
荔枝蒂蛀虫成虫羽化和交尾习性及性信息素释放节律的研究
2014-03-26 07:39:38陈晓琴江世宏
西北农林科技大学学报(自然科学版) 2014年10期
张 辉,陈晓琴,江世宏
(1 深圳职业技术学院 应用化学与生物技术学院,广东 深圳 518055;2 福建省农业科学院茶叶研究所,福建 福安 355015)
荔枝蒂蛀虫(ConopomorphasinensisBradley)属鳞翅目(Lepidoptera)细蛾科(Gracilariidae),主要分布于东南亚及我国广东、广西、海南、福建、香港、台湾等地[1],该虫为寡食性害虫,以幼虫钻蛀为害荔枝和龙眼的新梢、花穗和果实,荔枝常年被害率一般为20%~40%,高的达90%以上,严重影响了荔枝的产量和品质[2]。如广西报道荔枝被蛀果率达40%~60%,有些果园则高达60%~70%[3]。目前生物防治和耕作防治荔枝蒂蛀虫的效果十分有限,化学防治仍为最主要的防治方法,但该法容易导致害虫产生抗药性和果实农药残留,对出口创汇和农民增收造成一定影响。应用昆虫性信息素防治害虫是一种简便易行、高效无毒、无残留的无公害防治技术,已应用于多种鳞翅目害虫的防治实践[4-5]。而对于荔枝蒂蛀虫性信息素的研究较少,过去曾将在台湾为害荔枝和龙眼的荔枝蒂蛀虫与分布在东南亚的可可细蛾(C.cramerella)混为一个种,且鉴定出了可可细蛾的性信息素组分[6],但此信息素对荔枝蒂蛀虫的诱虫效果不佳[7],因为这2种细蛾确系不同的种类[8]。为此,本试验在观察荔枝蒂蛀虫成虫羽化和交尾习性的基础上,研究了雌蛾性信息素粗提物的生物活性和释放节律,期望为下一步性信息素组分的提取、分离、鉴定和性诱剂的研制提供依据。
1 材料与方法
1.1 供试昆虫
在深圳西丽果场捡拾荔枝落果,获取荔枝蒂蛀虫幼虫,放入养虫室人工气候箱中(温度(26±1) ℃,相对湿度(80±5)%,光照14L∶10D)饲养。成虫羽化后区分雌雄,分别放入不同的养虫笼内备用。
1.2 荔枝蒂蛀虫成虫的羽化动态观察
取1日龄荔枝蒂蛀虫蛹800头,置于养虫笼并放入上述人工气候箱中,每天观察1次成虫羽化情况。
1.3 荔枝蒂蛀虫成虫的求偶交尾规律观察
取10对同日龄(1,2,3,4,5日龄)的荔枝蒂蛀虫雌雄成虫,放入50 mL锥形瓶中,用透明纱布封口。在蒙上红布的台灯下每30 min观察1次求偶交尾情况。每处理重复3次。
1.4 荔枝蒂蛀虫雌蛾性信息素的提取
1.4.1 不同日龄雌蛾性信息素的提取 选取不同日龄的处女雌蛾各100头,进入暗期5 h后分别摘取腺体,放入5 mL具塞玻璃刻度试管内,注入1 mL冷正己烷,浸泡12 h后将上清液转移至1.5 mL的进样瓶中,然后将性腺粗提物置于-20 ℃冰箱内贮存备用。
1.4.2 不同暗期雌蛾性信息素的提取 选取3日龄的处女雌蛾,在交尾盛期的晚上21:30-01:30,每隔 1 h取100头处女雌蛾摘取腺体,提取性信息素,方法与1.4.1相同。
