
利用插入/缺失标记分析我国瓠瓜资源遗传多样性
2014-02-28 01:55鲁忠富吴晓花胡耀文汪宝根吴新义李国景
浙江农业科学 2014年5期
鲁忠富,王 莎,吴晓花,胡耀文,汪宝根,吴新义,徐 沛,李国景
(浙江省农业科学院蔬菜研究所,浙江杭州 310021)
利用插入/缺失标记分析我国瓠瓜资源遗传多样性
鲁忠富,王 莎,吴晓花,胡耀文,汪宝根,吴新义,徐 沛,李国景
(浙江省农业科学院蔬菜研究所,浙江杭州 310021)
瓠瓜作为重要的葫芦科作物之一,在我国已有悠久的栽培历史。瓠瓜分子标记研究大大落后于黄瓜、白菜等作物,应用分子标记技术开展瓠瓜种质资源的遗传多样性和亲缘关系研究,对促进瓠瓜重要种质资源的开发和利用具有重要意义。本研究选择96份具有广泛代表性的我国瓠瓜种质材料,利用插入/缺失分子标记技术,研究分析了其遗传多样性和亲缘关系,探讨我国瓠瓜品种间的基因差异和瓠瓜亚种间及亚种内的亲缘关系。结果表明,瓠瓜种质遗传相似系数0.30~1.00,在遗传相似系数0.68处可将96份瓠瓜种质材料分为8类,各种质材料分布较平均;相同瓜型的材料有聚集在一起的趋势,根据瓜型可将这96份瓠瓜种质大致分为3类。
瓠瓜;聚类分析;插入/缺失标记;遗传多样性
瓠瓜(Lagenaria siceraia(Molina)Standl.)为雌雄同株异花一年生草本植物(2n=2χ=22),隶属葫芦科(Cucurbitaceae)葫芦属(Lage naria),是世界上重要的葫芦科作物之一[1]。瓠瓜原产赤道非洲南部低地[2],是人类最早种植的作物之一,1万年前就有考古学上的证据表明人类已开始种植瓠瓜。我国瓠瓜遗传资源丰富,其栽培史可上溯到新石器时代,被认为是瓠瓜的次生起源中心之一[3]。瓠瓜在我国主要分布在长江流域及以南地区,近年来,山东、河北、河南等地也有栽培[4],可作为瓜果蔬菜食用,既可烧汤做菜,又可腌制晒干;其成熟果还可用作容器或乐器;此外,瓠瓜可用作嫁接西瓜的砧木,用来预防西瓜枯萎病[3,5]。
目前瓠瓜分类主要根据种质资源的表型特征,按形态分,有圆柱形、牛腿形、手榴弹形等;按大小分,老瓜大的可达8~10 kg,小的不足0.1 kg;按嫩瓜颜色分,有深绿、浅绿、白色、白底带花青斑等;按品质分有柔滑型、硬肉型(硬于黄瓜);按口味分,有淡味型、甜味型、清苦型(类似苦瓜之苦味)、剧苦型;按生育期可分为早熟型、中熟型、晚熟型。此外,不同类型、不同品种的长势强弱、抗病性、抗逆性等也都有差异[6-7]。但目前在DNA水平上检测瓠瓜不同种质间遗传多样性的研究还较少。
我国瓠瓜生产上以瓠子类品种为主,种质来源局限性大,缺乏遗传多样性。近年来瓠瓜生产中病毒病、白粉病、枯萎病等病害多发,苦味瓜不时产生,商品瓜性状多样性跟不上消费者需求等问题日益严重,亟需拓宽栽培瓠瓜的遗传多样性,以培养新一代良种,为此有必要深入了解我国现有瓠瓜品种的遗传多样性和亲缘关系。
1 材料和方法
1.1 材料
试验材料为浙江省农业科学院蔬菜所瓠瓜豇豆组历年搜集并具有广泛代性的96份地方品种资源所组成的自然群体。代号南秀为L S1,孝感瓠子为L S2,贵州圆瓠瓜为L S3,长瓠为L S4,龙岩长四月瓠为L S5,长蒲1为L S6,甬砧1号为L S7,新疆瓠瓜为L S8,东阳农家种1为L S9,白葫芦为L S10,萧山园蒲为L S11,广东早瓠瓜为L S12,松桃瓠瓜为L S13,金钱蒲为L S14,全化蒲瓜为L S15,东阳农家种2为L S16,四川长瓠
