
不同硒源及水平对肉鸡生长性能、血浆和组织硒含量及血浆谷胱甘肽过氧化物酶活性的影响
2014-03-14 03:40:38郭军蕊刘国华郑爱娟常文环蔡辉益
动物营养学报 2014年7期
郭军蕊 刘国华 郑爱娟 常文环 张 姝 蔡辉益
(中国农业科学院饲料研究所,农业部饲料生物技术重点开放实验室,北京 100081)
硒于1817年首次被发现,100多年后,硒以机体必需的微量元素被大家所认可,于是研究者开始对动物对硒的需要量,硒对动物的生长、肉质等的影响展开研究。随后,硒的抗氧化[1]、免疫[2]、促生长[3]、提高繁殖性能[4]、抗应激[5-7]、抗癌[8]等作用逐步被研究证实。由于无机硒有高毒性等缺点,日本等一些国家已禁止在饲粮和食品中添加亚硒酸钠(Na2SeO3)。有机硒的使用逐渐引起人们的重视。关于有机硒在动物体内的沉积及抗氧化性的研究在国内外有很多报道,但是对不同日龄、不同水平的硒沉积和抗氧化性报道较为少见。本试验主要研究了亚硒酸钠、酵母硒和不同水平蛋氨酸硒对不同日龄肉鸡生长性能、血浆和组织硒含量及血浆谷胱甘肽过氧化物酶(glutathione peroxi-dase,GSH-Px)活性的影响,旨在比较有机硒与无机硒在肉鸡上的作用效果,为有机硒在生产中的应用提供试验依据。
1 材料与方法
1.1 试验动物及分组
选用1日龄健康爱拔益加(AA)肉雏鸡756只,随机分为组1~7共7组,组1饲喂低硒基础饲粮(负对照),组2~7在基础饲粮的水平上分别添加 0.1、0.3、0.5、0.7 mg/kg 蛋氨酸硒、0.3 mg/kg酵母硒(正对照)和0.3 mg/kg亚硒酸钠(正对照)。每组6个重复,每个重复18只鸡(公母各占1/2)。试验期为42 d。
1.2 试验材料
亚硒酸钠,饲料级,硒含量为1%;酵母硒,硒含量为2 000 mg/kg;蛋氨酸硒,硒含量为2 000 mg/kg。
1.3 试验饲粮
本试验基础饲粮参考《肉鸡饲养标准》(NY/T 33—2004)配制(粉料),其组成及营养水平见表1。

表1 基础饲粮组成及营养水平(风干基础)Table 1 Composition and nutrient levels of basal diets(air-dry basis) %
1.4 饲养管理
试验分2阶段进行,1~21日龄为育雏期,22~42日龄为育肥期。采用立体3层笼养,24 h光照,乳头式饮水器供水,自由采食。1日龄接种马立克氏病、鸡新城疫苗,21日龄饮水接种法氏囊病疫苗。鸡舍卫生管理按常规进行。
1.5 样品采集与处理
1.5.1 生长性能
1~21日龄和22~42日龄分别对每个重复鸡进行称重,截料称料;记录每天鸡只死淘只数。计算1~21日龄、22~42日龄及1~42日龄初体重、末体重、平均日增重(average daily gain,ADG)、平均日采食量(average daily feed intake,ADFI)、料重比(feed/gain,F/G)和死淘率。
1.5.2 血浆指标
7个组每重复随机选取7、21、35和42日龄鸡2只(公母各1只),采集血浆3~5 m L。采血前禁食12 h,自由饮水。采集的血液置于肝素抗凝管内,轻轻混匀后静置,待有少量血浆析出后,2 000 r/m in离心 10 m in,取上清液于 EP管中,-70℃保存待测血浆GSH-Px活性和血浆硒含量。
1.5.3 组织指标