1.5 雄蛾对雌蛾性腺粗提物的反应
1.5.1 雄蛾触角电位反应测定 触角电位(Electroantenogram,EAG)反应测定在华南农业大学资源环境学院生态实验室进行。触角电位仪工作条件:连续气体流量为450 mL/min,刺激气体流量为40 mL/min;电位工作条件:直流,增益为200 V,测定电压范围为1 mV。调试触角电位仪至稳态后,将3日龄雄蛾的触角沿基部剪下,剪去触角两端,通过导电胶连接在电极上,调节仪器工作状态,待基线平稳后,进行触角电位仪测定。将直径10 cm的滤纸剪成纸条(0.5 cm×6 cm),滴加雌蛾性腺粗提物10 μL,放入巴斯管中,接入刺激气流管,踩脚踏板开始测定。测定时间从每晚的21:30开始,其中不同日龄雌蛾测定3个粗提物样品,不同暗期3日龄雌蛾测定5个粗提物样品,每个样品重复测定10次,结果取平均值,除去正己烷溶液对照。
1.5.2 雄蛾选择趋性反应测定 采用Y型嗅觉仪测定未交尾雄蛾对雌蛾性腺粗提物的选择趋性反应。测定方法如下:首先将雌蛾性腺粗提取物样品100 μL滴加在滤纸条(6 cm×2 cm)上,放入一个味源瓶中,另一味源瓶放入滴加100 μL正己烷的滤纸条作为对照,将2个味源瓶分别与Y型管的2个侧臂连接。然后将未交尾3日龄雄蛾1头放入Y型管主臂中,并开始记录成虫选择反应。若雄蛾飞至超过某臂2 cm,并持续1 min以上,则记录为做出了选择;若雄蛾20 min后没有做出选择,记为无反应,结束对该雄蛾的观察,未反应雄蛾的数据不计入统计数据。选择3日龄雌蛾不同暗期(22:30、23:30、00:30)的性腺粗提物进行测定,共3个处理,每个处理测定50头雄蛾,每测定5头后,调换两侧臂的位置,以消除管壁位置对试虫选择行为产生的影响;每测定10头后,用无水乙醇清洗Y型管并吹干备用,所有测定均从晚上22:00开始,测定数据用SPSS13.0统计软件进行χ2检验。
2 结果与分析
2.1 荔枝蒂蛀虫成虫的羽化动态
逐日观察荔枝蒂蛀虫成虫的羽化情况,结果见图1。

图1 荔枝蒂蛀虫成虫的羽化动态
从图1可以看出,荔枝蒂蛀虫化蛹后第3 天开始羽化,7~8 d达羽化高峰,10 d后羽化结束,羽化持续期约为6 d。在羽化高峰期前,雄虫的羽化数量多于雌虫;而在高峰期后,雌虫的羽化数量多于雄虫,但雌雄比例总体接近1∶1。
2.2 荔枝蒂蛀虫成虫的求偶交尾规律
荔枝蒂蛀虫成虫的求偶与交尾在暗期进行。首先进入求偶召唤期,雌蛾触角不动,双翅微张呈三角形,产卵管伸出体外向下,腹部偶而抽动;雄蛾触角则竖立微动,后开始展翅围绕雌蛾飞舞。然后开始交尾,雄蛾腹部交尾器下弯与雌蛾接合,雌雄并成“一”字型。交尾持续时间20~90 min,交尾过程中有时虫体可以走动。从不同日龄看,以3日龄成虫的交尾活动最为活跃,2、4日龄次之,1、5日龄较少(图2)。羽化5 d后未观察到成虫有交尾活动。从交尾时间看,在暗期的21:00-02:00均有成虫交尾,其中21:00后交尾率逐渐升高,至00:00达到交尾最高峰,此时交尾率达40%,之后下降;交尾高峰期发生在 22:00-01:00(图3)。