瓜为L S17,临夏瓠子为L S18,长蒲2为L S19,肖长蒲为L S20,湖南圆瓠瓜为L S21,小籽瓠瓜为L S22,长蒲3为L S23,安吉地蒲为L S24,武都瓠子为L S25,清秀为L S26,湖南瓠瓜为L S27,台江瓠瓜为L S28,津胜特选为L S29,青砧为L S30,辽宁瓠子为L S31,蒲县瓠瓜为L S32,新蔡瓠瓜为L S33,湖南圆瓠1号为L S34,G 22②①为L S35,绍兴长瓜为L S36,短筒瓠子为L S37, G 32②②为L S38,杭州长瓜为L S40,云南农家种为L S41,麦瓠为L S42,小葫芦为L S43,新选为L S44,汉龙青玉为L S45,长蒲4为L S46,极早熟瓠瓜为L S47,宁波夜开花为L S48,观赏瓠子分离出为L S50,本地义序芋瓠瓜为L S53,南平青花插杆瓠为L S54,清流长芋瓠为L S55,思南瓠瓜为L S56,松桃瓠瓜分离为L S57,贵州长瓠瓜1为L S58,长瓠子为L S59,湖北瓠子为L S60,花皮葫芦分离出为L S61,永泰圆筒瓠为L S63,贵州长瓠瓜2为L S64,榕江瓠瓜为L S65,正阳瓠瓜为L S66,湖南早瓠瓜为L S67,雄溪瓠瓜为L S68,菜葫芦为L S69,三江口葫子为L S70,瓢葫芦为L S71,九山园蒲1为L S72,丽水农家种为L S73,义乌农家种为L S75,东阳农家种分1为L S76,东阳农家种分2为L S77,东阳农家种分3为L S78,汉龙碧玉为L S79,葫芦蒲为L S80,火焰蒲为L S81,金华农家种为L S82,九山园蒲2为L S83,南秀公主为L S84,爬地蒲为L S86,青皮圆蒲为L S87,三山牛腿蒲为L S88,绍兴长蒲为L S90,透年蒲为L S91,腰葫芦为L S93,引10-1②②为L S94,圆葫芦为L S95,浙江圆瓠1号为L S96,园蒲为L S97,越蒲1号为L S98,云南为L S99,早熟瓠子为L S101,砧木为L S102,08扁蒲-1为L S103,G 26②①/②为L S104,J 077-1 I 77①为L S105和长条瓠瓜为L S106。
供试材料经浸种、催芽、育苗、移植等一系列处理后,使用改进的C T A B法提取基因组DNA[6-8]。
1.2 方法
1.2.1 插入/缺失标记的来源
本试验所使用的插入/缺失标记是由瓠瓜豇豆组前期开展的RAD-Seq基因组测序开发而来,包括85条差异片段超过10 bp的多态性插入/缺失标记。
1.2.2 P C R反应条件
P C R体系包括10 ng的DNA,2.5 pm o l的引物,2.5 nmol的dN T P s,18.6 nmol的Mg C l2,0.2 U的r T aq酶以及10 xB uf f e r,用水补足体积至12.5 µL。P C R扩增程序为:①预变性94℃3 min;扩增循环:②变性94℃30 s,③退火52℃30 s,④延伸72℃40 s,②-④共重复36个循环;⑤72℃延伸5 min,⑥4℃保存。
1.2.3 聚丙烯酰胺凝胶电泳检测
P C R产物在室温条件下使用8%的垂直非变性聚丙烯酰胺凝胶(A c r∶B i s为19∶1或29∶1或39∶1)电泳分离,银染法染色,拍照分析。根据电泳结果记录清晰可重复的条带,将电泳图中同一位置上清晰且可重复的不同带型分别赋值为0,1, 2,以此类推,统计各引物的多态性条带的比率并作进一步分析。
