
生长在不同基底材料上的网纹藤壶底板形貌结构比较
2014-03-07 07:12彭明栋王超超
海洋科学 2014年4期
彭明栋, 曹 瑞, 唐 敏, 商 群, 王超超, 陈 新
(1. 海南大学 材料与化工学院, 海南 海口 570228; 2. 海南大学 热带岛屿资源先进材料教育部重点实验室,海南 海口 570228; 3. 中国农业大学 动物医学院, 北京 100083; 4. 海南大学 农学院, 海南 海口 570228)
生长在不同基底材料上的网纹藤壶底板形貌结构比较
彭明栋1, 曹 瑞3, 唐 敏2, 商 群1, 王超超4, 陈 新2
(1. 海南大学 材料与化工学院, 海南 海口 570228; 2. 海南大学 热带岛屿资源先进材料教育部重点实验室,海南 海口 570228; 3. 中国农业大学 动物医学院, 北京 100083; 4. 海南大学 农学院, 海南 海口 570228)
通过观察在不同材质表面上黏附的网纹藤壶底板的微观结构, 发现不同的基底材料对网纹藤壶底板结构有明显影响。在底板不同区域, 其微观结构呈现多样化, 这可能与底板处于不同形成期或与底板承担的侧壁链接、藤壶胶运输等功能有关。网纹藤壶底板结构复杂性高、可塑性强, 这为藤壶适应严酷多变的海洋环境提供了必要的结构基础。
网纹藤壶(Balanus reticulatusUtinomi); 底板; 基底材料
藤壶是一类常见的海洋污损生物, 环境适应性很强, 在世界海域广泛分布, 给各种海事活动带来严重危害, 是重点防污对象[1-2]。国内外对藤壶黏附的研究多集中在以下几个方面, 如藤壶胶的生化及理化特征分析[3-4]、腺介幼虫的初步黏附行为[5]、基底材料表面理化特征(如化学组成、疏水性、粗糙度、生物膜等)及生态因子对藤壶黏附的影响[2,6-7]等, 但对藤壶底板结构方面的研究很少[8]。藤壶通过底板黏附于基底,随着藤壶个体的不断蜕皮和生长, 底板面积一般由水平方向向四周扩大, 随底板面积的增大, 藤壶的黏附强度也不断增强[9]。此外, 藤壶胶自腺体分泌后, 主要通过毛细管作用, 经由底板上分布的通道到达底板和基底的界面[9], 可见藤壶胶的运输和分布与底板结构密切相关[10]。网纹藤壶(Balanus reticulatus)属暖水性种,适应性强, 发育快, 在热带和亚热带海域可周年繁殖,常成为一些海域污损生物群落的优势种[2,11], 在海口近岸周边海域的潮间带广为分布。本试验采集自然海域中附有网纹藤壶的竹质材料、聚丙烯腈纤维材料、普通烧结砖质材料及表面为防污涂料的钢板, 通过观察研究藤壶底板及其界面的微观结构特征, 分析了基底材料对藤壶底板的影响。期望通过藤壶底板的研究, 进一步了解藤壶在不同基底环境条件下的黏附行为及其调节机理,为防污工作提供新的思路和理论基础[12-13]。
1 实验材料和方法
1.1 实验材料
在海南海甸岛(20°1' N, 110°19' E)潮间带的自然海域(28℃; 盐度, 33‰; pH 7.8), 采集附有网纹藤壶的实验材料, 包括天然竹质材料、烧结普通砖材料、聚丙烯腈纤维材料及表面为防污涂料的钢板。将采集的各种材料带回实验室经过处理, 用于试验。
1.2 实验方法
1.2.1 试验样品制备