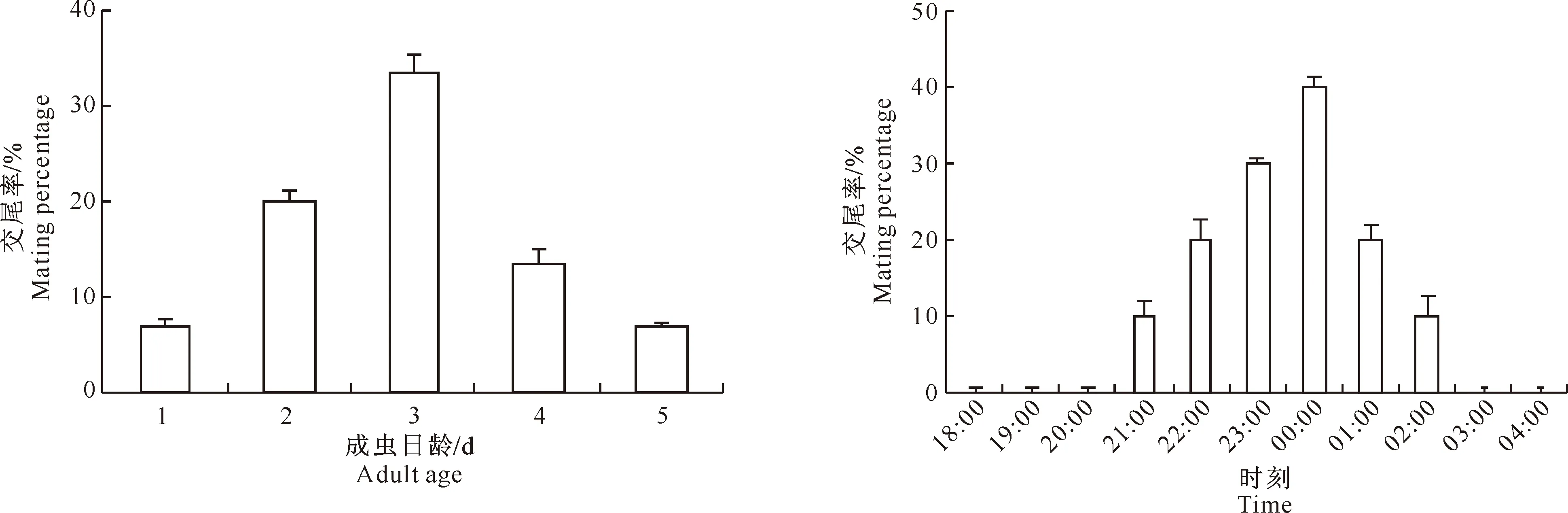
图2 不同日龄荔枝蒂蛀虫成虫的交尾规律
2.3 雄蛾对雌蛾性腺粗提物的EAG反应
2.3.1 对不同日龄雌蛾性腺粗提物的EAG反应 不同日龄荔枝蒂蛀虫雌蛾的性腺粗提物均能引起3日龄雄蛾的EAG反应(图4),且3日龄雄蛾对2、3、4日龄雌蛾性腺粗提物的EAG反应存在显著性差异(P<0.05)。其中3日龄雄蛾对3日龄雌蛾性腺粗提物的EAG反应值最大,其次是2日龄雌蛾粗提物,4日龄雌蛾粗提物反应值最小,表明雄蛾EAG反应值的大小与雌蛾的日龄密切相关,这与荔枝蒂蛀虫成虫的求偶交尾节律相一致,说明雌虫羽化后第3天是性信息素释放的高峰期,3日龄处女雌蛾性腺粗提物中的性信息素含量也最高,之后逐渐减少。
2.3.2 对3日龄雌蛾不同暗期性腺粗提物的EAG反应 3日龄雄蛾对3日龄处女雌蛾不同暗期性腺粗提物的EAG反应值如图5所示。由图5可以看出,3日龄雄蛾对3日龄雌蛾性腺粗提取物的EAG反应值从21:30开始逐渐增加,23:30达到最高峰,随后逐渐降低。统计分析表明,3日龄雄蛾对3日龄雌蛾不同暗期性腺粗提物的EAG反应存在明显差异,其中22:30-00:30的EAG绝对值显著高于其他时间(P<0.05),而21:30与01:30间无显著性差异。这与荔枝蒂蛀虫成虫的求偶交尾节律一致,说明3日龄雌蛾性信息素释放的高峰期为晚上22:30-00:30。
2.4 雄蛾对雌蛾性腺粗提物的选择趋性反应
利用Y型嗅觉仪测定荔枝蒂蛀虫雄蛾对雌蛾不同暗期性腺粗提物的选择趋性反应,结果(表1)表明,荔枝蒂蛀虫3日龄雄蛾对3日龄雌蛾 22:30-00:30的性腺粗提物具有明显的选择趋性,且各暗期差异显著,雄蛾对23:30雌蛾性腺粗提物的趋性与正己烷和22:30、00:30的粗提物存在显著差异。说明不同暗期荔枝蒂蛀虫雌蛾的性腺粗提物中均含有一定量的性信息素。
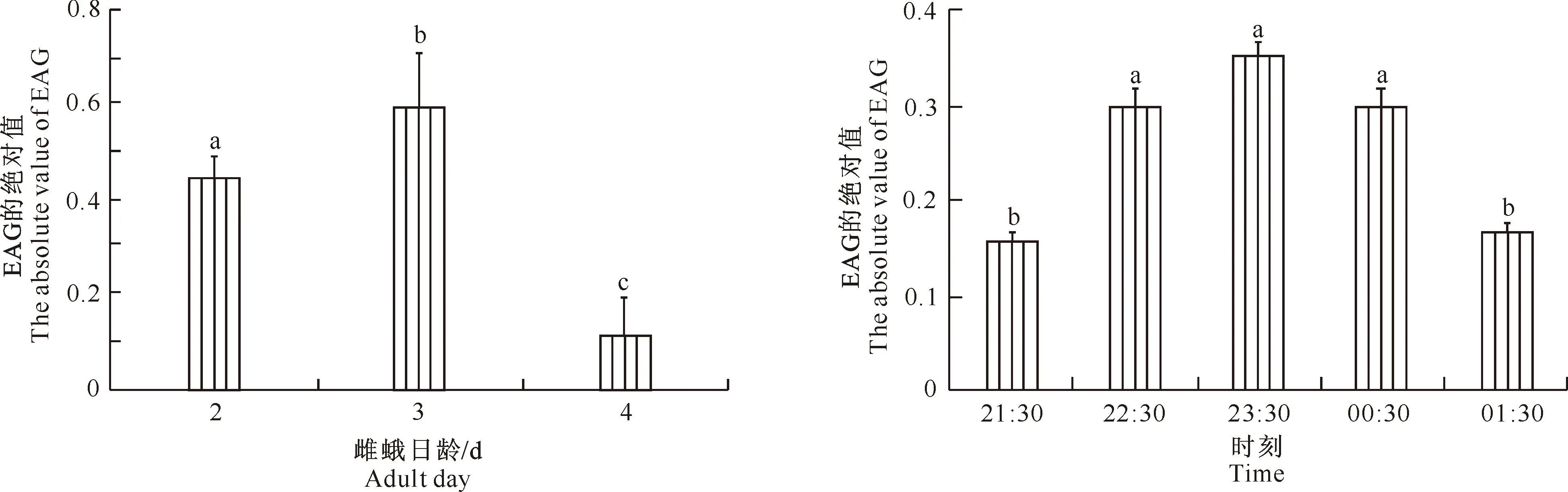
图4 3日龄雄蛾对不同日龄处女雌蛾性腺粗提物的EAG反应

表1 荔枝蒂蛀虫雄蛾对不同暗期3日龄雌蛾性腺粗提物的选择趋性反应
3 讨 论
本研究结果表明,荔枝蒂蛀虫成虫羽化高峰期在化蛹后7~8 d,这与姚监等[9]的研究结果一致。关于荔枝蒂蛀虫成虫的求偶交尾习性已有较多报道,从成虫日龄看,求偶交尾高峰均发生在2~3日龄的成虫;从求偶交尾时间看,也均在暗期进行,但不同文献报道的求偶交尾高峰期不同:洪巧珍等[10]报道在暗期的04:00-05:00,冯莉等[11]报道在凌晨的04:00-06:00,江世宏等[12]报道在21:00-22:00;本研究结果为22:00-01:00,与江世宏等的研究结果部分吻合。这些差异可能是因为饲养条件的不同所致,其他原因有待进一步探明。
