
贺兰山牦牛不同时间段的生境选择1)
2014-03-06 03:19刘振生孙兆惠李路云赵宠南李艳香
东北林业大学学报 2014年4期
刘振生 孙兆惠 李路云 赵宠南 李艳香 王 越
(国家林业局野生动物保护生物学重点实验室(东北林业大学),哈尔滨,150040)
牦牛(Bos grunnieus)别名野牛,属偶蹄目(Artiodactyla)牛科(Bovidae),单型种,无亚种分化,是高原地区的特有物种[1],在我国主要分布于西藏、青海、四川、甘肃、宁夏和内蒙古等地[2]。由于人为过度猎杀,其种群数量在过去的30 a 间锐减了30%,被列为国家Ⅰ级重点保护野生动物[3]。近年来,国内外的研究,多针对分布在青藏高原的牦牛,主要集中在牦牛分布状况及资源利用[2,4-5]、种群数量[6-11]和牦牛对改良家畜的影响[12-15]等方面。贺兰山地区的牦牛属于野化种群,已生存了几百年;其种群数量的扩增,在一定程度上说明牦牛已适应了贺兰山当地的环境。但其生态学研究的资料还很匮乏,仅有姚志诚等[16]、董嘉鹏等[17]、赵宠南等[18]和刘振生[19]对牦牛的形态特征、生态特征和该种群的冬春季食性、行为及生境选择进行了报道。因此,对贺兰山地区牦牛的不同时间段生境选择进行调查研究,不仅可以完善其生态学信息,还可探讨其与贺兰山同域分布的其他有蹄类动物之间的关系。
1 研究区概况
贺兰山坐落在宁夏回族自治区和内蒙古自治区交界处,位于银川平原与阿拉善高原之间,其地理坐标为北纬38°21'~39°22'、东经105°49'~106°42',为宁夏贺兰山国家级自然保护区和内蒙古贺兰山国家级自然保护区。属中温带干旱气候区,夏季炎热而短暂,秋季多风而凉爽。无霜期短,年平均气温为-0.7 ℃,6—9月份平均降水量达260.2 mm,占全年降水量的62%。贺兰山具有独特的地形、地貌及地理特征,是我国北方地区唯一的生物多样性中心。按植被型,可划分成4 个植被垂直带:山前荒漠与荒漠草原带;山麓与低山草原、灌丛带;中山针叶林带;高山或亚高山灌丛、草甸带。据统计,目前贺兰山牦牛的种群数量为400 ~500 只,集中分布在哈拉乌沟[19]。