将附有藤壶的各种材料从海水中拿出后, 用新鲜海水清洗干净藤壶及材料表面。选正常生长的网纹藤壶作为研究对象, 其底板直径约为10~12 mm。将选取好的藤壶从其基底表面小心取下, 保证藤壶正常黏附状态时与材料表面的相对位置和结构的完整性。随后清除藤壶内部脏器, 取底板, 用流水清洗干净, 超声波清洗2次, 每次10 min, 然后用蒸馏水润洗3遍。置于80℃烘箱烘干后待用。
1.2.2 样品表面微观结构观察
干净的藤壶底板样品完全干燥后, 经过真空镀金(E-1010型离子镀膜仪), 于扫描电子显微镜(日立S-4800)下观察样品表面形貌和结构特征。
2 实验结果
本实验发现网纹藤壶能够以其底板牢固地黏附于竹质材料、烧结普通砖材料、聚丙烯腈纤维材料及表面为防污涂料的钢板四种材料表面。个体较小的藤壶较易从竹质材料表面剥离下来, 其底板较薄;但较大个体不易从各种材料表面上剥落。裸眼观察藤壶底板与四种材料的黏附表面, 发现界面界线明显, 紧密无缝隙。在此, 把藤壶底板朝向软组织的一侧称为内侧, 而黏附基底材料表面的一侧为外侧。在扫描电镜下进一步观察, 发现藤壶底板的内、外侧的微观结构各具特征。
2.1 网纹藤壶生长在烧结普通砖质材料表面
砖质材料表面黏附的网纹藤壶很难与基底分离。较低倍数下, 发现在藤壶底板边缘断面上有规律地分布着间隔大致相等的圆形空洞, 其直径约为1.0~1.5 mm。在低倍镜下, 底板外表面显得较为平整,但放大倍数较高时可见其表面形貌多样, 有的区域较为粗糙, 间或出现宽窄不一的空隙, 局部堆积着形状不规则的块状物(图 1-a); 同时, 在底板表面还分布有直径大小不一的圆形孔, 直径约在3~5 µm, 孔壁光滑, 孔内显示出其下层物质; 有些区域表面凹凸不平, 似胶状物凝固后的形成, 间杂有不规则形状的颗粒物质(图1-b)。

图1 黏附于砖质材料表面的网纹藤壶底板外侧边缘处局部表面形貌差异Fig. 1 The morphological difference between different parts of the outside edge of base plate of B. reticulatus adhering to the brick material
2.2 网纹藤壶在竹质材料表面
观察网纹藤壶底板中心部分和节间竹秆壁界面的横截面, 在低倍镜下可见在竹纤维表面上紧密附有一层较平滑的连续膜状物, 此为藤壶底板及其下的藤壶胶, 同时可见底板表面上有不规则分布的突起及空洞; 局部放大, 底板表面呈现水纹状, 且凹凸不平; 藤壶底板及其下的藤壶胶厚大约10~15 µm(图2-a)。观察因机械力而导致藤壶底板与竹秆壁分离处的断面, 可见底板的厚度不均一, 但底板中央部分质地致密均匀, 浑然一体。进一步局部放大发现底板断面较粗糙, 而且在其表面和断面上都分布有1 µm以下的孔洞(图2-b)。
观察网纹藤壶底板靠近边缘处的内侧, 可见其表面由大小不一、无规则形状的块状物堆砌嵌合呈凹凸不平状(图2-b)。

图2-a 黏附于竹秆壁表面的网纹藤壶底板横截面Fig. 2-a The cross-section of the base plate of B. reticulatus adhering to bamboo material
2.3 网纹藤壶生长在聚丙烯腈纤维材料表面