试验结束时,7个组每重复随机选取2只鸡(公母各1只),供水、禁食12 h后放血并进行解剖,采集胸肌、肝脏、肾脏、皮肤和腹脂,-20℃保存待测硒含量。
1.6 指标及测定方法
1.6.1 血浆和组织硒含量的测定
参照GB/T 13883—2008,采用原子荧光光谱法测定血浆和组织硒含量。
1.6.2 血浆 GSH-Px活性测定
采用5,5’-二硫硝基苯甲酸(DTNB)比色法,按照南京建成生物工程研究所提供的试剂盒测定血浆GSH-Px活性。
1.7 数据统计与分析
试验数据采用SPSS 17.0软件的一般线性模型(GLM)程序进行双因素方差分析,因子显著性采用F检验。对主效应显著的指标进行最小显著差数(LSD)法多重比较,显著和极显著水平分别设为0.05和0.01,试验数据表示为平均值±标准差表示。采用曲线估计(curve estimate)程序对蛋氨酸硒添加水平与组织硒含量、血浆硒含量及GSH-Px活性进行线性和二次模型回归分析。
2 结果
2.1 不同硒源和水平对肉鸡生长性能的影响
由表2、表3、表4可知,不同硒源和水平饲粮处理对1~21日龄、22~42日龄及1~42日龄肉鸡的 ADG、ADFI、F/G、初体重、末体重及死淘率无显著影响(P>0.05)。42日龄的末体重,各加硒组高于负对照组(P>0.05)。
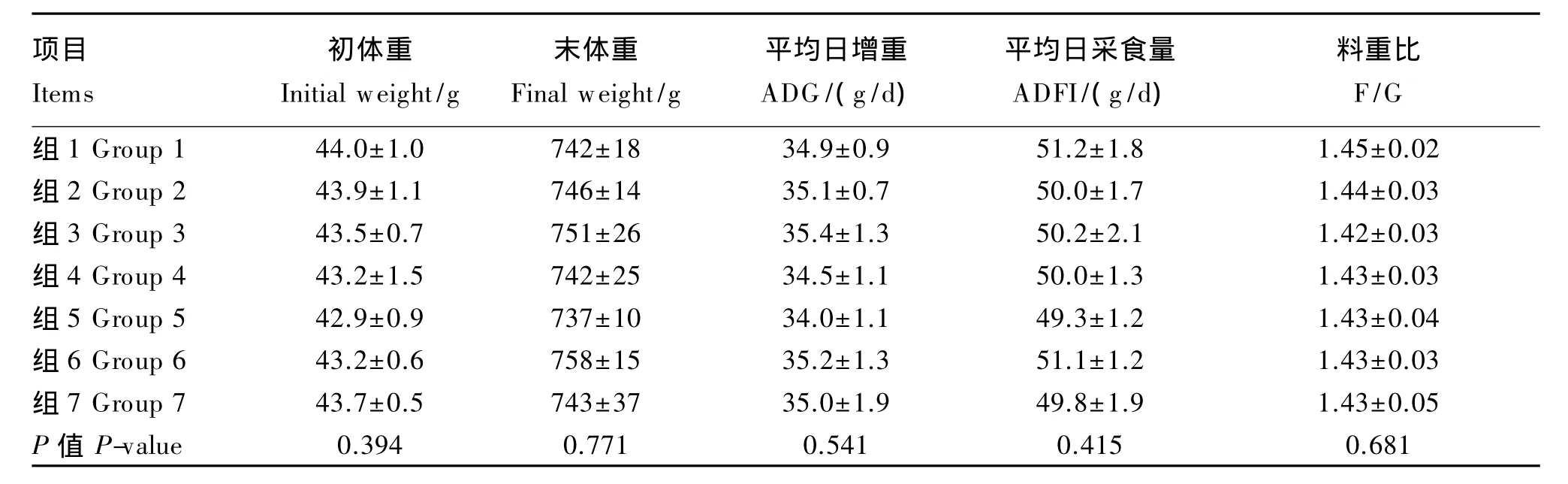
表2 不同硒源和水平对1~21日龄肉鸡生长性能的影响Table 2 Effects of different selenium sources and supplemental levels on grow th performance of broilers at 1 to 21 days of age(n=6)
2.2 不同硒源和水平对血浆硒含量和 GSH-Px活性的影响
由表5可知,对于不同日龄血浆硒的含量,各加硒组硒含量均显著高于负对照组(P<0.05),且随着硒添加水平的增加而增大,相同添加水平蛋氨酸硒组优于2个正对照组,日龄和添加水平之间存在极显著交互作用(P<0.01);对于不同日龄血浆GSH-Px活性,各加硒组血浆GSH-Px活性显著高于负对照组(P<0.05),且组2 GSH-Px活性显著低于其他各加硒组(P<0.05),日龄和添加水平之间存在极显著互作效应(P<0.01)。对于7、35和42日龄血浆GSH-Px活性,组3显著高于组4、组5(P<0.05),21日龄时组4 GSH-Px活性显著高于其他各组(P<0.05)。结合表6和表7可知,不同日龄血浆硒含量有线性(P<0.01)及二次(P<0.01)剂量效应,随饲粮硒添加水平呈线性(P<0.01)和二次性(P<0.01)增长;不同日龄 GSH-Px活性也有相同趋势,从决定系数R2来看,更符合二次剂量关系,呈抛物线型的二次曲线(P<0.01)。
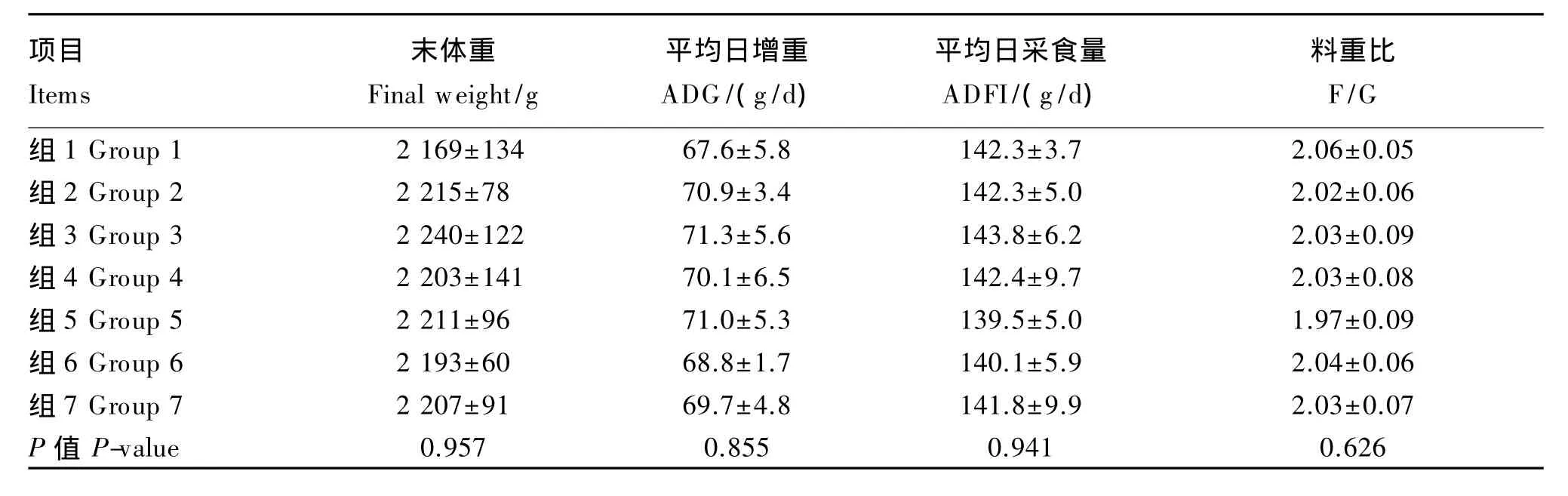
表3 不同硒源和水平对22~42日龄肉鸡生长性能的影响Table 3 Effects of different selenium sources and supplemental levels on grow th performance of broilers at22 to 42 days of age(n=6)
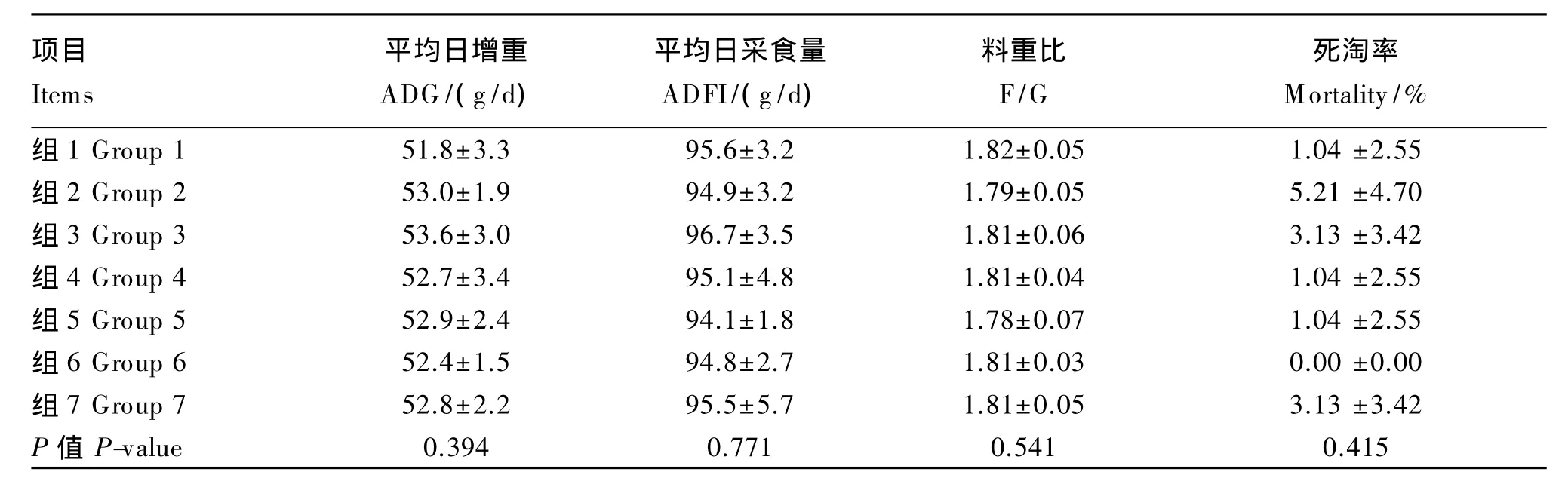