本研究结果表明,荔枝蒂蛀虫的求偶交尾节律与雄蛾对雌蛾性腺粗提物的电生理和行为反应存在一致性。如求偶交尾高峰期为3日龄成虫且在晚上22:00-01:00,而雄蛾对3日龄处女雌蛾22:30-00:30性腺粗提物的EAG反应最强,3日龄雄蛾对3日龄雌蛾22:30-00:30性腺粗提物也有明显趋性,两者的高峰期基本吻合,说明荔枝蒂蛀虫的求偶交尾行为与性信息素的释放也存在明显的暗期节律,这种现象在小地老虎(AgrotisypsilonRottemberg)[13]、甜菜夜蛾(Spodopterqaexigua)[14]、豆野螟(Marucavitrata(Fabricius))[15]、榆木蠹蛾(Holcoerusvicarious(walker))[16]等其他蛾类昆虫中也比较常见。
昆虫性信息素因具有高效、专一、无毒、操作简单、效果持久等特点,受到国内外学者的重视,成为害虫无公害治理的重要措施之一。研究荔枝蒂蛀虫的羽化交配和性信息素的释放节律是利用性信息素防治害虫研究中的重要内容,也是其繁殖过程性行为反应研究的重要环节。本试验结果为下一步的理论与应用研究奠定了坚实的基础。
[参考文献]
[1] 姚振威,刘秀琼.危害荔枝和龙眼的两种细蛾科昆虫 [J].昆虫学报,1990,33(2):207-212.
Yao Z W,Liu X Q.Gracillariid insect pest attacking litchi and longyan [J].Acta Entomologica Sinica,1990,33(2):207-212.(in Chinese)
[2] 陈加福.中国有害生物综合治理文集 [M].北京:中国农业科技出版社,1996:839.
Chen J F.Proceedings of the national symposium on IPM in China [M].Beijing:China Agricultural Technology Press,1996:839.(in Chinese)
[3] 陈献萍.荔枝蒂蛀虫发生为害特性及防治关键技术研究 [D].广西南宁:广西大学,2006.
Chen X P.Studies on the occurrence ofConopomorphasinensisBradley and its key control techniques [D].Nanning,Guangxi:Guangxi University,2006.(in Chinese)
[4] 葛绍奎,李典谟,谢宝瑜,等. 棉铃虫性诱剂对棉铃虫成虫行为的影响及田间控制作用分析 [J].应用生态学报,1997,8(3):291-294.