2 研究方法
2009年7—8月份和9—11月份,根据贺兰山各个沟段牦牛数量及分布,在牦牛活动较集中沟段(北寺沟、哈拉乌沟、后沟、强岗岭沟、镇木关沟等)设置了30 条样线。样线覆盖贺兰山所有植被类型。
沿样线行走,发现牦牛的足迹、卧迹、食痕、粪便、尿迹和残留的毛发后,以此选择痕迹为中心设置一个10 m×10 m 样方,在该样方中心及4 个角各设置一个1 m×1 m 小样方;记录大样方内的18 种生态因子,包括海拔高度,植被类型、地形特征、优势乔木、乔木密度、乔木高度、牦牛距乔木的距离、灌木种类、灌木密度、灌木高度、牦牛距灌木的距离、植被盖度、坡度、坡向、坡位、距水源距离、距人为干扰距离和隐蔽级。具体测定方法和等级划分标准,参见文献[8]。7—8月份测量了101 个选择样方,9—11月份测量了93 个选择样方。同时,在样带上利用GPS 每隔1 000 m 设置一个对照样方,其设定方法和测定内容同利用样方一样,7—8月份测量了115个对照样方,9—11月份测量了108 个对照样方。
本研究使用拟合优度卡方检验牦牛对18 种生态因子是否有选择性,再用Bonferroni 不等式分析牦牛对所设定生境因子中的哪些种类偏好或回避。然后基于Marcum 法[20]对生境选择和的描述,使用卡方检验对比分析贺兰山牦牛对不同时间段生境的18 种生态因子的选择存在的差异性。在主成分分析中,取特征值大于1 的成分作为主成分。主要对牦牛不同时间段选择生境的18 中生态因子进行主成分分析,确定主要因子。
对牦牛选择生境中的植被类型、坡位、地形特征、优势乔木、坡向、灌木种类6 种非数值型生态因子进行卡方检验。当χ2值越大时,说明不同时间段选择生境中的生态因子差异性显著。当P<0.01 时,差异极显著;P<0.05 时,差异显著;P>0.05 时,差异不显著[21]。对牦牛选择生境样方内的海拔高度、乔木密度、乔木高度、牦牛距乔木的距离、灌木密度、灌木高度、牦牛距灌木的距离、植被盖度、坡度、距水源距离、距人为干扰距离和隐蔽级共12 种数值性生态因子进行U 检验。当P<0.05 时,差异显著;P>0.05时,差异不显著。