表4 不同硒源和水平对1~42日龄肉鸡生长性能的影响Table 4 Effects of different selenium sources and supplemental levels on grow th performance of broilers at1 to 42 days of age(n=6)
2.3 不同硒源和水平对组织硒含量的影响
由表8可知,对于42日龄肝脏、肾脏、肌肉、皮肤和腹脂硒含量,各加硒组组织硒含量随添加水平的增加而增大(P>0.05),且均显著高于负对照组(P<0.05),相同添加水平下,蛋氨酸硒组优于2个正对照组(P>0.05)。另外,本试验结果显示组织硒含量高低依次为肾脏、肝脏、胸肌、皮肤和腹脂。结合表9可知,42日龄肉鸡肝脏、肾脏、肌肉、皮肤和腹脂硒含量存在线性(P<0.01)及二次(P<0.01)剂量效应。
3 讨论
3.1 不同硒源和水平对肉鸡生长性能的影响
有关硒源对动物生长性能的影响不同研究者的结论不尽相同。Upton等[9]证明加硒(亚硒酸钠和酵母硒)组的饲料转化率优于对照组(P<0.05),而添加 0.2 mg/kg亚硒酸钠组 AA 肉鸡体重(BW)与对照组差异不显著(P>0.05)。相同硒添加水平无机硒和有机硒对BW和饲料转化率影响差异不显著(P>0.05),但有机硒优于无机硒。李业国等[10]研究表明,饲喂酵母硒饲粮的肉鸡4周龄ADG、BW和F/G显著优于无机硒饲粮组(P<0.05)。许飞利[11]对黄羽肉鸡的研究也发现无机硒和有机硒对49日龄肉鸡F/G有改善作用(P<0.05)。而Payne等[12]试验报道却表明,硒源(亚硒酸钠和酵母硒)对生长性能影响不显著(P>0.05)。王秀娜[13]和王宝维等[14]发现不同硒源对猪和鹅的生长性能无显著影响(P>0.05)。本试验未发现硒源之间对肉鸡生长性能有显著影响(P>0.05),这可能与鸡的品种、饲养环境、饲养周期或者基础饲粮的硒含量有关。

表5 不同硒源和水平对肉鸡血浆硒含量和GSH-Px活性的影响Table 5 Effects of different selenium sources and supplemental levels on selenium content and GSH-Px activity in plasma of broilers(n=12)

续表5

表6 蛋氨酸硒添加水平(X)与血浆硒含量(Y)的回归方程Table 6 Regression equation of selenium methionine supplemental level(X)and selenium content in plasma(Y)

表7 蛋氨酸硒添加水平(X)与血浆GSH-Px活性(Y)的回归方程Table 7 Regression equation of selenium methionine supplemental level(X)and GSH-Px activity in plasma(Y)
