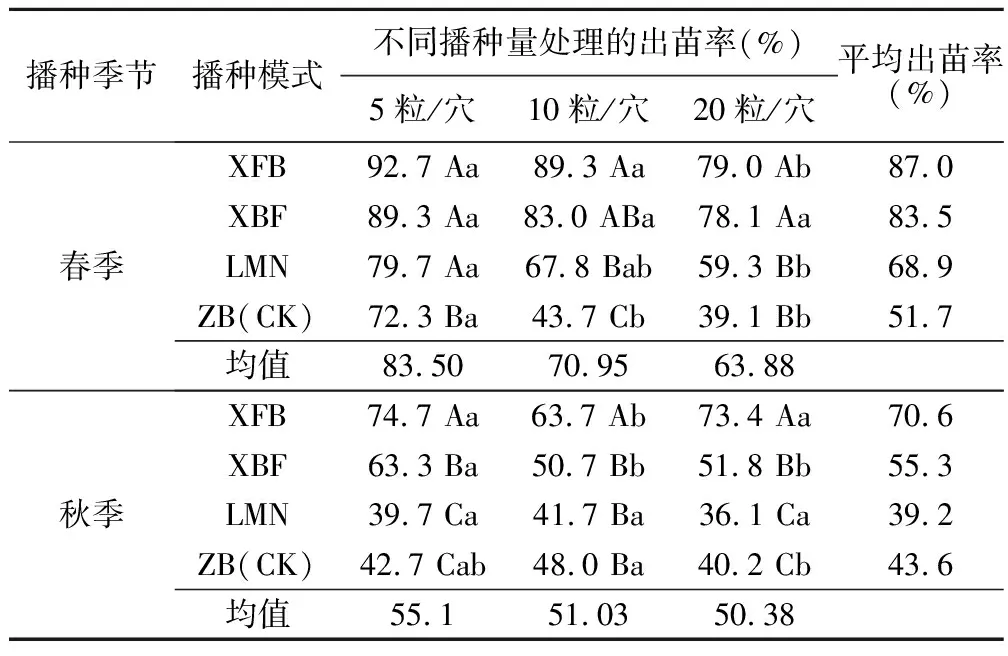
表1 春、秋季不同播种模式条件下草原锦鸡儿出苗率比较
2.1.2 播种模式和播种量对出苗率的影响
双因素方差分析结果显示(表2):春播条件下,播种量对出苗率的影响不显著(P>0.05),播种模式对出苗率的影响极显著(P<0.01),双因素交互作用对出苗率的影响显著(0.01 秋播条件下,仅播种模式对出苗率的影响极显著(P<0.01);播种量、播种模式和播种量的交互作用对出苗率的影响均不显著(P>0.05)。综上所述,春、秋季播种量对出苗率的影响均不显著,说明播种模式是影响种子出苗的重要因子。 表2 播种模式和播种量对出苗率影响的双因素方差分析 (1)不同播种量对出苗率的影响。在春播时节,XFB、LMN、ZB三种播种模式中5粒/穴播种量处理的出苗率显著高于20粒/穴,其中ZB播种模式中5粒/穴播种量处理与10、20粒/穴播种量处理均达到显著差异;XBF播种模式条件下各播种量处理出苗率差异均不显著(P>0.05)。春季播种条件下,出苗率随着每穴播种量的增加呈现降低趋势,各播种模式平均出苗率5粒/穴比10、20粒/穴分别高出12.55、19.62百分点。秋播条件下,仅在LMN播种模式下各播种量处理之间出苗率无显著性差异(P>0.05),XFB、XBF播种模式5粒/穴播种量的出苗率均高于其他播量,而ZB播种模式出苗率则表现为10粒/穴略高于5粒/穴、显著高于20粒/穴。各播种模式出苗率均值也表现为5粒/穴略高于其他播种量处理。综上所述,春、秋季草原锦鸡儿播种量选择5粒/穴最佳,既可保证出苗率又可以达到节约种子的目的。 (2)不同播种模式对出苗率的影响。由表1可知,春、秋季播种模式对出苗率的影响极显著。整体上,春、秋季各播种模式之间普遍表现为:在不同播量下,XFB播种模式出苗率显著大于对照的出苗率,XFB与XBF播种模式在春播时均无显著差异,而在秋播时差异均显著(P<0.05)。在各处理条件下,覆膜模式出苗率均大于未覆膜模式,并且同为覆膜模式,秋播20粒/穴XFB比XBF播种模式出苗率增长21.6百分点,先覆膜后播种更有利于出苗。综上所述,XFB播种模式下种子具有相对的出苗优势。因此,XFB播种模式为草原锦鸡儿播种最佳模式。 根据每日统计的幼苗存活率与时间关系绘图,见图1—6。对比春、秋播幼苗存活率与时间关系图可知,春播各模式出苗率高、出苗集中,且死亡率较低,因此幼苗存活率与时间关系图无明显波动,较稳定(图1、3、5);秋播于次年4月10日左右开始出苗,各播种模式出苗时间差异较大,出苗持续时间均较长,短期内幼苗出现严重死亡,导致幼苗存活率与时间关系图波动较大(图2、4、6)。 