3 结果与分析
3.1 牦牛7—8月份生境选择特征
3.1.1 选择生态因子特征
在7—8月份,贺兰山牦牛对14 种生态因子有选择性。牦牛偏好选择的生境:海拔高度>3 000 m、植被类型为亚高山灌丛和草甸带、乔木密度<100株/hm2、乔木高度<3 m、牦牛距乔木的距离>3 m、灌木种类是混合型灌木、灌木密度>2 000 丛/hm2、牦牛距灌木的距离<1 m、植被盖度>70%、上坡位、距水源距离<800 m 或800 ~1 200 m 之间的距离、距人为干扰距离>4 000 m、隐蔽级>70%(表1)。
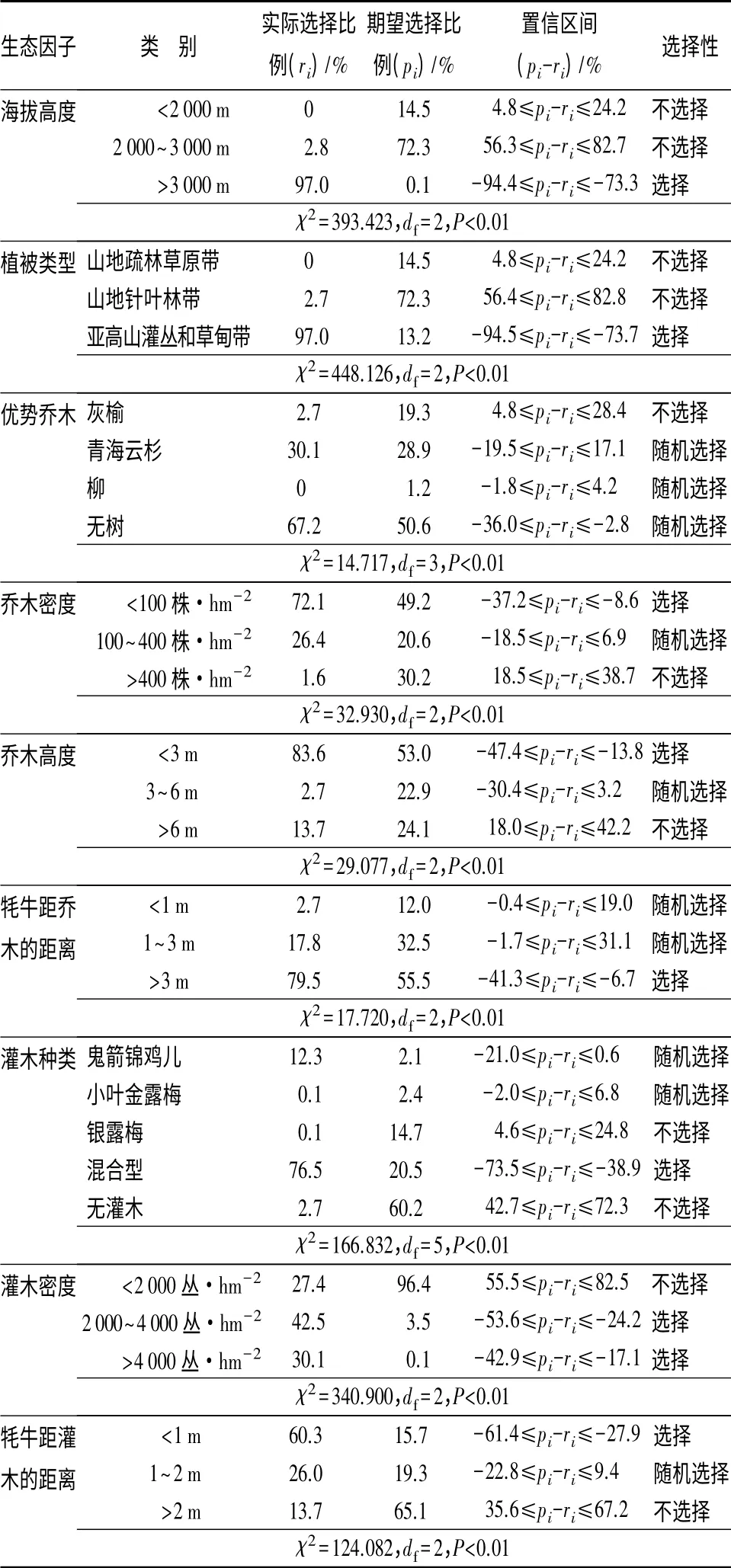
表1 牦牛7—8月份对14 种生态因子的选择性

续(表1)
3.1.2 对选择生态因子的主成分分析
贺兰山牦牛7—8月份选择生境的主成分分析结果表明,牦牛7—8月份选择生境的前4 个成分的特征值大于1,累积贡献率为62.734%,可以较好的反映出野化牦牛7—8月份选择生境的特征(表2)。在第一主成分里,植被类型、优势乔木、牦牛距乔木的距离、隐蔽级、距水源的距离的相关系数较高,对第一主成分的贡献大,绝对值较大的相关系数出现在植被类型、海拔高度、优势乔木、隐蔽级、距水源距离上,说明这几个生态因子有较大的信息载荷量;第二主成分里,地形特征、坡度和灌木种类相关系数较高,对第二主成分的贡献大,绝对值较大的相关系数出现在坡位、坡度、地形特征、灌木种类上,说明这几个生态因子有较大的信息载荷量。第三主成分里,海拔高度、距水源的距离、植被类型的相关系数较高,对第三主成分贡献大,绝对值较高的相关系数出现在海拔高度、距水源的距离、植被类型上,说明这几个生态因子有较高的信息载荷量。
3.2 牦牛9—11月份生境选择特征
3.2.1 选择生态因子特征