不同研究者关于硒水平与生长性能关系的研究结果有较大差异。李业国等[10]研究表明不同的硒水平可显著影响 ADG、ADFI和 F/G(P<0.05),且添加0.15 mg/kg酵母硒的4周和7周肉鸡BW显著高于添加 0.3 mg/kg 酵母硒组(P<0.05);而亚硒酸钠则相反,添加0.3 mg/kg组4周和7周肉鸡 BW 显著高于添加 0.15 mg/kg 组(P<0.05)。Ševcˇikova 等[15]认为酵母硒和小球藻硒与对照组相比,可以显著提高BW(P<0.05),但对改善饲料转化率无显著影响(P>0.05)。Chantiratikul等[16]研究发现,不同硒源对蛋鸡生产性能无显著影响(P>0.05)。刘明宝[17]在基础饲粮(含硒量为0.018 5 mg/kg)中 添 加 0.02、0.10、0.50 和2.50 mg/kg亚硒酸钠饲喂猪结果表明,各处理间猪ADG、ADFI和 F/G 均无显著差异(P>0.05)。宋清华等[18]研究也表明,同一硒源不同水平对良凤花肉鸡ADG和F/G无显著影响(P>0.05)。朱宏娟[19]和崔国强等[20]也得出相同的结论。本试验也发现,饲粮硒水平对肉鸡生产性能没有显著影响(P>0.05),且 即 使 硒 水 平 低 至 0.035~0.042 mg/kg(基础饲粮)也未能影响肉鸡的生长(P>0.05)。罗培林等[21]在大鼠硒耗竭试验中也发现,在硒含量为0.03 mg/kg的基础饲粮中添加不同水平的亚硒酸钠和酵母硒并未影响经2周硒耗竭 大 鼠 的 ADFI、ADG 及 F/G(P>0.05)。Thompson等[22]研 究 则 证 明,当 鸡 饲 粮 中 含100 mg/kg的维生素 E时,硒最低需要量低于0.01 mg/kg;而当维生素 E含量为10 mg/kg时,添加 0.02 mg/kg的硒(基础饲粮硒含量<0.005 mg/kg)即可维持鸡的正常生长;当饲粮中不含维生素E时,硒的最低需要量则接近0.05 mg/kg,与本试验的结果近似。这可能暗示满足动物正常生长的最低硒需要量与饲养试验条件有关,即在严格控制的试验条件下,动物较少受不利环境因素的刺激,其最低需要量有可能远低于生产条件下的需要量,但尚需更多的试验证据证实。
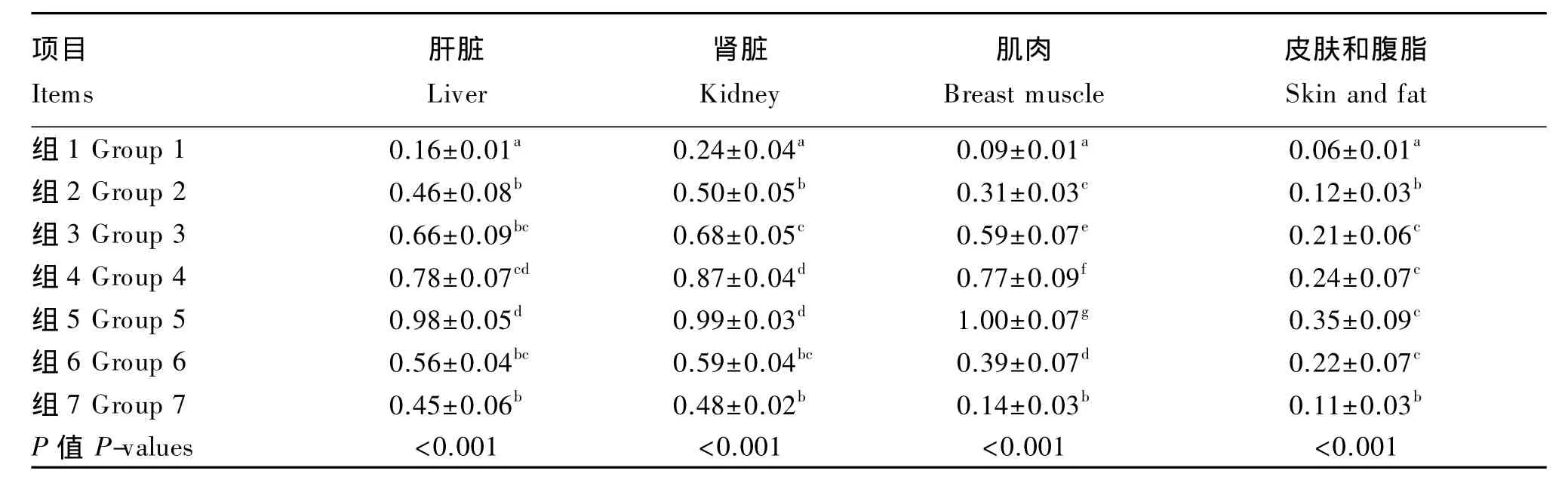
表8 不同硒源和水平对组织硒含量的影响Table 8 Effects of different selenium sources and supplemental levels on tissue selenium content(n=12) mg/kg
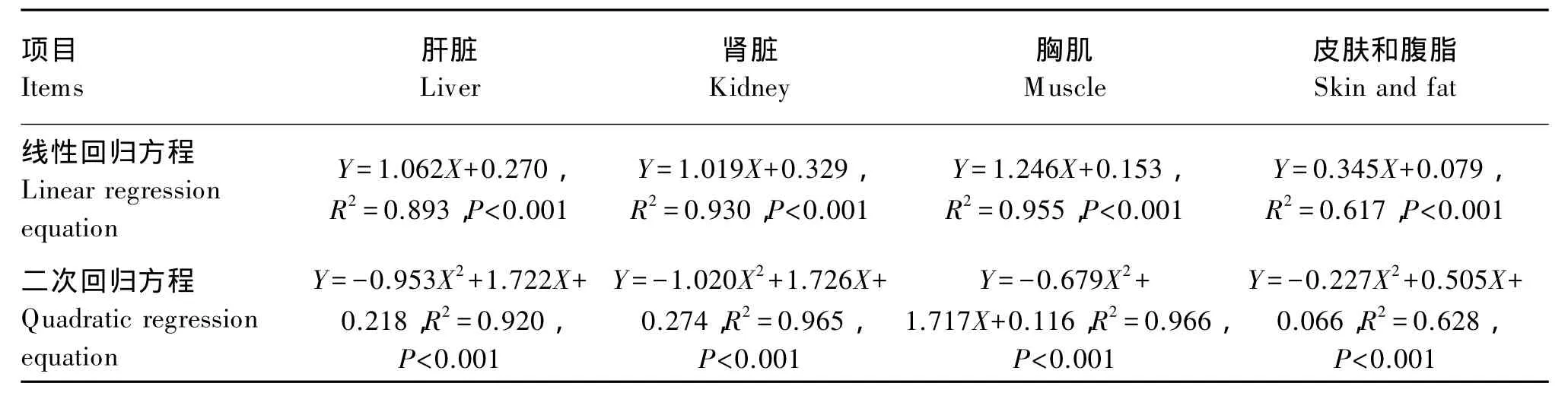
表9 蛋氨酸硒添加水平(X)与组织硒含量(Y)的回归方程Table 9 Regression equation of selenium methionine supplemental level(X)and tissue selenium content(Y)
3.2 不同硒源和水平对血浆硒含量和 GSH-Px活性的影响
硒在十二指肠被吸收进入血液后,与血液中的α-球蛋白、β-球蛋白结合,经血浆运载进入各组织[23]。研究表明,饲喂硒代蛋氨酸的肥育猪血浆中的硒含量显著高于相同添加水平的亚硒酸钠组(P<0.05)[24]。王海宏[25]研究不同硒源对肉鸡血浆硒含量的影响发现,21日龄时蛋氨酸硒组极显著高于无机硒和酵母硒组(P<0.001),42日龄时不同硒源无显著差异(P>0.05)。据林长光等[26]报道,与亚硒酸钠相比,泌乳母猪饲粮添加纳米硒显著提高仔猪血浆硒含量(P<0.05),且添加亚硒酸钠组的仔猪血浆硒含量显著低于对照组(P<0.05)。本试验结果也发现,添加蛋氨酸硒组肉鸡的7和42日龄血浆硒含量均显著高于无机硒组(P<0.05)。显示有机硒和无机硒在吸收和运载机制上存在较大差异,与无机硒相比,有机硒似乎更易于被动物吸收。