图1 春播5粒/穴幼苗存活率随时间的变化 图2 秋播5粒/穴幼苗存活率随时间的变化 图3 春播10粒/穴幼苗存活率随时间的变化 图4 秋播10粒/穴幼苗存活率随时间的变化 不同播种模式对幼苗存活率与时间关系的影响差异非常明显。春播XFB和XBF播种模式表现为出苗早、出苗速度快的特点,出苗较为集中,播种后5 d达到出苗高峰期,10 d左右出苗率达到最大值,并且出苗要比其他播种模式早5 d左右,死亡率也较低,分别为2%~3%和4%~8%,因此幼苗存活率较高;LMN播种模式出苗慢但稳定,累计出苗率较高,集中出苗率较低,无明显的出苗高峰期,出苗最为松散,出苗时间持续较长,死亡较少且死亡率持续平稳,一般稳定在10%以内,因此幼苗存活率与时间关系图波动变化不大;ZB播种模式出苗较晚,且出苗不集中,出苗时间持续25 d左右,出苗率也较低,死亡率高达16%~33%,因此幼苗存活率与时间关系图波动变化明显。秋播XBF与XFB播种模式受早春虫害和低温影响,幼苗死亡严重,幼苗存活率与时间关系图前期波动较大,后期较为平稳;而ZB和LMN模式出苗相对较晚,出苗率低,短期内出苗过程中有死亡现象,且死亡率高达80%左右,导致幼苗存活率与时间关系图稍有起伏波动。各播种模式相比而言,覆膜模式下草原锦鸡儿种子由于出苗率较高且出苗集中、存活率较高,在持续阴雨天种子萌发受水分胁迫和天气影响不明显,因此幼苗存活率与时间关系图波动较小,而其他模式受气候条件的影响极为明显。 图5 春播20粒/穴幼苗存活率随时间的变化 图6 秋播20粒/穴幼苗存活率随时间的变化 春秋播不同处理幼苗保存率见表3。由表3可知,春播各播种模式的保存率均在77%以上,均值为91.58%;而秋播各播种模式保存率不足20%,均值为11.53%,XBF模式幼苗全部死亡。而且,春播出苗率与保存率差异较小,大部分差异在20百分点以内;秋播出苗率与保存率差异较大,近一半差异在50百分点以上。因此,春播各处理模式保存率明显优于秋播各处理模式,尤其是春播XFB模式可以大幅度提高幼苗的保存率。春季采用XFB模式5粒/穴处理,可使草原锦鸡儿种子出苗率达92.7%,幼苗保存率达97%,在大面积直播绿化方面相对比较适宜。 表3 不同处理幼苗保存率比较 不同的播种期不但影响植物的生长发育,而且影响种子的出苗率[8]。草原锦鸡儿春播出苗率明显高于秋播,这与河北地区臭椿直播造林的研究结果[10]一致,而与山西臭椿的直播造林结果[11]相反,究其原因主要是研究地点的气候差异造成的。山西雨量充足,秋播种子在冬季层积催芽下吸足了水分,翌年春天随着气温升高,种子自然萌发,出苗率较高,而春播由于春旱水分严重不足,整地时土壤水分散失影响种子的萌发;河北地区与本试验区气候条件相似,冬季雨雪多,春季随气温升高冰雪融化,为春季播种提供了良好的水分条件,因此春播出苗率较高。