
光周期对莲草直胸跳甲幼虫发育的影响
2014-03-25 00:55:56赵龙龙刘蓉郭艳琼贾栋李娜马瑞燕
山西农业大学学报(自然科学版) 2014年3期
赵龙龙,刘蓉,郭艳琼,贾栋,李娜,马瑞燕
(山西农业大学 农学院,山西 太谷 030801)
喜旱莲子草Althernantheraphiloxeroides是一种多年生的草本植物,起源于南美洲巴拉那河流域,现已扩散到北美洲、南美洲、大洋洲、澳洲、亚洲等洲,其中包括美国、澳大利亚、新西兰、东南亚各国、印度和中国等多个国家和地区[1,2]。由于喜旱莲子草具有较强的耐受和干扰能力,适应多种生境,且难以根除,对入侵地的生态系统造成灾难性的影响,现已被许多国家列为典型的入侵杂草[3~5]。中国国家环保总局于2003年将喜旱莲子草列入《中国第一批外来入侵物种名单》。
对喜旱莲子草的防治,采用化学防治手段可迅速在短时间内清除喜旱莲子草,但因喜旱莲子草多生长在水域或湿地,残留的农药易渗入饮水和河流系统,不仅威胁到人畜健康,而且也威胁到昆虫及其它物种的生长。喜旱莲子草在入侵地主要以无性繁殖的方式进行扩散,在物理防治喜旱莲子草的过程中,常对喜旱莲子草进行片段化处理,所产生的茎节、根节片段均具有较高的再生能力,如这些碎片处理不善,可萌生成新的植株,即间接帮助了喜旱莲子草的繁殖和扩散[4,6]。综合比较几种防治措施,引进喜旱莲子草的专食性天敌莲草直胸跳甲Agasicleshygrophila是一种经济、环保、高效的手段,已被世界多个国家引入并推广。我国于1986年引入该天敌,经安全测试后,在我国南方多地释放并取得良好收效,为生物防治中的成功案例之一[7]。
利用莲草直胸跳甲防治喜旱莲子草虽取得一定收效,但在实际调查中发现,喜旱莲子草的分布面积仍大于天敌莲草直胸跳甲的分布。在高纬地区,该天敌少有分布或种群随季节波动变化巨大,影响其控草效果[8]。研究表明,高温和低温均能造成莲草直胸跳甲的大量死亡[9,10]。此外,不同生态型的喜旱莲子草对莲草直胸跳甲化蛹能力也有明显的影响[11,12]。虽然这些研究可在一定程度上说明莲草直胸跳甲种群分布界限及季节波动的原因,但在自然环境中,影响昆虫种群分布和变化的常为多因素综合的结果,除温度、食物资源外,光周期也是影响昆虫生长和发育的重要因子之一。其作用方式主要包括诱导昆虫进入滞育状态及影响昆虫的生长、发育等[13~18]。莲草直胸跳甲同其寄主植物喜旱莲子草均起源于南美洲,在引入我国后,经历了不同纬度梯度的变化,而纬度的变化直接决定着光周期的变化特点。关于光周期对莲草直胸跳甲的生长发育的影响少有报道。因此,本文通过设置不同组合的光周期条件,研究其对莲草直胸跳甲幼虫生长和发育的影响,以明确光周期对莲草直胸跳甲幼虫的影响作用。
1 材料和方法
1.1 供试虫源
供试莲草直胸跳甲虫源采集于华南野外种群,在实验室用温室常年种植的喜旱莲子草进行喂养并作为试验材料。
1.2 试验方法
