
富营养化对浅水湖泊轮虫种群结构影响研究
2014-02-25 02:49:08郑金秀池仕运李聃汪红军胡菊香
生态环境学报 2014年12期
郑金秀,池仕运,李聃,汪红军,胡菊香
富营养化对浅水湖泊轮虫种群结构影响研究
郑金秀,池仕运,李聃,汪红军,胡菊香*
水利部中国科学院水工程生态研究所 水利部水工程生态效应与生态修复重点实验室,湖北 武汉430079
石臼湖和固城湖是长江中下游两个浅水湖泊,富营养化问题日益突出。为探索湖泊富营养化对轮虫的影响,于2012年平水期和枯水期对这两个湖泊轮虫种群结构进行了调查,并检测水体理化因子,分析轮虫分布与水环境因子及水体富营养状况的关系。水质检测结果显示,平水期石臼湖除个别点水体综合营养状态指数(TLI)>50为轻度富营养状态外,大多数站点TLI<50,为中营养状态;枯水期两个湖泊TLI>50,处于轻度-中度富营养化状态。水生生物调查检出轮虫35种,其中石臼湖33种,固城湖27种,平水期种类多于枯水期。出现的轮虫以臂尾轮虫(Brachionus)和异尾轮虫(Trichocerca)居多,优势种主要有螺形龟甲轮虫(Keratella cochlearis)、广生多肢轮虫(Polyarthra vulgaris)、疣毛轮虫(Trichocerca sp.)、沟痕泡轮虫(Pompholyx sulcata)、异尾轮虫属和臂尾轮虫属等。轮虫平均密度3023.19 ind·L-1,平均生物量745.72 µg·L-1。现存量季节变动明显,大部分站点平水期大于枯水期。轮虫多样性指数也是平水期大于枯水期,石臼湖轮虫多样性大于固城湖。水体综合营养状态指数(TLI)与轮虫种类数和多样性成显著负相关,随着富营养化加剧,轮虫种类减少,多样性降低。典范对应分析(Canonical Correspondence Analysis,CCA)表明,影响轮虫种群分布的主要环境因子为NH4+-N、TN、TP和CODMn。按轮虫丰度分布与水环境因子的关系排序,2次调查的24个站点可大致分成3个组。轮虫种群结构受水体富营养状态影响,每组站点的代表性种类对水体营养状态有较好的指示作用。组1的代表性种类为曲腿龟甲轮虫,指示营养级别O-β;组2代表性种类萼花臂尾轮虫,指示的营养级别为β-α;组3以O营养级种类为主。
轮虫;种群结构;富营养化;浅水湖泊
1 材料与方法
1.1样品采集
2012年10月(平水期)、12月(枯水期)对石臼湖和固城湖进行轮虫采样调查,同时对水体理化因子进行分析。根据湖泊面积及生境特点,在石臼湖湖区布设8个采样点,在固城湖湖区布设4个采样点。具体采样点设置如图1。
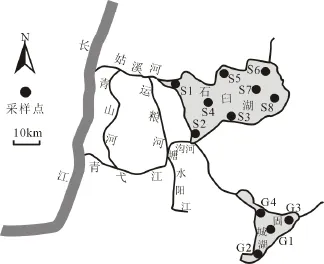
图1 采样点示意图Fig. 1 Geographical distribution of sampling sites
1.2样品处理