此外,该研究区春播各处理模式出苗率及保存率比秋播高,还可能因为:①秋播经覆膜处理翌年早春膜内升温快,出苗早,但试验区昼夜温度不稳定,夜间温度可降至0 ℃以下,可能导致幼苗细胞内外结冰,对膜系统造成机械损伤[12],使早期幼苗出现冻害致死[13]。②秋播覆膜模式种子出苗较早,试验区早春拟步甲虫害较为严重,5月为发生高峰期,覆膜后地温升高,地温变化幅度减小[14],适宜的温度容易吸引虫子集聚,导致幼苗被食。③早春试验区雪、雨水较多,温度变化大,秋播的一部分种子长期在此环境下导致变质、发霉并彻底失去萌发能力,即使出苗,也会因水分过多而致幼苗受胁迫死亡。对柠条锦鸡儿的研究也表明过多的水分会强烈地抑制柠条种子在极高或极低的温度下发芽[15]。而春播出苗晚,避开了这种种不利影响,且播种前种子经温水处理后提高了萌发率和集中萌发优势,为种子萌发及保存提供了可靠保障。 研究结果显示,5粒/穴播种量出苗率及保存率均高于其他处理。随着播量的增加,出苗率和保存率反而降低,这与刘功朋等[8]研究早稻的结果一致。这主要与幼苗出苗期间相互竞争空间和光照及养分有关。随着播量的增加,种子萌发破土能力增强,大量萌发的种子集中破土时易使将要开始萌发的部分种子裸露,芽期易出现“闪芽”现象,随着每穴播种量的增加,芽期出现“闪芽”现象的概率变大,导致出苗率降低。柠条高密种植下也会由于对水分条件的竞争使苗木生长不良,甚至死亡[16]。适宜的播量有利于提高幼苗保存率。 种子萌发是受多种内、外因素影响的复杂过程。水分是种子萌发的重要影响因素之一,草原锦鸡儿种子在土壤水分含量3.75%~17.50%时萌发率最高[6]。覆膜可以有效地抑制水分的蒸发,增强土壤的蓄水和保水能力,使土壤湿度恒定在8.7%~16.8%之间,为种子萌发提供良好的水分条件。温度也是种子萌发的必要条件,低温会抑制种子生理活动,从而影响种子萌发。采取覆膜措施可以形成微环境,产生温室效应提高地温[14],使地温迅速达到种子的最佳萌发温度[4],通过改善温度条件促进种子与地面的水热循环,提高种子萌发能力。研究认为,覆膜模式出苗率一般优于未覆膜模式。这与小叶锦鸡儿[17]、皂荚[7]的研究结果相似。 与直播出苗率相比,提高种子保存率是直播造林技术的又一重点难题。春、秋播覆膜模式的保存率差异显著,可能是受春、秋播出苗时间差异的影响。秋播覆膜模式早春出苗早、膜下温度高、虫害严重;另外,秋播覆膜后,翌年早春杂草在高温环境下先出苗,且无法锄草,与幼苗竞争资源,从而降低了覆膜模式的幼苗成活率。而春播播种晚,避开了这些不利影响。宋彬[4]的研究表明,未萌发的草原锦鸡儿种子在萌发策略上表现出在适宜水分条件下可再次萌发,秋播中,梁峁播种模式体现了这种萌发策略的优势,而直播(ZB)模式早春出苗则会遇到雨水腐烂种子现象,降低出苗率。 