网纹藤壶分泌的藤壶胶将底板紧密地黏附于聚丙烯腈纤维材料表面, 将纤维艰难地从底板剥离后,在扫描电镜下观察。较低倍数下可见藤壶底板与纤维的接触面凹凸不平, 在中心部位有一类似圆形的孔洞, 直径约2 mm。较高倍数下观察发现, 膜状平整区域是层状物覆在细密颗粒表面上形成的, 推测膜状平整区域可能是藤壶胶覆盖区(图 3-a), 其下底板表面为类似石钟乳突起的大小不一的块状物或颗粒状密集堆叠而成(图 3-b), 颗粒之间可见分布有大小不一、形状不规则的空洞开口。

图 2-b 黏附于竹秆壁表面的网纹藤壶底板内侧边缘处局部表面形貌Fig. 2-b The morphology of the outside edge of base plate of B. reticulatus adhering to bamboo material
2.4 网纹藤壶生长在防污涂层表面
在防污涂料表面附着的藤壶较难剥离, 在显微镜下可见接触防污漆的底板在边缘呈明显的分层现象, 即在两层底板物质之间夹有一层较为疏松的黑褐色物质, 呈现独特的“三明治”层状结构(图 4)。进一步观察底板的超微结构, 发现各层的形态结构差别较大。外层板壁表面呈现出层片和颗粒物堆积的不规则排列形貌, 其间分布有开口大小不一的间隙(图 5-a); 中间黑褐色物质存在的部分呈疏松蜂窝状结构, 紧接着的下层按一定规则排列成较密集的网络结构(图5-b)。
3 讨论
3.1 藤壶底板不同区域的表面形貌差异

图3 生长在聚丙烯腈纤维表面的网纹藤壶底板外侧形貌结构Fig. 3 The morphology of the outside edge of base plate of B. reticulatus adhering to the surface of Polyacrylonitrile fiber

图4 生长在防污涂料上网纹藤壶底板形貌结构, 显示分层特征Fig. 4 The base plate of B. reticulatus adhering on the antifouling paint, showing the laminated structure
正常生长的网纹藤壶(Amphibalanus reticulatus)底板为坚硬的生物钙质化结构[9], 微观构造复杂, 在底板不同区域, 底板内外侧及内部微观结构特征都呈现出明显差异。
生长在竹质材料表面的藤壶底板内侧, 靠近中央区域的底板致密, 浑然一体, 而靠近边缘处的底板则呈现堆积的块状结构。藤壶自腺介幼虫黏附于基底营固着生活开始, 其底板随藤壶的生长而不断向四周扩延[9]。因此, 在藤壶附着后的生长发育阶段,藤壶底板围绕着其初始黏附点, 一直处于一个不断更新和扩建的过程, 从藤壶底板不同区域的微观结构差异可反映出这个特征。不断扩大的底板和积累的藤壶胶使藤壶的黏附强度随藤壶生长时间的增加而加强。故推测, 边缘处的底板可能因为形成期较短, 堆积块状物在结构上还未形成如中央部分的致密整体结构。
从横截面上进一步观察藤壶底板内部结构的特征, 在底板边缘处的横截面上明显可见分布较规则的孔腔结构, 其截面呈椭圆形, 500 µm左右, 长径与边缘平行, 这与Sangeetha 和Ravi[8]所观察到的情况非常类似, 只是此处在砖质材料表面观察到的孔间距较大。推测底板边缘出现的这些空腔结构一方面可能加强底板与侧板的链接, 另一方面可能也参与或辅助藤壶胶的运输和分布[8]。而在藤壶底板中央部分横截面上分布的只有直径约5 µm的圆形空腔孔洞,研究发现藤壶钙质底板分布有用以运输藤壶胶质的微导管[6], 故推测藤壶胶从分泌腺分泌后, 底板的孔腔结构可能作为运输通道, 使藤壶胶在一定的时间内可顺利到达底板与基底的空隙。