本试验证实,血浆硒含量与饲粮硒水平之间存在极显著的剂量效应(P<0.01),血浆硒含量随着饲粮硒水平增加而呈线性升高趋势。肥育猪试验也发现,饲粮中添加硒代蛋氨酸或者亚硒酸钠均可提高肥育猪血浆硒含量[24]。添加 0.5和0.7 mg/kg酵母硒或纳米硒均显著提高泌乳期母猪血浆硒含量(P<0.05)[26]。Juniper等[27]在肉牛试验中也得到了相似的结果。可见血浆硒含量可作为敏感指标用于判断动物的硒营养状况。
本研究还发现,蛋氨酸硒添加水平与血浆硒含量的关系在一定程度上受日龄影响。7日龄和21日龄时,血浆硒含量随蛋氨酸硒添加水平的提高而呈增加趋势,当饲粮硒添加水平达到0.5 mg/kg时,血浆硒含量最高,进一步提高硒添加水平血浆硒含量不再随之增加。而在35和42日龄时,血浆硒含量随蛋氨酸硒添加水平的提高而持续增加,添加0.7 mg/kg蛋氨酸硒组血浆硒含量达到最大值。一般认为,血浆硒含量受吸收和清除的双重影响。肉鸡生长前期饲粮硒添加水平达到0.5mg/kg时血浆硒含量即达到稳恒(硒含量为0.16 mg/kg),而后期即使饲粮硒添加水平高达0.7 mg/kg时也未达到稳恒(硒含量≥0.20 mg/kg),这有可能是因为生长前期肉鸡硒吸收途径更易达到饱和或其α-球蛋白、β-球蛋白与硒的亲和力更弱,且其生长迅速导致组织对血硒的利用和清除速率更快,而后期对硒的吸收能力、血浆亲和力发育性增强,且其相对生长速率降低对血硒的利用和清除速率也降低,导致其有较高的血硒含量。然而局限于目前对硒吸收、转运、组织利用机制研究尚不充分,其具体机制尚需进一步研究探讨。
GSH-Px是机体内广泛存在的一种过氧化物分解酶。硒半胱氨酸是其活性中心,酶活性大小可反映机体硒水平。机体中约30%~40%的硒以GSH-Px的形式存在,硒缺乏将引起GSH-Px活性变化,从而引起动物的疾病发生和功能紊乱[28]。GSH-Px能催化谷胱甘肽(GSH)变为氧化型谷胱甘肽(GSSG),使有毒的过氧化物还原成无毒的羟基化合物,从而保护细胞膜的结构及功能不受过氧化物的干扰及损害[29]。硒和GSH系统在抗氧化方面起到了重要作用[1]。最近,尹俐[30]制备了一种人工模拟GSH-Px-含硒谷胱甘肽硫转移酶,并且证明其在体外能够表达抗氧化作用。机体内的抗氧化/促氧化平衡,对鸡的健康、胚胎发育、精子质量以及家禽的生长与繁殖都非常重要[31]。本研究结果表明,与低硒基础饲粮组相比,加硒组血浆GSH-Px活性显著提高(P<0.05),该结果与许多报道[32-34]一致。研究表明,硒添加饲粮可以提高鸡血液中GSH-Px活性,且其活性与硒添加水平呈一定的相关性[35-36]。猪血清GSH-Px活性则随饲粮硒水平对数而呈线性(P<0.05)和二次性增长(P<0.05),呈一抛物线型的二次曲线(P<0.05)[17]。Wang 等[33]和蒋立等[37]的研究也表明添加硒可分别使肉鸡和肉兔血浆GSH-Px活性升高(P<0.05)。Wang 等[33]还发现 21 日龄血浆GSH-Px活性有机硒组显著高于无机硒组(P<0.05),这与本试验研究结果以及 Chen 等[38]的结果一致。而王海宏[25]的研究则表明相同添加水平有机硒与无机硒间差异不显著(P>0.05),但42日龄时无机硒效果优于酵母硒。本试验结果表明,相同添加水平的无机硒和蛋氨酸硒组差异不显著(P>0.05),但二者均显著高于酵母硒组(P<0.05)。