轮虫定性样品用25号浮游生物网(孔径为64 μm)大范围拖捞获得,取50 mL用5%的福尔马林固定。定量样品用2.5 L采水器于3个深度(表层、中层、底层)各采水5 L,混合后取2 L水用15%的鲁哥氏液固定。所获样品均带回实验室进行分类、鉴定(王家楫,1961;诸葛燕,1997)。
水环境监测指标包括总氮、总磷、铵氮、硝氮、亚硝氮、可溶性磷酸盐、叶绿素a和化学耗氧量等,各指标的测定方法参照《水和废水监测分析方法》进行(国家环境保护总局《水和废水监测分析方法》编委会,2002)。
1.3数据处理
根据中国环境监测总站制定的《湖泊(水库)富营养化评价方法及分级技术规定》,选取叶绿素a(Chl a)、总磷(TP)、总氮(TN)、透明度(SD)和高锰酸盐指数(CODMn)等为参数,计算综合营养状态指数(金相灿等,1995)。计算公式为:
式中:TLI(∑)—综合营养状态指数;
Wj—第j种参数的营养状态指数的相关权重;
TLI(j)—代表第j种参数的营养状态指数。
以Chl a作为基准参数,则第j种参数的归一化的相关权重计算公式为:
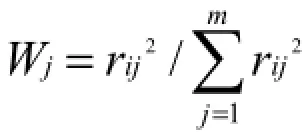
(rij为第j种参数与基准参数chl a的相关系数,Chl a、TP、TN、SD和CODMn的rij值分别为1、0.84、0.82、-0.83和0.83,m为评价参数的个数)
各参数的营养状态指数计算公式为:
TLI(chl a)=10(2.5+1.086lnChl a)
TLI(TP)=10(9.436+1.624lnTP)
TLI(TN)=10(5.453+1.694lnTN)
TLI(SD)=10(5.118-1.94lnSD)
TLI(CODMn)=10(0.109+2.661ln CODMn)
式中Chl a单位为mg·m-3,SD单位为m,其它指标单位均为mg·L-1。
轮虫的生物多样性采用Margalef多样性指数(D)和Shannon-Wiener多样性指数(H),公式分别为:
D=(S-1)/lnN,H=-Σ(Pi)(lnPi)
式中,S为种类数,Pi为某种轮虫的密度占总个体密度(ind·L-1)的比例,N为轮虫的密度(ind·L-1)。
用软件PC-ORD 5进行典范对应分析(Canonical Correspondence Analysis,CCA),用于分析轮虫群落与水体环境参数之间的关系,得出物种与环境二维排序图,再用非参数程序多响应置换过程(multi response permutation procedures,MRPP)检验各组之间的差异性,并做指示种分析(indicator species analysis,ISA),找出各组的特征指示种。
2 结果与分析
2.1水环境因子变化
石臼湖和固城湖平水期平均水温20 ℃,枯水期平均水温9 ℃。根据地表水环境质量标准(GB3838-2002)单因子评价方法,两个湖泊各采样点总体符合地表水环境质量III类标准,平水期水质优于枯水期。枯水期石臼湖S2、S4和S8等采样点及固城湖G2、G3、G4采样点高锰酸盐指数含量较高,枯水期固城湖氨氮含量较高,均超过III类