试验所用莲草直胸跳甲幼虫为采集同日成虫所产卵块,采卵时间集中在1 h内,于人工气候箱中进行饲养,温度为25~27℃,光照:黑暗(L∶D)=12∶12(L表示光期时长,D表示暗期时长),相对湿度为85%。莲草直胸跳甲预化蛹时,从田间取新鲜的带有5节粗茎的喜旱莲子草,栽入盛有清水的钵头,每钵10株,将老熟幼虫接在喜旱莲子草上后,外置玻璃罩,放置待其化蛹。
试验温度设置为25℃,光周期为24 h循环光周期,共设置为9组,分别为D∶D,L∶D=2∶22,L∶D=4∶20,L∶D=8∶16,L∶D=16∶8, L∶D=20∶4,L∶D=22∶2,L∶L,对照光周期条件为L∶D=12∶12。将初孵的幼虫转入设置的光周期条件下,每处理6个重复,每重复15头幼虫。每龄期幼虫发育历期的计算方式为每日定时观察幼虫的发育情况,当有该龄幼虫出现时,其所用日数为其发育历期并统计该龄幼虫所占的比率和存活情况。待幼虫发育至3龄末时,接于喜旱莲子草茎秆上化蛹,记录羽化头数和化蛹历期。
1.3 数据统计
利用Microsoft Excel软件进行数据记录,并用SPSS进行统计学相关分析。试验中测得百分率经对数或者反正弦转化后,对其进行单因素方差分析(One-way ANOVA)。
2 结果与分析
2.1 光周期对莲草直胸跳甲1~2龄幼虫发育历期的影响
结果表明,25℃条件下,光周期对莲草直胸跳甲1~2龄幼虫发育历期有明显的影响。在不同光周期条件下,幼虫的发育历期有显著差异(F=6.812,df=(8,53),P<0.05)。1~2龄幼虫的发育历期多分布在3~4日,对照条件下,幼虫发育所需的历期最短。在光周期为L∶D=2∶22,L∶D=4∶20,L∶D=8∶16和L∶D=16∶8,跳甲1龄发育所需的时长最大;光周期L∶D=20∶4,L∶D=22∶2和L∶L下的幼虫发育历期次之(图1)。在所设置的不同光周期中,2龄幼虫所占比例在不同的光周期条件下,具有极显著差异(F=3.648,df=(8,53),P<0.05)。在初始阶段D∶D、L∶D=2∶22和L∶D=4∶20,随着光周期中光期时长比例的增加,2龄幼虫所占数目也逐渐增多,但在光周期为L∶D=8∶16,L∶D=20∶4和全光条件时,则与之相反(图2)。

图1 不同光周期对莲草直胸跳甲1~2龄幼虫发育历期的影响Fig.1 Effects of different photoperiods on the 1st ~2nd larvae development duration
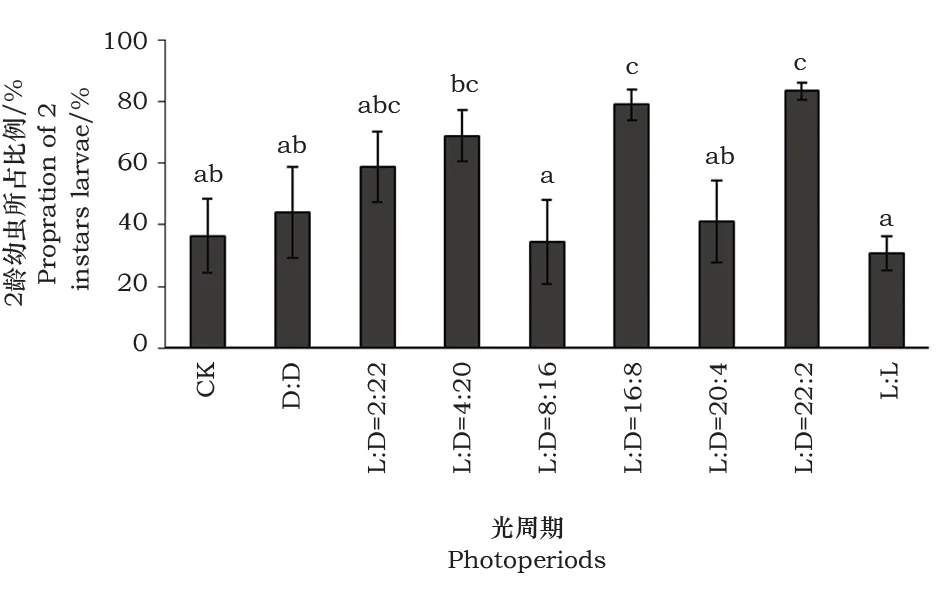
图2 不同光周期下莲草直胸跳甲2龄幼虫所占比率(平均发育历期内)Fig.2 The proportion of 2nd instar larvae under different photoperiods (mean developmental period)
2.2 光周期对莲草直胸跳甲2~3龄幼虫发育历期的影响