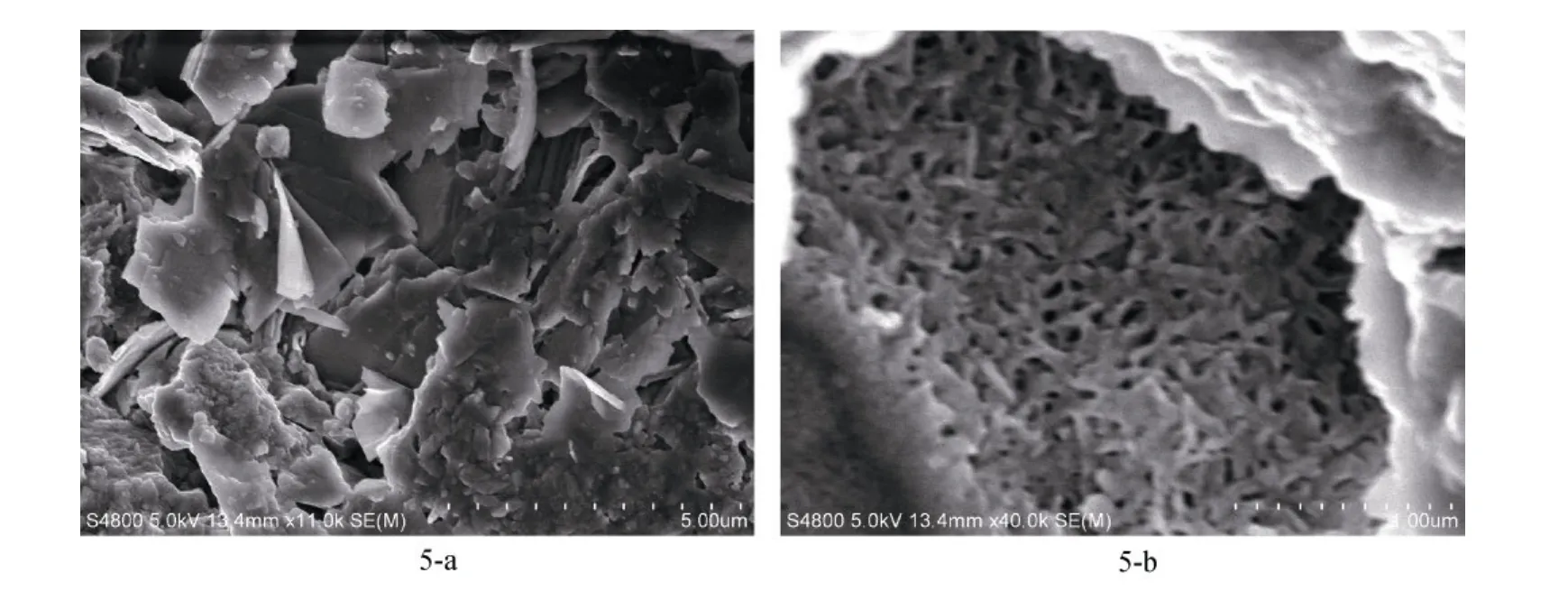
图5 生长在防污涂料表面的网纹藤壶底板外侧形貌结构Fig. 5 The morphology of the outside edge of base plate of B. reticulatus adhering to the surface of the antifouling paint
3.2 黏附于不同基底材料的藤壶底板表面形貌差异
通过观察实验中四种基底材料上附着藤壶底板,发现在不同的基底材料表面上底板的微观形貌结构差异较大。
竹质材料表面黏附生长的藤壶底板较平整, 聚丙烯腈纤维上的藤壶底板凹凸不平, 沟壑交纵。显然,聚丙烯腈纤维非常不规整的表面作为藤壶黏附基底,直接导致了不规则的底板结构形成, 生长在聚丙烯腈纤维表面的藤壶底板外侧表面形貌呈块状和细密颗粒结构, 底板和黏附基底之间的藤壶胶表面亦凹凸不平。此外, 底板与基底材料的结构在界面高度吻合, 可推断藤壶底板在形成初期具有一定的可塑性,藤壶胶在抵达目的地固化前流动性和渗透性较强,因此藤壶胶能较充分地渗入一些多孔基底材料表面以下的微细空腔结构中, 有利于增加藤壶底板与黏附基底表面的结合强度。
比较在砖质和竹质材料表面生长的藤壶底板,尽管其内侧都分布有用以输导藤壶胶的圆形孔腔,但孔径大小相差约一个数量级。推测在底板形成时,砖质材料表面粗糙多孔的特征影响了底板的微观结构, 致使其微导管的直径变大, 以输送较多藤壶胶于多孔的黏附基底, 加强底板在基底表面的黏附强度。
表面观察正常生长在无防污涂料表面的藤壶底板,可见其整体为较均一、不分层的致密钙化结构。而生长在防污涂层表面的网纹藤壶底板呈现出的独特的“三明治”层状结构。推测这种差异是由于在腺介幼虫黏附初期和藤壶生长过程中, 受到涂料中防污活性物质的影响,与涂料接触的底板在形成和发育过程中结构出现了异常变化, 体现在底板的微观形貌与正常藤壶底板有明显的结构差异。其影响机理还需进一步探索。
显然, 为适应复杂的外部生存环境, 网纹藤壶的腺介幼虫在选择了一定特征的材料作为黏附基底后,不同表面特征的基底材料会影响藤壶底板的形貌结构。基底对藤壶底板的形成及其藤壶胶的分泌运输有较强的调节作用。要认识基底材料对藤壶黏附及其生长的影响因素及其机理, 为防污材料的研发提供更有价值的参考资料, 还需今后进一步的深入研究。
[1] 黄宗国, 蔡如星. 海洋污损生物及其防除[M]. 北京:海洋出版社, 1984.
[2] 黄宗国. 海洋污损生物及其防除[M]. 北京: 海洋出版社, 2008.
[3] Kamino K, Inoue K, Maruyama T. Barnacle Cement Proteins. Importance of disulfide bonds in their insolubility [J]. The Journal of Biological Chemistry, 2000, 275 (35): 27360-27365.