回归分析表明,蛋氨酸硒添加水平与血浆GSH-Px活性存在显著的二次剂量关系(P<0.05),说明获得最高GSH-Px活性所需要的硒添加水平在 0.3~0.7 mg/kg。本试验还发现,日龄也是影响GSH-Px活性的因素,并存在一定的发育性规律,但这种发育规律因饲粮硒水平而有差异。当采食低硒饲粮时,血浆GSH-Px活性并未随日龄增长而发生显著变化(P>0.05),各日龄 GSH-Px活性均保持在较低水平上。但当硒添加水平为0.3 mg/kg及以上时,GSH-Px活性因日龄增长而提高,21、35和42日龄显著高于7日龄(P<0.05)。这一结果也间接说明充足的饲粮硒添加水平(或血浆硒含量)是GSH-Px正常表达的必要条件而并不仅仅是充分条件。同样地,饲粮硒添加水平对血浆GSHPx活性的影响也受日龄的影响,这种影响主要表现为7、35和42日龄肉鸡血浆GSH-Px活性达到最高值时,对应的蛋氨酸硒添加水平为0.3 mg/kg,而21日龄时对应的添加水平为0.5 mg/kg,这可能意味着肉鸡不同日龄时达到等活性GSH-Px所需的饲粮硒添加水平不同,但这种日龄效应的机制和生理意义尚不清楚,有待进一步的研究。
3.3 不同硒源和水平对组织硒含量的影响
有研究报道,随着饲粮中无机硒和有机硒(酵母硒、蛋氨酸硒)添加水平的增加,大鼠[17]、猪[17]和肉鸡[39-40]等的心脏、肝脏、肾脏等组织中硒含量随之升高,且还可提高孵化率和1日龄雏鸡组织硒含量[41-42]。大量研究表明,随着饲粮硒添加水平的增加,组织硒含量随之增加,且有机硒的沉积效果优于无机硒[43-45]。并有研究表明,0.2 mg/kg的酵母硒对组织硒含量来讲可替代0.3 mg/kg的亚硒酸钠[39],这与本研究结果相似。Dlouhá 等[46]研究绿藻硒和亚硒酸钠对肉鸡胸肌硒含量的影响表明,绿藻硒更有利于硒的沉积(P<0.05)。饲粮中有机硒添加效果明显优于无机硒,且大部分硒在畜禽体内以蛋氨酸硒形式替代蛋氨酸形成硒蛋白质,小部分硒则以半胱氨酸硒形式参与体蛋白质组成,在体内起广泛作用[47]。这也可能与有机硒和无机硒在体内的吸收代谢方式有关。有报道认为,无机硒代谢过程中,仅有少量结合于人体蛋白质中,大多数则进入肾脏,最终排出体外;而有机硒因与含硫氨基酸类似,在蛋白质的合成中常替代含硫氨基酸而结合于蛋白质中,因此有机硒能增加机体总硒的贮存量[48]。试验证明与无机硒相比,添加有机硒更有利于体组织硒的沉积。此外,回归分析结果表明,蛋氨酸硒添加水平与组织硒含量之间呈显著的线性及二次回归关系(P<0.001),说明提高饲粮蛋氨酸硒水平可明显增加组织硒的沉积量,且在硒添加水平高达0.7 mg/kg时组织硒含量也未达到饱和或平衡状态。
4 结论
①饲粮中添加亚硒酸钠、酵母硒和不同水平蛋氨酸硒对肉鸡生长性能无明显改善作用,但对42日龄的末体重稍有提高。
②饲粮中肉鸡血浆和组织硒含量随硒添加水平的升高而增加,且蛋氨酸硒优于亚硒酸钠和酵母硒。且对于血浆硒含量,日龄和硒添加水平之间有交互作用。
③饲粮中添加亚硒酸钠、酵母硒和不同水平蛋氨酸硒能显著提高肉鸡血浆GSH-Px活性,且日龄和硒添加水平之间存在交互作用。
[1] ARTEEL G E,SIES H.The biochem istry of selenium and the glutathione system[J].Environmental Toxicology Pharmacology,2001,10(4):153-158.