莲草直胸跳甲2龄幼虫在不同的光周期条件下其发育历期差异极显著(F=3.47,df=(8,53),P<0.05)。在不同光周期条件下,2龄幼虫在光周期L∶D=20∶4下的发育历期最短,光周期L∶D=16∶8条件下次之,但和其他组之间无明显差异,均在2~3日(图3)。对莲草直胸跳甲2龄幼虫的发育进度进行统计发现(图4),不同光周期处理下的3龄幼虫发育进度均衡,各个处理间并无显著差异(P>0.05)。3龄幼虫所占的比率多分布在30%~60%之间。
2.3 光周期对莲草直胸跳甲幼虫存活率的影响
莲草直胸跳甲幼虫在不同的光周期条件下,所表现的存活率有明显差异(F=9.014,df=(8,53),P<0.05)幼虫的存活率随着光周期中光照时长比例的增加而提高,但在光周期L∶D=8∶16条件下,幼虫所表现出的存活率却最低,全光条件下存活率次之(图5)。

图3 不同光周期对莲草直胸跳甲2~3龄幼虫发育历期的影响Fig.3 Effects of different photoperiods on the 2nd~3rd larvae development duration
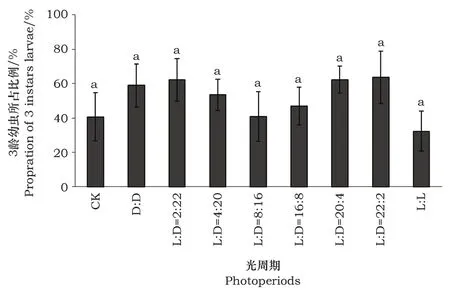
图4 不同光周期下莲草直胸跳甲3龄幼虫所占比率Fig.4 The proportion of 3rd larvae under different photoperiods

图5 不同光周期下莲草直胸跳甲幼虫的存活率Fig.5 The larvae survival rates under different photoperiods
2.4 光周期对化蛹的影响
经光周期处理后的莲草直胸跳甲幼虫接于化蛹装置中,置于室温饲养,经统计学分析发现不同光周期处理后的莲草直胸跳甲幼虫所表现出的化蛹率差异显著(F=1.71,df=(8,26),P<0.05)(图6)。在光周期L∶D=4∶20条件下化蛹率最低,全光条件下次之。其它组之间无显著差异,化蛹率均在半数以上,以对照组最高为70%。试验测得其化蛹历期多分布在9~10日,且各个处理之间差异显著(F=4,df=(8,26),P<0.05)。以在光周期L∶D=22∶2条件下所需的化蛹历期最短(图7)。

图6 不同光周期下莲草直胸跳甲幼虫的化蛹率Fig.6 The pupation rates under different photoperiods

图7 不同光周期对莲草直胸跳甲化蛹历期的影响Fig.7 The pupation duration under different photoperiods
3 讨论与结论
3.1 光周期对莲草直胸跳甲幼虫发育的影响