Ge S K,Li D M,Xie B Y,et al.Effect of sex pheromone on behaviors of adultHeliothisarmigeraand its field control [J].Chinese Journal of Applied Ecology,1997,8(3):291-294.(in Chinese)
[5] 王永模,戈 锋,刘向辉,等.应用性信息素迷向法防治茶毛虫的田间试验 [J].昆虫知识,2006,43(1):60-63.
Wang Y M,Ge F,Liu X H,et al.Field experiments for controling the tea tussock moth,Euproctispseudoconspersa,by mating disruption with sex pheromone [J].Chinese Bulletin of Entomology,2006,43(1):60-63.(in Chinese)
[6] Beevor P S,Cork A,Hall D R,et al.Components of female sex pheromone of cocoa pod borer mothConopomorphacramerella[J].Journal of Chemical Ecology,1986,12(1):1-23.
[7] 黄振声,洪巧珍,严耀平,等.荔枝细蛾(ConopomorphasinensisBradley)之性引诱剂及诱虫器 [J].植物保护学会会刊(台湾),1996,38(2):129-136.
Huang Z S,Hong Q Z,Yan Y P,et al.Sex attraction and trap with sex attraction of litchi fruit borer,ConopomorphasinensisBradley (Lepidoptera: Gracillariidae) [J].Plant Protection Bulletin Taipei,1996,38(2):129-136.(in Chinese)
[8] 洪巧珍,黄振声.台湾为害荔枝及龙眼之细蛾种类 [J].植物保护学会会刊(台湾),1996,38(1):75-78.
Hong Q Z,Huang Z S.Gracillariidae insect pest attacking litchi and longyan in Taiwan [J].Plant Protection Bulletin Taipei,1996,38(1):75-78.(in Chinese)
[9] 姚 监,苏 丽,蒙永军,等,荔枝蒂蛀虫羽化及交配行为研究 [J].安徽农业科学,2009,37(25):12046-12048.