[2] KIREM IDJIAN-SCHUMACHER L,ROY M,W ISHE H I,et al.Supplementation w ith selenium augments the functions of natural killer and lymphokine-activated killer cells[J].Biological Trace Element Research,1996,52(3):227-239.
[3] ARTHUR JR,NICOL F,HUTCHINSON A R,et a1.The effects of selenium depletion and repletion on the metabolism of thyroid hormones in the rat[J].Journal of Inorganic Biochem istry,1990,39(2):101-108.
[4] W ILSONM E,ROZEBOOM K J,CRENSHAW T D.Boar nutrition for optimum sperm production[J].Advances in Pork Production,2004(15):295-306.
[5] 袁施彬.仔猪氧化应激及硒的抗应激效应和机理的研究[D].博士学位论文.雅安:四川农业大学,2007:45-69.
[6] FAN C,YU B,CHEN D W.Effects of different sources and levels of selenium on performance,thyroid function and antioxidant status in stressed broiler chickens[J].International Journal of Poultry Science,2009,8(6):583-587.
[7] M ILLER L L.The effects of selenium on the physiological stress response in fish[D].Master’s dissertation Lethbridge:University of Lethbridge,2006:51-59.
[8] 邵世和,孙丽媛.微量元素硒与肿瘤的关系[J].北华大学学报:自然科学版,2003,4(3):223-226.
[9] UPTON J R,EDENS F W,FERKET P R.Selenium yeast effect on broiler performance[J].International Journal of Poultry Science,2008,7(8):798-805.
[10] 李业国,郭峰,李同树.日粮不同硒源对肉仔鸡生产性能、肉质和血清甲状腺激素的影响[J].畜牧与兽医,2005,37(8):30-32.
[11] 许飞利.不同硒源对黄羽肉鸡生产性能、理化指标及肉品质的影响[D].硕士学位论文.石河子:石河子大学,2007:21-23.
[12] PAYNE R L,SOUTHERN L L.Comparison of inor-ganic and organic selenium sources for broilers[J].Poultry Science,2005,84(6):898-902.
[13] 王秀娜.不同硒源对仔猪生长性能及组织中GPX 1 mRNA表达量的影响[D].硕士学位论文.大庆:黑龙江八一农垦大学,2010:19-20.
[14] 王宝维,王娜,葛文华,等.不同硒源对鹅早期生产性能、屠宰性能、肉品质、肌肉常规养分、免疫与抗氧化功能的影响[J].中国农业科学,2011,44(14):3016-3026.
[15] ŠEVCˇIKOVÁ S,SKRˇIVAN M,DLOUHÁ G,et al.The effect of selenium source on the performance and meat quality of broiler chickens[J].Czech Journal of Animal Science,2006,51(10):449-457.
[16] CHANTIRATIKUL A,CHINRASRIO,CHANTIRATIKUL P.Effectof sodium selenite and Zinc-L-selenomethionine on performance and selenium concentrations in eggs of laying hens[J].Asian-Australasian Journal of Animal Sciences,2008,21(7):1048-1052.
[17] 刘明宝.不同硒水平对猪和大鼠的生长性能、组织硒含量、血GSH-Px活性的影响及比较[J].兽医大学学报,1989,9(1):31-36.
[18] 宋清华,田科雄.不同硒源与硒水平对肉鸡生长性能、胴体特性的影响[J].饲料工业,2009,30(24):14-16.
[19] 朱宏娟.不同硒源及硒水平对肉仔鸡生产性能和血液抗氧化指标的影响[D].硕士学位论文.长沙:湖南农业大学,2006:17-19.
[20] 崔国强,王海宏,初芹,等.不同硒源及硒水平对产蛋鸡生产性能和蛋品质的影响[J].中国畜牧兽医,2012,39(9):98-101.
[21] 罗培林,郑萍,何军,等.不同硒源及硒水平对大鼠生长性能、血清抗氧化能力和组织硒沉积的影响[J].动物营养学报,2012,24(7):1311-1319.
[22] THOMPSON JN,SCOTT M L.Role of selenium in the nutrition of the chick[J].Journal of Nutrition,1969,97(3):335-342.
[23] 石军,许金新,李咸良.微量元素硒的生理功能研究进展[J].饲料博览,2003(1):4-7.
[24] 蒋宗勇,王燕,林映才,等.硒代蛋氨酸对肥育猪血浆和组织硒含量及抗氧化能力的影响[J].中国农业科学,2010,43(10):2147-2155.