光照对昆虫生长、发育具有间接或直接的影响作用,如有些昆虫只有在光照条件下才进行活动或者取食,所以光照条件间接的调节了昆虫的生长发育[14,17]。本研究发现莲草直胸跳甲不同龄期幼虫对光周期的敏感程度不同,在幼虫的1龄阶段,当光周期的光期在2~16 h时,对其影响明显,所需发育历期最大。除光周期L∶D=8∶16外,其幼虫所表现的发育历期随光期时长比例增加而增高。这表明在此光周期条件下,跳甲幼虫较其他光周期下敏感,但并未抑制其个体发育,其个体发育进度只受一定范围内的光期调节,但全光条件对其有明显的抑制作用。莲草直胸跳甲2龄幼虫对光周期的响应模式同1龄幼虫,光周期的光期在2~16 h时,对幼虫发育历期影响明显,但对其整体发育进度影响较小。综合比较莲草直胸跳甲幼虫在所设置的光周期条件下,光周期中光期时长比例对其发育有明显的影响作用,幼虫对光周期敏感的临界点分布在L∶D=8∶16左右,即莲草直胸跳甲幼虫可能对光照不敏感或对光的反应为负趋光性,这也影响到跳甲幼虫的取食习性,多在喜旱莲子草的叶背位置进行取食[19]。
3.2 光周期对莲草直胸跳甲幼虫存活率的影响
光周期对莲草直胸跳甲幼虫存活率的影响和其发育程度呈相反的规律,除光周期L∶D=8∶16和全光外,莲草直胸跳甲幼虫的发育历期随着光周期中光照时长而增加,但其成活率却呈递增的趋势,可能是跳甲延时发育,导致营养物质积累,进而提高了跳甲的存活能力。由此可得出,在跳甲的人工饲养过程中,可适当延长饲养过程中光周期中的光期比例,以提高跳甲的存活能力。
3.3 光周期对莲草直胸跳甲化蛹能力的影响
在许多昆虫中,光周期对其发生作用,并不直接作用于当代的个体,而是将这种影响作用积累在下一个虫态或者后代中表现出来。最典型的例子如滞育的昆虫个体,通常是前一个虫态或者母体感受到光周期的变化,而在下一个虫态或者子代中表现出滞育的特征[20]。在本研究中,莲草直胸跳甲幼虫的整个发育阶段置于不同的光周期条件下,在化蛹过程中解除了这种条件,于正常条件下饲养,发现不同光周期的处理,并未对其发生作用。其化蛹历期多分布在8~10日,均未出现明显的发育停滞现象,说明跳甲幼虫不会将光周期影响在其后续虫态中表现出来。在光周期L∶D=4∶20条件下,出现较低化蛹率的原因可能是人为选择喜旱莲子草的茎秆差异所造成的,因不同生物型和不同营养含量的喜旱莲子草对其化蛹能力均有影响[11,12]。
莲草直胸跳甲引种中国后,引种地的光周期条件和其原产地相比较发生了较大的变化。本研究证实了不同光周期条件对莲草直胸跳甲幼虫的发育历期和存活率都有明显的影响作用,进而影响到莲草直胸跳甲种群的繁盛程度。这也说明了在不同的季节中,跳甲种群波动变化的原因。但在自然环境中,光周期并不是唯一调控昆虫的因子,常和温度等耦连在一起共同作用,因此在讨论和评价光周期对跳甲的作用时,应该联系温度这一环境因子,共同探讨对莲草直胸跳甲种群的影响作用,并进一步为生物防治喜旱莲子草提供理论依据。
参 考 文 献
[1]张光富,陈瑞冰.外来入侵植物喜旱莲子草的研究进展[J].安徽大学学报:自然科学版, 2005, 29(6):87-93.
[2]潘晓云,耿宇鹏,Alejandro SOSA,等.入侵植物喜旱莲子草——生物学、生态学及管理[J].植物分类学报,2007,45(6):884-900.