在9—11月份,贺兰山牦牛对16 种生态因子有选择性,牦牛偏好选择的生境:海拔高度2 000~3 000 m、植被类型为山地针叶林带、乔木密度<100 株/hm2、乔木高度<3 m、灌木种类是混合型灌木或者无灌木、灌木密度<2 000 丛/hm2、牦牛距灌木的距离>2 m、植被盖度>50%、下坡位阳坡、距水源距离>1 200 m、坡度<10°、隐蔽级>70%(表3)。
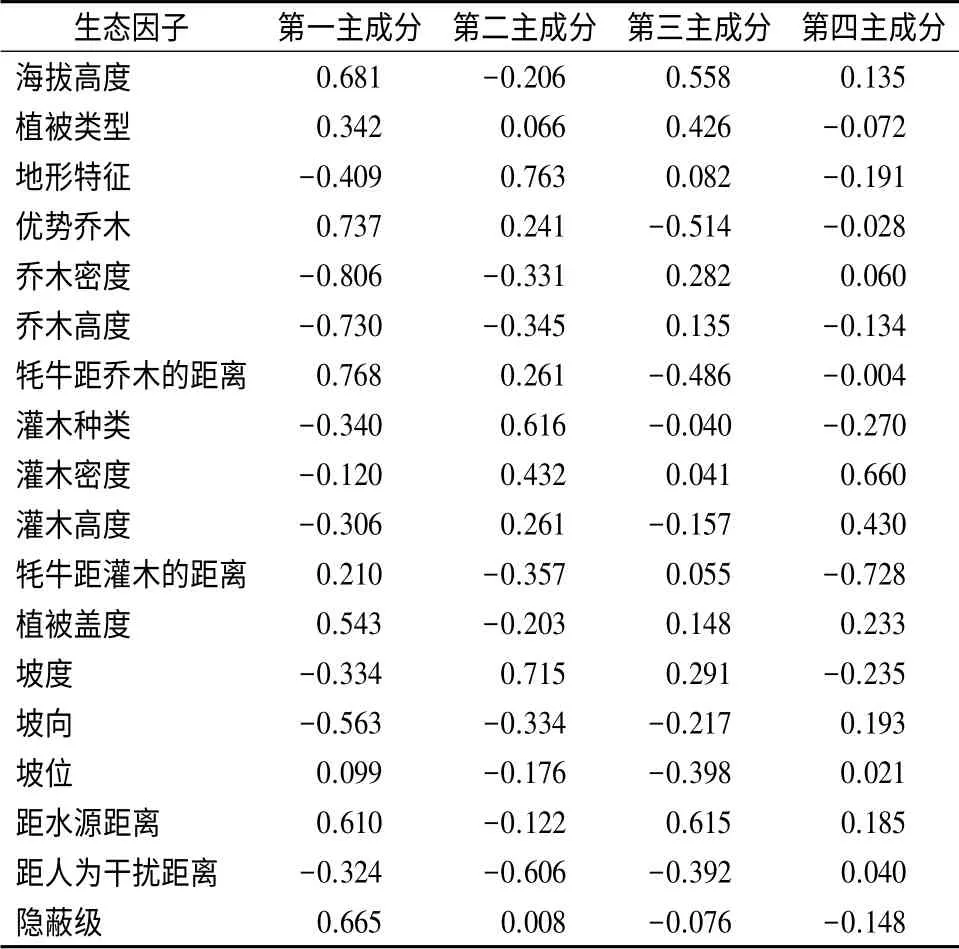
表2 牦牛7—8月份选择的18 种生态因子的主成分分析

表3 牦牛9—11月份对16 种生态因子的选择性
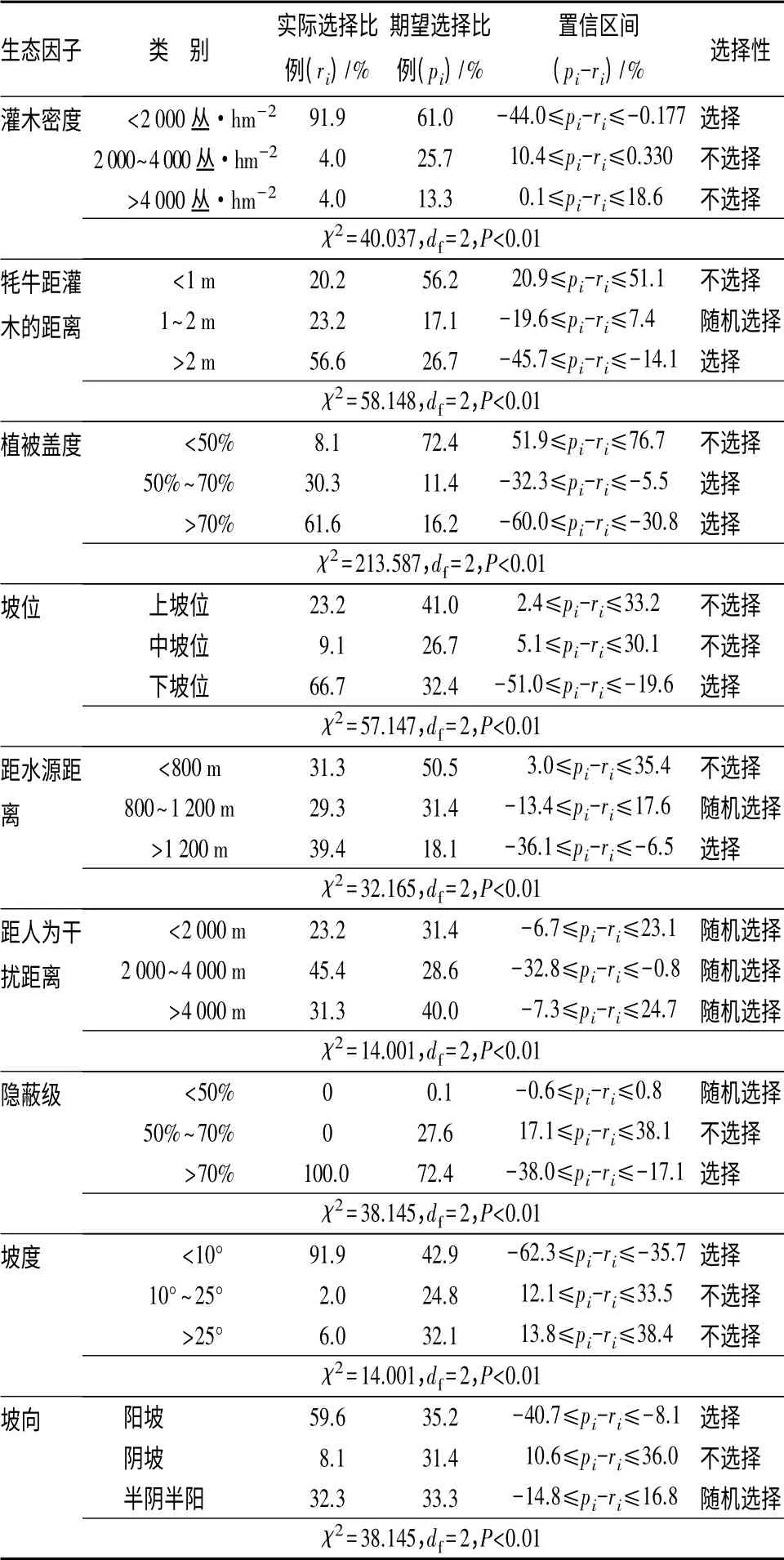
续(表3)
3.2.2 对选择生态因子的主成分分析
贺兰山牦牛9—11月份选择生境的主成分分析结果表明,牦牛9—11月份选择生境的前4 个成分的特征值大于1,累积贡献率为72.795%,可以较好的反映出野化牦牛9—11月份选择生境的特征(见表4)。在第一主成分里,乔木高度的相关系数较高,对第一主成分的贡献大,其中绝对值较大的相关系数出现在乔木高度和乔木密度上,说明这2 个生态因子有较大的信息载荷量;第二主成分里,地形特征、坡度和灌木密度相关系数较高,对第二主成分的贡献大,绝对值较大的相关系数出现在坡位、坡度、地形特征、坡向上,说明这几个生态因子有较大的信息载荷量。第三主成分里,海拔高度、植被类型的相关系数较高,对第三主成分贡献大,绝对值较高的相关系数出现在海拔高度、植被类型上,说明这2 个生态因子有较高的信息载荷量。

表4 牦牛9—11月份选择的18 种生态因子的主成分分析
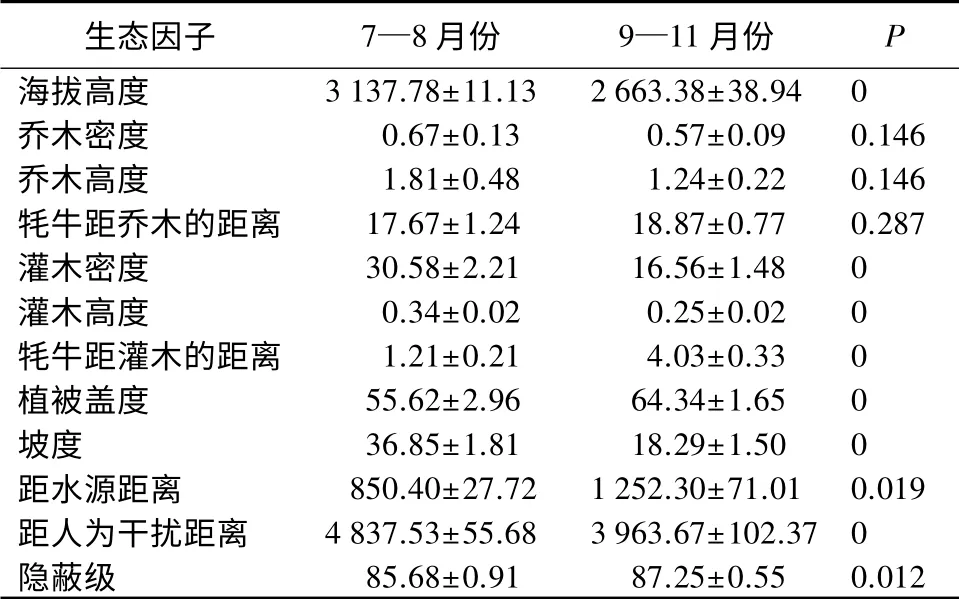
表5 牦牛不同时间段选择生境的12 种生态因子比较
3.3 牦牛不同时间段选择生境生态因子的比较