[25] 王海宏.不同硒源对肉仔鸡生产性能、组织硒含量和GSH-Px活力的影响[D].硕士学位论文.北京:中国农业科学院,1999:32-37.
[26] 林长光,林金玉,林枣友,等.不同硒源对母猪泌乳期生产性能、血浆和乳中硒含量的影响[J].中国畜牧杂志,2013,49(21):48-52.
[27] JUNIPER D T,PHIPPSR H,RAMOS-MORALESE,et al.Effect of dietary supplementation w ith seleniumenriched yeast or sodium selenite on selenium tissue distribution and meat quality in beef cattle[J].Journal of Animal Science,2008,86(11):3100-3109.
[28] SCHISLER N J,SINGH SM.Modulation of seleniumdependentglutathione peroxidase(Se-GSH-Px)activity in m ice[J].Free Radical Biology and Medicine,1988,4(3):147-153.
[29] 梁轩,刘福柱,沈伟.家禽营养与微量元素硒[J].饲料博览,2000(9):22-24.
[30] 尹俐.含硒谷胱甘肽硫转移酶的真核表达及抗氧化作用研究[D].博士学位论文.长春:吉林大学,2012:68-74.
[31] SURAIP,DVORSKA J.Is organic selenium better for animals than inorganic sources?[J].Feed M ix,2001,9(4/5):8-10.
[32] 廖一骁,胡振英,程富胜,等.酵母硒对肉鸡血清中GSH-Px、GSTs活性的影响[J].湖北农业科学,2010,49(8):1918-1920.
[33] WANG Y B,XU B H.Effect of different selenium source(sodium selenite and selenium yeast)on broiler chickens[J].Animal Feed Science and Technology,2008,144(3/4):306-314.
[34] 寇庆,梁咪娟,陶亮亮.酵母硒对肉鸡组织硒含量及抗氧化能力的影响[J].粮食与饲料工业,2012(1):48-50.
[35] CANTOR A H,MOORHEAD P D,MUSSER M A.Comparative effects of sodium selenite and selenomethionine upon nutritional muscular dystrophy,selenium-dependent glutathione peroxidase,and tissue selenium concentrations of turkey poults[J].Poultry Science,1982,61(3):478-484.
[36] 李锋,陈敏,梁新乐.酵母硒对肉鸡免疫器官及组织GSH-Px活力的影响[J].安徽农业科学,2010,38(4):1846-1848.
[37] 蒋立,吴照民,陆爱霞.有机硒对仔獭兔生长性能和组织抗氧化能力的影响[J].中国畜牧杂志,2012,48(17):27-31.
[38] CHEN G,WU J,LI C.Effect of different selenium sources on production performance and biochem ical parameters of broilers[J/OL].Journal of Animal Physiology and Animal Nutrition,2013,doi:10.1111/jpn.12136.
[39] 田金可,HUSSAIN A,李伟,等.不同硒源及水平对肉鸡组织硒含量及抗氧化功能的影响[J].动物营养学报,2012,24(6):1030-1037.
[40] 王永侠.硒代蛋氨酸对肉鸡的生物学效应及其分子机理研究[D].博士学位论文.杭州:浙江大学,2011:54-55.
[41] WANG Y X,ZHAN X A,YUAN D,et al.Influence of dietary selenomethionine supplementation on performance and selenium status of broiler breeders and their subsequentprogeny[J].Biological Trace Element Research,2011,143(3):1497-1507.
[42] YUAN D,ZHAN X,WANG Y.Effects of selenium sources and levels on reproductive performance and selenium retention in broiler breeder,egg,developing embryo,and 1-day-old chick[J].Biological Trace Element Research,2011,144(1/2/3):705-714.
[43] YOON I,WERNER T M,BUTLER J M.Effect of source and concentration of selenium on grow th performance and selenium retention in broiler chickens[J].Poultry Science,2007,86(4):727-730.
[44] 董永军,王丽荣.硒的添加形式对肉仔鸡生长性能及其肝沉积量的影响[J].中国兽医科技,2003,33(9):43-44.
[45] 杨华,傅衍,陈安国.有机硒对杜大长商品猪生产性能、胴体性状、肉质的影响[J].中国饲料,2004(6):21-24.
[46] DLOUHÁ G,ŠEVCˇÍKOVÁ S,DOKOUPILOVÁ A,et al.Effectof dietary selenium sources on grow th performance,breastmuscle selenium,glutathione peroxidase activity and oxidative stability in broilers[J].Czech Journal of Animal Science,2008,53(6):265-269.
[47] SCHRAUZER G N.Selenomethionine:a review of its nutritional significance,metabolism and toxicity[J].The Journal of Nutrition,2000,130(7):1653-1656.
[48] 丁文杰,戴晋军,杨志龙,等.富硒猪肉的研究进展[J].饲料与畜牧:新饲料,2008(7):55-57.
猜你喜欢
动物营养学报(2022年8期)2022-09-01 03:20:52
今日农业(2020年18期)2020-12-14 19:08:44
动物营养学报(2017年8期)2017-08-16 09:40:00
广东饲料(2016年3期)2016-12-01 03:43:11
兽医导刊(2016年12期)2016-05-17 03:51:44
动物营养学报(2015年9期)2016-01-07 11:29:44
少儿科学周刊·少年版(2015年11期)2015-12-17 23:47:17
少儿科学周刊·儿童版(2015年11期)2015-12-17 03:53:46
现代农业(2015年5期)2015-02-28 18:41:02
中国畜牧业(2014年4期)2014-10-16 03:50:40