1.2.4 遗传多样性分析
对每种基因型在每个位点上的等位基因数和出现频率进行统计。使用P I C_C a l c0.6(ht t p:// w w w.e s ni ps.com/do c/9171097b-a c 41-424a-9d35-e 7d 4e 540e c 9f/P i c a l c)计算每个插入/缺失位点的P I C值,计算公式为:其中:Pij代表在第i个位点出现第j个等位基因的频率[9]。使用N T SYSpc 2.10进行N e i's遗传距离计算(D A)和主坐标分析(P C o A)。最后根据D A的结果用SA H N程序中的U P G MA法进行聚类分析,并用E I G E N程序求特征值和特征向量进行主坐标分析,用T r e eP l o t模块生成聚类图,构建分子进化树,用3DP l o t生成主坐标图。
2 结果与分析
等位基因变异的多态性位点数和百分率不仅可以反映植物的多态性指数,而且可以评价引物反应多样性信息量的多少。本研究从85条插入/缺失引物中共筛选出22条多态性较好且能稳定扩增的引物。不同的引物所产生的条带数不同,最多的为8条(如引物插入/缺失-33590),最少的为1条(如引物插入/缺失-24033),扩增的DNA片段大小范围为75~500 bp,除了插入/缺失-17370(多态性条带大小为500 bp左右),多态性条带大小均为100~200 bp。22条引物共扩增出64条带,其中27条带表现出多态性,平均每条引物扩增2.78条带,平均多态性比例为42.19%(表1)。
基于插入/缺失扩增的22个多态性标记,通过N T SY S-pc(2.1版)软件对96份瓠瓜种质资源进行了聚类分析,得到了基于插入/缺失的聚类树状
图(图1)。瓠瓜种质之间的遗传距离(G D)变幅0~2.54。其中,L S73(丽水农家种)和L S82(金华农家种),L S12(广东早瓠瓜)和L S105(J 077-1 I 77①),L S2(孝感瓠子)和L S11(萧山圆蒲),L S9(东阳农家种1)和L S103(08扁蒲-1),L S25(武都瓠子)和L S61(花皮葫芦分离种)的G D均接近0.00,说明它们的亲缘关系极近。
从树状图中并无明显可作为分类标准的特性,但可以根据各种不同的趋势,将96份瓠瓜种质材料在遗传相似系数为0.68处分为8类(图1)。第1类与其他材料相似性系数最小,仅有0.36,即与其他材料的距离最远,仅有1份瓠瓜种质,即来自辽宁的小葫芦,该材料瓜型为单腰且较小,瓜长仅10cm左右。第2类也只有1份种质,来自宁夏的临夏瓠子,瓜型中长棒形,瓜色绿白,瓜条较长,早熟,该材料与其他材料相比地理来源较远,亲缘关系也较远。第3类同样只包括1份瓠瓜材料,为来自新疆的瓢葫芦,瓜色浅绿,瓜型为长把梨形且较小,瓜长19cm左右,早熟。第4类包括3份材料,分别为来自浙江的全华蒲瓜、肖长蒲和杭州长瓜,瓜型均为长棒形,瓜色绿色,瓜长45~50cm,早熟。第5类共有9份材料,均表现为早熟,其中7份材料瓜色为绿白相间,而这7份材料中有5份材料的瓜型均为牛腿形,2份来自云南的材料也聚集在一起。第6类包括5份材料,分别为清秀、新蔡瓠瓜、麦瓠、南平青花插杆瓠、砧木。这组材料瓜色均表现为绿色,其中除砧木以外的4份材料瓜型为棒形,瓜长较短,早熟。第7类仅有1份材料,绍兴长蒲,瓜型棒形,瓜色油绿,瓜条较长(41cm),熟性较早。