Yao J,Su L,Meng Y J,et al.Study on the eclosion and mating behaviors ofConopomorphasinensisBradley [J].Journal of Anhui Agricultural Sciences,2009,37(25):12046-12048.(in Chinese)
[10] 洪巧珍,江碧媛,黄振声.荔枝细蛾之饲育技术及其羽化与交尾行为 [J].植物保护学会会刊(台湾),2002,44(2):86-99.
Hong Q Z,Jiang B Y,Huang Z S.Rearing techniques,eclosion and mating behavior of litchi fruit borer,ConopomorphasinensisBradley (Lepidoptera:Gracillariidae) [J].Plant Protection Bulletin Taipei,2002,44(2):86-99.(in Chinese)
[11] 冯 莉,李敦松,张宝鑫,等.荔枝蒂蛀虫综合防治技术 [J].中国植保导刊,2004,24(4):24-26.
Feng L,Li D S,Zhang B X,et al.The control techniques ofConopomorphasinensisBradley [J].Plant Protection Bulletin China,2004,24(4):24-26.(in Chinese)
[12] 江世宏,杨长龙.荔枝蒂蛀虫繁殖生物学特性及饲养技术研究进展 [J].广西农业科学,2006,37(6):690-693.
Jiang S H,Yang C L.Advances in reproductive biology and rearing techniques ofConopomorphasinensisBradley [J].Guangxi Agricultural Sciences,2006,37(6):690-693.(in Chinese)
[13] 向玉勇.小地老虎性信息素的提取、鉴定及相关生物学研究 [D].贵州贵阳:贵州大学,2007:45-60.
Xiang Y Y.Extraction and identification of the black cutworms sex pheromone and correlative studies on its biology [D].Guiyang,Guizhou:Guizhou University,2007:45-60.(in Chinese)
[14] 董双林,杜家纬.交尾和温度对甜菜夜蛾(Spodopteraexigua)性信息素产生的影响 [J].应用生态学报,2002,13(12):1633-1636.
Dong S L,Du J W.Effects of mating experience and temperature on sex pheromone production of beet armyworm,Spodopteraexigua[J].Chinese Journal of Applied Ecology,2002,13(12):1633-1636.(in Chinese)
[15] 陆鹏飞,乔海莉,王小平,等.豆野螟成虫行为学特征及性信息素产生与释放节律 [J].昆虫学报,2007,50(4):335-342.
Lu P F,Qiao H L,Wang X P,et al.Adult behavior and circadian rhythm of sex pheromone production and release of the lengume pod bore,Marucavitrata(Fabricius)(Lepidoptera:Pyralidae) [J].Acta Entomologica Sinica,2007,50(4):335-342.(in Chinese)
[16] 杨美红,张金桐,刘金龙,等.榆木蠹蛾生殖行为及信息素产生与释放节律 [J].昆虫学报,2010,53(11):1273-1280.
Yang M H,Zhang J T,Liu J L,et al.Reproductive behavior and circadian rhythm of sex pheromone production and release ofHolcocerusvicarious(walker) (Lepidoptera:Cossidae) [J].Acta Entomologica Sinica,2010,53(11):1273-1280.(in Chinese)
猜你喜欢
——育王批次和场地的安排
蜜蜂杂志(2023年9期)2023-11-26 23:52:55
公民与法治(2022年12期)2023-01-07 09:17:38
公民与法治(2022年4期)2022-08-03 08:20:44
中国蜂业(2021年7期)2021-08-02 01:05:20
四川蚕业(2021年3期)2021-02-12 02:38:48
昆虫学报(2020年6期)2020-08-06 06:42:52
河北大学学报(自然科学版)(2017年2期)2017-04-26 05:26:26
蚕学通讯(2017年3期)2017-03-23 08:00:02
中国蜂业(2016年9期)2016-01-30 06:04:45
快乐作文·高年级(2014年6期)2014-08-16 01:31:07