卡方检验结果显示:贺兰山牦牛不同时间段选择的生境,在植被类型(χ2=37.90,df=4,P<0.01)、地形特征(χ2=229.79,df=4,P<0.01)、优势乔木(χ2=123.83,df=2,P<0.01)、灌木种类(χ2=144.21,df=5,P<0.01)、坡位(χ2=64.38,df=2,P<0.01)、坡向(χ2=119.90,df=2,P<0.01)上,差异极显著。Mann-whitey U 检验显示:不同时间段,贺兰山牦牛在海拔高度、灌木密度、灌木高度、牦牛距灌木的距离、植被盖度、坡度、距人为干扰距离上,差异显著(P<0.05)。与7—8月份相比,9—11月份,牦牛偏好选择海拔低、灌木密度小、灌木高度低和距离远、植被盖度大、坡度小、距人为干扰距离近的生境。
4 讨论
动物选择不同的生境类型,反映出自身的生理需求和捕食风险之间的一种平衡关系;在不同的时间段选择不同的生境,也是为了自身获得更大的收益。通过对比贺兰山牦牛不同时间段选择的生态因子可以看出:7—8月份,牦牛主要在海拔3 000 m 以上的亚高山灌丛和草甸带活动;9—11月份,迁移到海拔高度为2 000~3 000 m 的山地针叶林带中。这表明,贺兰山牦牛在不同的时间段选择不同的生态因子,以此寻找适合自身生理特性的生境[7-8]。
贺兰山属于典型的大陆性气候,9—11月份增温迅速,降水量少,天气炎热。这样的气候特点对耐寒怕热的牦牛来说,是一个挑战。为了躲避高温,牦牛尽可能的向海拔高的区域迁移。贺兰山地区,海拔相对较低,因此牦牛只能在3 000 m 以上区域活动,在海拔3 100 m 左右的区域风速大且风口多,天气凉爽,既可避免高温这种不利条件的影响,也可躲避人为干扰。由于牦牛体型较大,乔木密度高会影响其运动,因此选择乔木密度低、灌木种类为混合型灌木和植被盖度大的生境。由于高寒草原或荒凉贫瘠的寒漠地带的空气稀薄、植被贫乏,加之牦牛食量大,所以牦牛大部分时间都用来摄食,因此食物资源分布对牦牛的迁移有着重要作用。7—8月份,亚高山灌丛和草甸带的食物丰富,主要包括珠芽蓼(Polygonum viviparum)、内蒙葶苈(Draba mongolica)、高山柳(Salix cupularis)、针茅(Stipa breviflora)和禾叶风毛菊(Saussurea graminea)等[16]。7—8月份温度高,热量散失快,加剧了牦牛对水源的需求,在气温较高的中午,牦牛会到湖泊或溪流中沐浴吃草,那里的干旱草原植物的水分含量高,而且品种众多。