余下的75份材料被归为第8类,以瓜型不同又可细分为3组,第1组瓜型为棒形(除腰葫芦外),包括8份材料,瓜皮绿色,瓜长中等(25~35cm),早熟。第2组材料包括17份,瓜型多为牛腿形,近圆形有5份,占到近圆形材料总数的1/2;瓜色均表现为绿色,瓜长较短,熟期较晚,仅有的3份晚熟型的材料(火焰蒲、透年蒲和圆葫芦)均聚集在本组。第3组包括50份材料,来源较广泛,瓜型大部分为棒形,瓜色多变,以纯绿色为主,本试验中全部9份极早熟材料(萧山园蒲、长蒲1、长蒲2、长蒲3、长蒲4、津胜特选、湖南圆瓠1号、观赏瓠子分离种)均聚集在本组。
在主成分分析中,没有明显的可以作为分类标准的特征,材料较平均地分布于图中,但相对于产地而言,相同瓜型的材料有聚集在一起的趋势(图2)。上窄下宽的牛腿形、长把梨形材料,大部分集中分布在图左部和右上的位置;近圆形材料在图上分布较分散,但大部分集中分布在中间靠左的位置;棒筒形材料最多,集中在图中间部分,这与树状图的结果基本一致。

表1 插入/缺失引物多态性分析
3 小结和讨论
我国瓠瓜资源众多,遗传变异非常丰富,但较为全面、系统的遗传多样性分析还比较少见。应用分子标记技术研究分析主栽品种的亲本与优良品系间的亲缘关系,可为瓠瓜育种实践中优化亲本选配,减少杂交组合数目,提高品种质量及育种效率提供依据。通常,在阐明种质资源的遗传多样性后,选择能最大程度代表物种遗传多样性的核心种质构成核心种质库是长期整合、保存种质基因资源、方便育种亲本选配的重要途径。
插入/缺失标记是指两亲本间在全基因组序列中的差异,在生物的遗传多样性分析、纯度鉴定、亲子分析等方面已有广泛的应用。然而,插入/缺失引物在蔬菜中应用较少,在瓜类中更加少见。本研究首次使用插入/缺失引物研究非模式物种---
瓠瓜的遗传多样性。Decker-Walters等[10]利用38个R A P D标记研究了世界范围内74个瓠瓜品系的遗传多样性,发现世界范围内瓠瓜可以大洲为界限划为不同类群,非洲、亚洲的瓠瓜种质相对独立,新大陆(美洲)的种质起源于非洲,但受到亚洲种质的一些影响,而商业品种由于地理起源和遗传组成的差异有很大不同,由于这些品种的流通使得难以完整地界定某些品种。Y e t i s i r等[11]利用形态
学标记研究土耳其的182份瓠瓜的果实和种子材料,在统计学和多变量分析共同研究所得的数据基础上,发现瓜型和大小可以作为区分瓠瓜亚类的主要标准,这与本研究的结论一致。
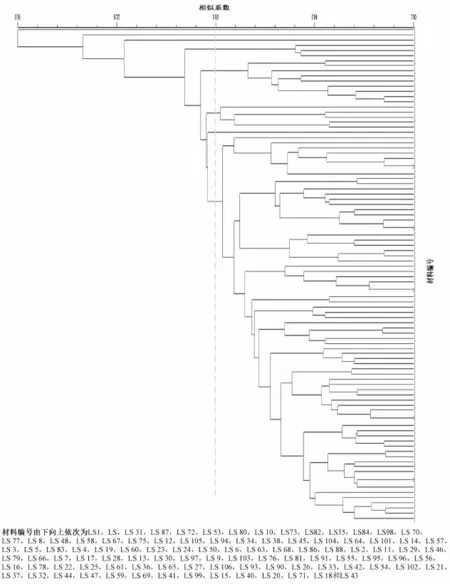
图1 96份瓠瓜材料聚类分析树状图
本研究利用插入/缺失标记方法,从分子水平上对我国96份瓠瓜资源的遗传多样性及亲缘关系进行研究,可以为我国瓠瓜资源的合理开发利用以及进一步开展瓠瓜育种研究、指导亲本选配提供重要依据;同时可为瓠瓜种质的分类、核心种质库的构建,以及瓠瓜种质资源的进一步收集保存、创新利用提供分子水平上的依据,以缩短瓠瓜育种年限,提高育种效率。
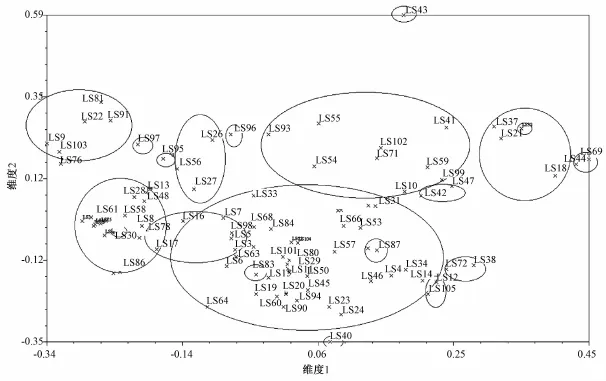
图2 96份瓠瓜材料插入/缺失聚类分析二维坐标图
[1]BeevyS S, Kuriachan P.Chromoso menumbers of south IndianCucurbita ceae and anoteon thecytologicalevolution in thefamily[J].Journal of Geneticsand Cytology, 1996, 31: 65 -71.