在该试验地区,草原锦鸡儿不适宜秋播,春播覆膜模式可保证较高的出苗率和保存率。在实际生产中,应结合具体的自然地理条件采取适当的播种方式。在降水量大的地区,可采用梁峁播种模式,降低造林覆膜成本;在干旱寒冷地区,可采用春播覆膜播种模式。该研究结论可为今后草原锦鸡儿大面积推广应用提供参考依据,但在研究中发现草原锦鸡儿幼苗及成苗期均出现根腐病,这一问题尚需要进一步研究。 [参考文献] [1] 中国科学院中国植物志编辑委员会.中国植物志:第42卷,第1分册[M].北京:科学出版社,1993. [2] 安韶山,黄懿梅.黄土丘陵区柠条林改良土壤作用的研究[J].林业科学,2006,42(1):70-74. [3] Moukoumi J,Farrell R E,Van Rees K J C,et al.IntercroppingCaraganaarborescenswithSalixmiyabeanato satisfy nitrogen demand and maximize growth[J].BioEnergy Research,2012,5(3):719-732. [4] 宋彬,朱小虎,韩娜,等.温度及盐胁迫对新疆两种锦鸡儿种子萌发的影响[J].新疆农业大学学报,2011,34(2):119-124. [5] 苗昊翠,刘博,邓江宇,等.新疆两种锦鸡儿种子萌发特性研究[J].新疆农业科学,2009,46(4):810-817. [6] 李晓梅,赵晓英,赵法,等.新疆3种锦鸡儿属植物种子萌发对温度和土壤水分的响应[J].草业科学,2009,26(10):141-145. [7] 李庆梅,刘艳,侯龙鱼,等,几种处理方式对皂荚直播造林地微环境和出苗率的影响[J].林业科学研究,2009,22(6):851-854. [8] 刘功朋,陈恺林,张玉烛,等.播期与播量对直播早稻生长特性及产量的影响[J].湖南农业科学,2013,21(1):13-16. [9] Gutteman Y.Seed Germination in Desert Plants[M].Berlin:Springer,1993:145-230. [10] 李士民.丘陵山地臭椿直播造林试验[J].河北林业科技,2002,12(6):11-13. [11] 陈家锋.臭椿直播造林成活率初探[J].山西农业大学学报,2002(2):151-153. [12] 李先文,黄新华,刘京,等.植物冻害发生的机制及预防措施[J].湖北农业科学,2010,49(10):2579-2581. [13] Burke M J,Gusta L V,Quamme H A,et al.Freezing and injury in plants[J].Annual Review of Plant Physiology,1976,27:507-528. [14] 王卫华,王全九,刘建军.