9—11月份,牦牛偏好选择海拔2 000~3 000 m、阳坡(阳坡的灌木和草本的长势比阴坡的好)、植被类型为山地针叶林带、乔木密度小且高度低、灌木种类和植被盖度丰富的地方活动。在贺兰山地区,9—11月份气温变化大,10月份早霜就已到来,有时还伴有小雪,很快进入冬季,在高海拔地区灌木和草本植物已开始枯萎,食物质量也随之下降。同时,牦牛也没有刨雪取食的本领。因此,食物数量和质量的下降,开始影响牦牛的生存,所以,牦牛要迁移到低海拔地区。在中等海拔地区的山地针叶林带,开阔区域中的草本和灌木数量多,食物质量好。虽然该区域距离水源较远,但牦牛在9—11月份不需要水来降温,对水源的需求没有7—8月份多。牦牛在冬季来临之前,需要通过充足的采食来增加自身脂肪的储备,以备过冬,因此,为了寻求食物,是牦牛在秋季迁移的主要原因。
贺兰山已没有大型食肉动物的存在[22],赤狐(Vulpes vulpes)也只是对幼体构成一定的威胁,因此,天敌对牦牛没有太大的影响。与贺兰山牦牛同域分布的有蹄类食草动物,还有岩羊(Pseudois nayaur)、马鹿(Cervus elaphus)和高山麝(Moschus chrysogaster)[16]。7—11月份,牦牛在高海拔地区的亚高山灌丛和草甸带以及山地针叶林带活动,活动范围大,且食物丰富度高;因此,种内竞争对牦牛影响不大。与牦牛同域分布的岩羊、马鹿和高山麝,7—11月份则主要分布于山地针叶林带的密林中和山地疏林草原带,因此,种间竞争也较小。放牧,对贺兰山野生动物有明显的负作用[23],家畜是牦牛食物和栖息地的潜在竞争者。因此,远离人为干扰,是牦牛生存的重要策略。在青藏高原,牧民和家畜占据了广阔的高山草甸和高山干旱草原,使得牦牛只能栖息在无人居住的荒漠草原[24]。
[1] 刘务林.浅谈野牦牛的起源与现状[J].西藏大学学报,1999,22(1):114-116.