[2] Decker-WaltersD S, Wilkins-EllertM, ChungS M, etal.Discoveryand geneti cassess ment of wild bottlegourd [ Lagenaria siceraria (Mol.) Standley; Cucurbitaceae] fromZimbabwe[J].EconomicBotany, 2004, 58 (4): 501 -508.
[3] Lee JM.Cultivation of grafted vegetablesI.Currentstatus,grafting methods, and benefits[J].HortScience, 1994, 29(4) : 235 -239.
[4] 彭庆务,何晓明,谢大森,等.瓠瓜的特征特性及育种对策[J].广东农业科学,2003(1):18-19.
[5] Yetisir H,Sari N.Effect of different root stock onplant growth,yield and quality of watermelon[J].Ani alProduction Science,2003,43(10):1269-1274.
[6] 潭俊杰,蒋光明.蔬菜栽培学各论[M].北京:中国农业出版社,1999.
[7] 吕家龙.蔬菜栽培学各论:南方本[M].北京:中国农业出版社,2001.
[8]MaguireTL, CollinsGG, SedgleyM.Amodified CTABDNAextraction procedureforplantsbelongingtothefamilyProteaceae[J].PlantMolecularBiologyReporter, 1994, 12 (2) : 106 -109.
[9]Anderson JA, ChurchillGA, AutriqueJE, etal.0ptimizingparentalselection forgeneticlinkagemaps[ J].Genome,1993, 36 (1) : 181 -186.
[10]Decker-WaltersD, Staub J, Lopez-SeseA, etal.Diversityin land racesand cultivars of bottlegourd ( Lagenaria siceraria;Cucurbitaceae) asassessed byrandom amplified polymorphicDNA [J].GeneticResourcesand Crop Evolution, 2001, 48(4) : 369 -380.
[11]YetişirH, ŞakarM, SerceS.Collection and morphologicalcharacterization ofLagenaria siceraria germplasm from theMediterranean region ofTurkey[J].GeneticResourcesandCrop Evolution, 2008, 55 (8) : 1257 -1266.
(责任编辑:高 峻)
Q75文献标志码:A文章编号:0528-9017(2014)05-0668-05
2013-11-05
浙江省农业新品种选育专项(2012C 12903)
鲁忠富(1963-),男,浙江杭州人,实验师,从事蔬菜育种及新品种推广工作。E-mail:l z f 0522@163.com。
,李国景。E-mail:l ig j@m a i l.z a a s.a c.c n。
文献著录格式:鲁忠富,王莎,吴晓花,等.利用插入/缺失标记分析我国瓠瓜资源遗传多样性[J].浙江农业科学,2014(5):668-672.
猜你喜欢
今日农业(2022年1期)2022-11-16
今日农业(2022年14期)2022-09-15
今日农业(2022年13期)2022-09-15
娃娃乐园·综合智能(2022年8期)2022-07-28
西江月(2021年3期)2021-11-13
心声歌刊(2021年6期)2021-02-16
心声歌刊(2020年5期)2020-11-27
儿童时代(2020年5期)2020-06-03
四川蚕业(2020年4期)2020-02-10
中国麻业科学(2018年6期)2018-04-09