南疆棉花苗期覆膜地温变化分析[J].干旱地区农业研究,2011,29(1):139-145. [15] Zheng Yuanrun,Xie Zhixiao,Gao Yong,et al.Germination responses ofCaraganakorshinskiiKom. to light,temperature and waters tress[J].Ecological Research,2004,19(5):553-558. [16] Jia Haikun,Liu Yinghui,Xu Xia,et al.Simulation of soil water dynamics in aCaraganaintermediawoodland in Huangfuchuan watershed[J].Frontiers of Forestry in China,2007,2(2):119-127. [17] 房绍艳.小叶锦鸡儿高垄整地播种造林技术初探[J].内蒙古林业,2008(1):20.
2.2 不同播种处理下幼苗存活率与时间的关系
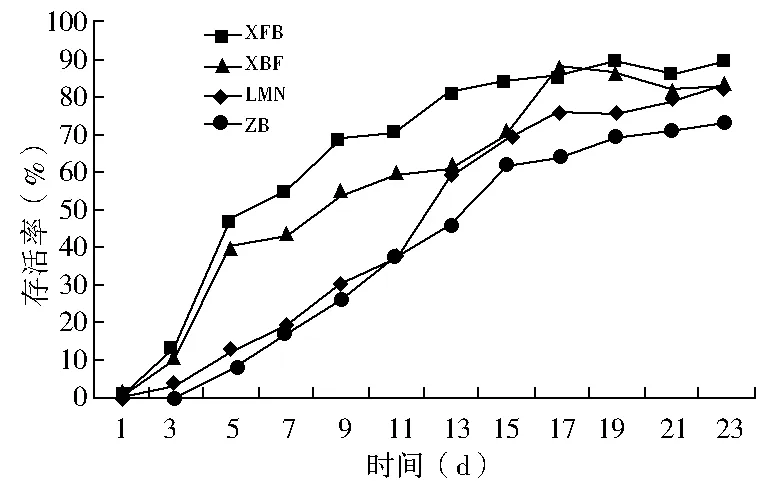
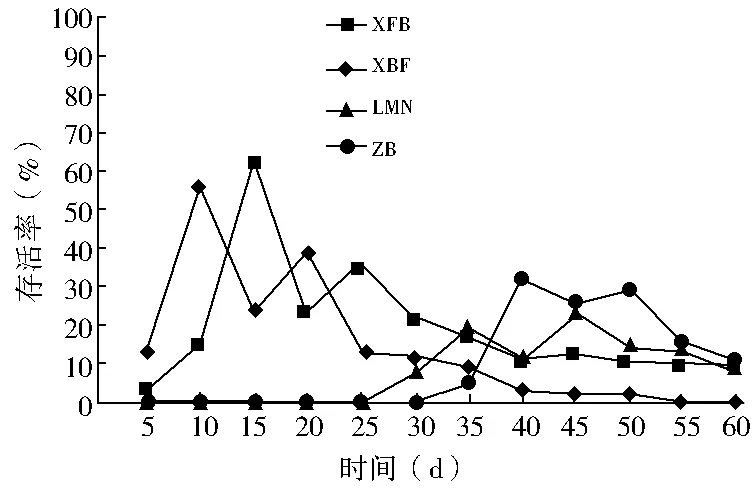
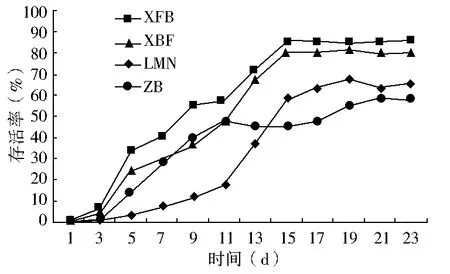
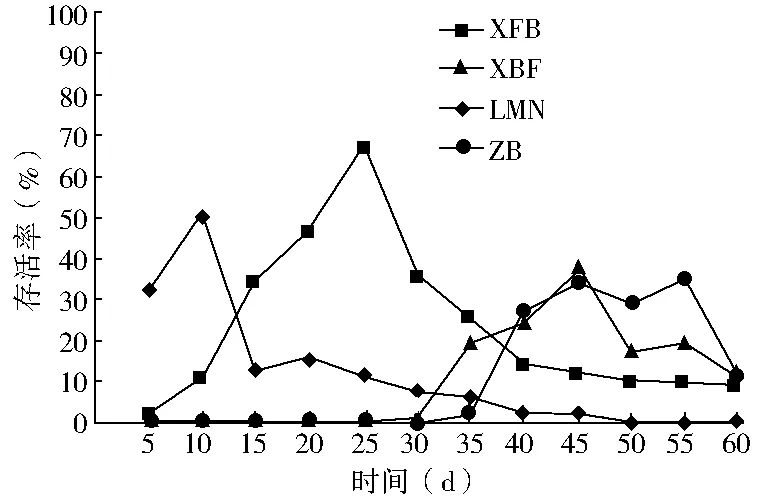
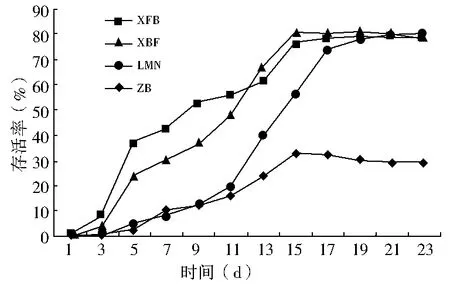
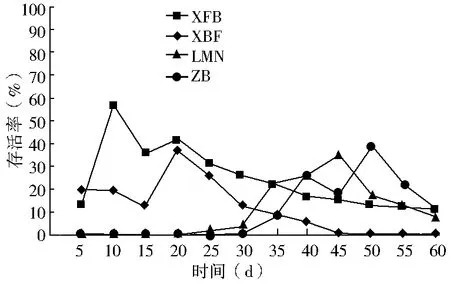
2.3 不同播种处理对幼苗保存率的影响

3 讨 论
3.1 春播、秋播对比
3.2 播种量对出苗率的影响
3.3 播种模式处理比较
4 结 语
猜你喜欢
世界热带农业信息(2022年11期)2022-10-18 10:20:12
吉林林业科技(2020年6期)2020-12-16 10:09:36
防护林科技(2020年5期)2020-07-21 01:53:32
水土保持研究(2020年1期)2020-04-27 08:06:22
烟台果树(2019年1期)2019-01-28 09:34:52
西藏农业科技(2018年4期)2018-04-25 06:39:24
现代农业科技(2017年22期)2018-01-08 12:58:41
党员生活(2016年11期)2016-11-15 18:58:59
山西林业科技(2016年3期)2016-11-12 03:44:26
食品与机械(2012年2期)2012-12-28 00:47:12