[4] Sullan, R M A, Gunari N, Tanur A E, et al. Nanoscale structures and mechanics of barnacle cement [J]. Biofouling, 2009, 25 (3): 263-275.
[5] Phang Y, Aldred N, Clare A S, et al. Towards a nanomechanical basis for temporary adhesion in barnacle cyprids (Semibalanus balanoides) [J]. Journal of the Royal Society Interface, 2008, 5: 397-401.
[6] Lindner, E. The attachment of macrofouling invertebrates[C]//Annapolis MD. Marine Biodeterioration: An Interdisciplinary Study. US Naval Institute: Costlow, J.D., Tipper, R.C, 1984: 183-201.
[7] 张明明, 赵文, 于世超, 等. 我国海洋污损生物的研究概况[J].水产科学, 2008, 27 (10): 545-549.
[8] Sangeetha R, Ravi K. Construction and nanomechanical properties of the exoskeleton of the barnacle,Amphibalanus reticulates[J]. Journal of Structural Biology, 2011, 176: 360-369.
[9] Wiegemann M. Structure formation in barnacle adhesive[C]//Possart W, Wiegemann M. Adhesio: Current Research and Application. Weinheim: Wiley-VCH Verlag GmbH &Co. KGaA, 2006: 143-156.
[10] Kamina K. Barnacle attachment. [C]//Smith A M, Callow J A. Biological Adhesives. Springer-Verlag Berlin and Heidelberg GmbH & Co. K., 2006: 146-166.
[11] 方芳, 严涛. 南海污损生物的研究现状及展望[J]. 热带海洋学报, 2004, 23(1): 76-85.
[12] 周文木, 王孝杰, 胡碧茹, 等. 海洋污损生物黏附机制及防污涂层表面工程[J]. 应用化学, 2010, 27(9): 993-997.
[13] Raman S, Karunamoorthy L, Doble M, et al. Barnacle adhesion on natural and synthetic substrates: Adhesive structure and composition[J]. International Journal of Adhesion & Adhesives, 2013, 41: 140-143.
(本文编辑: 康亦兼)
Comparison of the base plate structure of barnacle adhered on different substrates
PENG Ming-dong1, CAO Rui3, TANG Min2, SHANG Qun1, WANG Chao-chao4, CHEN Xin2
(1. Faculty of Material and Chemical Engineering, Hainan University 570228, China; 2. Key Lab of Advanced Materials of Tropical Island Resources, Ministry of Education, Hainan University 570228, China; 3. College of Veterinary Medicial, China Agricultural University 100083, China; 4. College of Agriculture, Hainan University 570228, China)
May, 16, 2013
Balanus reticulatusUtinomi; base plate; substrate
In this paper, the microstructure of base plates ofBalanus reticulatuswhich adhered to different materials was studied. It was found that the microstructure of base plates showed different appearances on different substrates. So it is speculated that the substrate has a significant impact on the base plate structure. In addition, different texture features between the edge and center part of the same base plate was also found, which may be related to the function of base plate at different developing stage, such as the stabilization of lateral plates, and the transportion of barnacle cement. The results of this study shed light on the complexity and plasticity of barnacle base plates, which may provide evidence to the adaption of barnacles to the adverse environment variations.
P745
A
1000-3096(2014)04-0034-05
10.11759/hykx20130516001
2013-05-16;
2013-10-16
国家自然科学基金项目(31160098); 海南省自然科学基金项目(313104); 海南大学博士科研启动基金项目(kyqd1046); 海南省高等学校科学研究项目(hjjg2013-05); 海南大学教育教学研究项目(hdjy1208)
彭明栋(1992-), 男, 本科生, 山东滕州人, 研究方向: 生物污损, E-mail: 23543920@qq.com; 陈新, 通信作者, 讲师, 硕士,主要从事海洋污损方面的研究, E-mail: chenxincx@tom.com
猜你喜欢
食品研究与开发(2022年16期)2022-08-24
军事文摘(2021年22期)2022-01-18
大科技·百科新说(2021年5期)2021-07-27
今日农业(2020年14期)2020-08-14
热带生物学报(2019年3期)2019-07-29
校园英语·中旬(2018年11期)2018-12-07
新教育(2018年11期)2018-02-26
延河(下半月)(2015年4期)2015-09-24
延河·绿色文学(2015年4期)2015-06-01
中国校外教育(2010年8期)2010-08-15