[3]许凯扬,叶万辉,段学武,等.PEG诱导水分胁迫下喜旱莲子草的生理适应性[J].浙江大学学报:农业与生命科学版,2004,30(3):37-43.
[4]贾昕,杨兴中,潘晓云,等.喜旱莲子草营养繁殖特征对干扰的响应[J].生物多样性, 2008, 16(3): 229-235.
[5]许凯扬,叶万辉,李国民,等.入侵种喜旱莲子草对光照强度的表型可塑性反应[J].武汉植物学研究, 2005,23(6):560-563.
[6]张彩云,刘卫,徐志防,等.入侵种喜旱莲子草和莲子草的营养生长和光合作用对温度的响应[J].热带亚热带植物学报,2006,14(4):333-339.
[7]马瑞燕,王韧.喜旱莲子草在中国的入侵机理及其生物防治[J].应用与环境生物学报, 2005, 11(2):246-250.
[8]陈立立,余岩,何兴金.喜旱莲子草在中国的入侵和扩散动态及其潜在分布区预测[J].生物多样性, 2008,16(6):578-585.
[9]赵鑫,傅建炜,万方浩,等.短时高温暴露对莲草直胸跳甲生殖特性的影响[J].昆虫学报, 2009, 52(10):1110-1114.
[10]陈磊,蔡笃程,陈青,等.短时高温对莲草直胸跳甲成虫存活及繁殖的影响[J].昆虫知识, 2010, 47(2):308-312.
[11]郝炯,刘艳红,张萍萍,等.喜旱莲子草的营养累积及对莲草直胸跳甲化蛹能力的影响[J].中国农业大学学报,2011,16(4):48-51.
[12]马瑞燕,王韧.不同生态型的喜旱莲子草对莲草直胸跳甲化蛹能力的影响[J].植物生态学报, 2004,28(1):24-30.
[13]陈夜江,罗宏伟,黄建,等.光周期对烟粉虱实验种群的影响[J].华东昆虫学报,2003,12(1):38-41.
[14]陈广平,郝树广,庞保平,等.光周期对内蒙古三种草原蝗虫高龄若虫发育、存活、羽化、生殖的影响[J].昆虫知识,2009,46(1):51-56.
[15]易传辉,陈晓鸣,史军义,等.光周期对枯叶蛱蝶幼虫生长发育的影响[J].西北林学院学报, 2008, 23(5):124-126.
[16]许永玉,牟吉元,胡萃,等.光周期和温度对中华通草蛉成虫生殖的影响[J].华东昆虫学报, 2002, 11(1):39-43.
[17]易传辉,陈晓鸣,史军义,等.光周期对柑橘凤蝶幼虫生长发育及存活的影响[J].西北农业学报, 2009,18(1):162-164,169.
[18]蔡明飞,刘彦飞,王艳蓉,等.光周期对梨小食心虫生长发育和生殖的影响[J].西北农业学报, 2010,19(11):169-172.
[19]陈磊,蔡笃程,陈青,等.莲草直胸跳甲成虫产卵选择性及叶位与幼虫发育适合度的关系[J].热带作物学报,2009,30(3):353-356.
[20]徐卫华.昆虫滞育研究进展[J].昆虫知识,2008,45(4):512-517.
猜你喜欢
遗传(2023年9期)2023-09-25 09:31:44
农业科技通讯(2023年1期)2023-02-12 07:08:50
中老年保健(2022年7期)2022-09-20 01:16:16
湖北农业科学(2021年22期)2021-12-13 13:00:36
红领巾·探索(2018年12期)2018-01-26 12:34:14
少儿科学周刊·少年版(2017年7期)2017-09-29 08:02:41
少儿科学周刊·儿童版(2017年7期)2017-09-29 02:22:58
小学生作文选刊(2017年6期)2017-07-03 12:16:36
中国稻米(2017年2期)2017-04-28 08:00:06
丝路艺术(2017年5期)2017-04-17 03:11:51