[2] 张知贵,夏霖,杨奇森.牦牛的分布及保护[J].动物学杂志,2009,44(1):148-150.
[3] 汪松.中国濒危动物红皮书:兽类[M].北京:科学出版社,1998:295-299.
[4] 刘务林,Schaller G.野牦牛的分布和现状[J].西藏科技,2003(11):17-23.
[5] 陆仲璘.中国的牦牛资源[J].中国草食动物,1999,2(1):42-46.
[6] 焦增华,杨孝朴,曾玉峰.高原之王:野牦牛[J].中国畜禽种业,2009,5(4):41-42.
[7] 常明华,郎侠.野牦牛行为及其环境关系的研究[J].中国牛业科学,2009,35(3):23-24.
[8] 马宗祥,窦耀宗.中国牦牛产区的生态特点和牦牛的生物学特性[M].成都:四川科学技术出版社,1989:21-35.
[9] 李孔亮,陆仲璘.牦牛科学论文集[M].兰州:甘肃民族出版社,1990:78-82.
[10] 朴仁珠,马逸清,崔花淑.中国野牦牛现状研究[J].生命科学研究,1999,3(2):161-169.
[11] 尹秉高,刘务林.西藏珍稀野生动物与保护[M].北京:中国林业出版社,1993:1-54,91.
[12] 尚有安,杨安圈,王照忠,等.野牦牛改良家牦牛的生产性能效果试验研究[J].中国牛业科学,2011,37(5):31-32.
[13] 阎萍,潘和平.野牦牛种质特性的研究与利用[J].中国畜牧杂志,2004,40(12):31-33.
[14] 张发慧.肃南牦牛导入野牦牛杂交改良试验观察[J].中国畜牧兽医,2005,32(7):25-26.
[15] 闫萍,陆仲璘,王敏强,等.野牦牛种用价值的研究和利用[J].中国草食动物,2005(z2):247-249.
[16] 姚志诚,刘振生,王兆锭,等.贺兰山野化牦牛冬春季食性[J].生态学报,2011,31(3):673-679.
[17] 董嘉鹏,刘振生,滕丽微,等.贺兰山野化牦牛秋季昼间行为时间分配和活动规律[J].经济动物学报,2013,17(2):71-74,77.
[18] 赵宠南,苏云,刘振生,等.贺兰山牦牛冬春季的生境选择[J].生态学报,2012,32(6):1762-1772.
[19] 刘振生.宁夏贺兰山脊椎动物志[M].银川:宁夏人民出版社,2009.
[20] Marcum C L,Loftsguarden D O.A nonmapping technique for studying habitat preferences[J].Journal of Wildlife Management,1980,44(4):963-968.
[21] Sachot S,Perrin N,Neet C.Winter habitat selection by two sympatric forest grouse in western Switzerland:Implications for conservation[J].Biological Conservation,2003,112(3):373-382.
[22] Wang X M,Schaller G.Status of large mammals in Inner Mongolia,China[J].华东师范大学学报:特刊哺乳动物,1996(6):93-104.
[23] 刘志霄,李元广,于海,等.干旱与放牧对贺兰山野生有蹄类影响的初步观察[J].华东师范大学学报:自然科学版,1997,7(3):107-109.
[24] 姚军,杨博辉,闫萍,等.中国野牦牛栖息地环境及种群行为分析[J].草业学报,2006,15(2):124-128.
猜你喜欢
建筑与预算(2022年6期)2022-07-08
天津农林科技(2022年2期)2022-04-19
文萃报·周二版(2021年5期)2021-03-02
农村实用技术(2020年1期)2020-12-16
金秋(2019年24期)2019-06-17
乡村科技(2018年8期)2018-02-13
现代园艺(2018年14期)2018-01-18
诗潮(2017年2期)2017-03-16
湖南林业科技(2017年1期)2017-02-06
军事体育学报(2014年4期)2014-02-27