水质标准。各采样点水环境因子平均值如表1。采用综合营养状态指数法,对石臼湖和固城湖水体营养状态进行评价,各采样点的综合营养状态指数变化如图2。平水期时只有石臼湖的S1、S2和S3采样点TLI>50,为轻度富营养状态,其余站点都<50,处于中营养状态。枯水期石臼湖和固城湖所有站点TLI都>50,呈富营养状态,其中S7和S8站点分别达到60.4和60.7,为中度富营养状态。
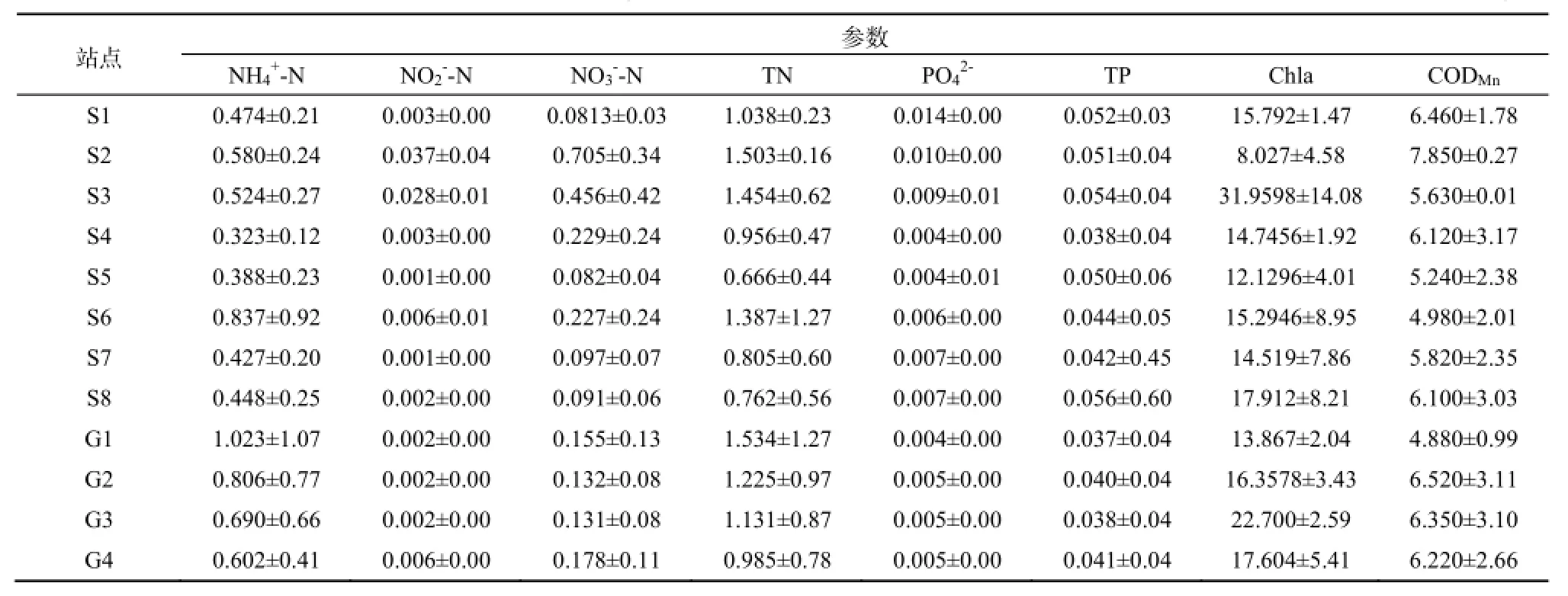
表1 两个湖泊水环境因子平均值Table 1 Average value of environmental factors in two lakes mg·L-1
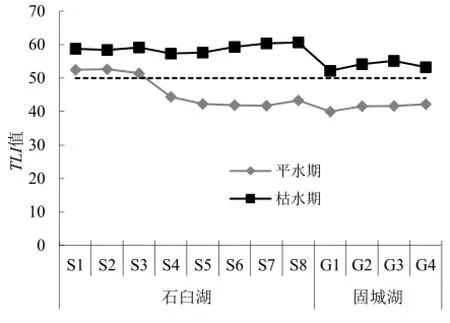
图2 石臼湖和固城湖综合营养状态指数变化Fig. 2 Comprehensive nutrition state index variation in Shijiu Lake and Gucheng Lake
2.2种类组成
石臼湖和固城湖调查中共采集到轮虫35种,隶属于10科21属,其中石臼湖采集到33种,固城湖采集到27种。轮虫的种类组成中,异尾轮属(7种)和臂尾轮属(6种)种类最多,分别占总种数的20.0%和17.1%;其次是龟甲轮属(4种),占总种数11.4%,其它各属种类均很少。石臼湖常见种主要有螺形龟甲轮虫(Keratella cochlearis)、广生多肢轮虫(Polyarthra vulgaris)、疣毛轮虫(Trichocerca sp.)等;固城湖常见种主要为螺形龟甲轮虫、广生多肢轮虫、沟痕泡轮虫(Pompholyx sulcata)、异尾轮虫(Trichocerca sp.)、角突臂尾轮虫(Brachionus angularis)等(见表2)。
两个湖泊轮虫皆是平水期种类多于枯水期种类,石臼湖平水期27种,枯水期22种;固城湖平水期20种,枯水期17种(图3)。石臼湖各采样点检出的轮虫种类数差异较大,平水期岸边种类多于湖心,枯水期则以湖心种类居多。石臼湖轮虫季节变化幅度也比固城湖大。这与各采样点的生境有很大关系。石臼湖的8个采样点,除了S4和S7位于湖心,其余6个点都靠近岸边。受围湖造田、养殖等人类活动影响,石臼湖岸边带有机质含量高,轮虫饵料丰富,种类增多。而固城湖的4个采样点皆位于敞水水面,离岸边带有一定距离,且固城湖为江苏省饮用水源区,人类活动较石臼湖少,也导致了固城湖轮虫种类数量及其季节变化的稳定。此外,石臼湖是通江浅水湖泊,受长江影响,湖泊水位变幅大,生境较固城湖复杂,也导致了其生物类群时间和空间的较大差异。
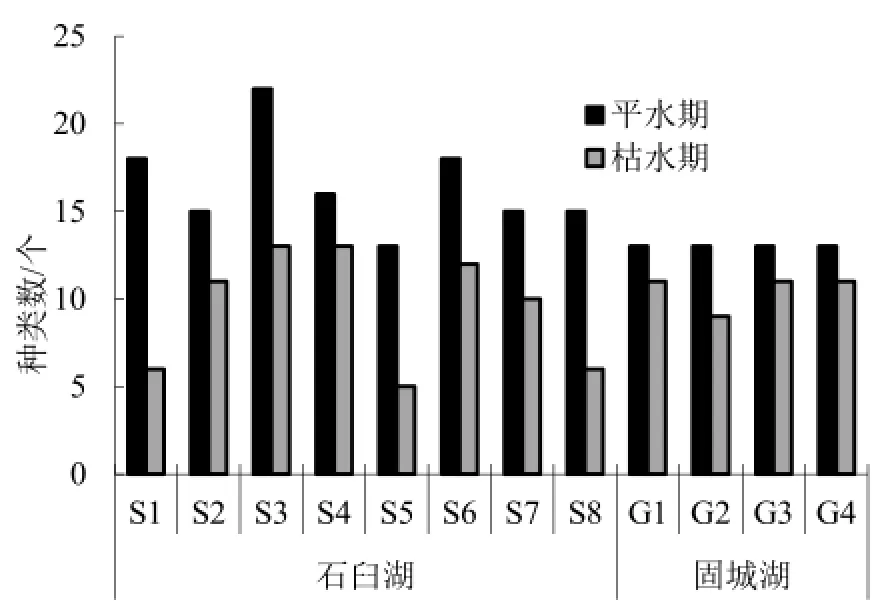
图3 两个湖泊的轮虫种类分布Fig. 3 Rotifer species distribution in two lakes
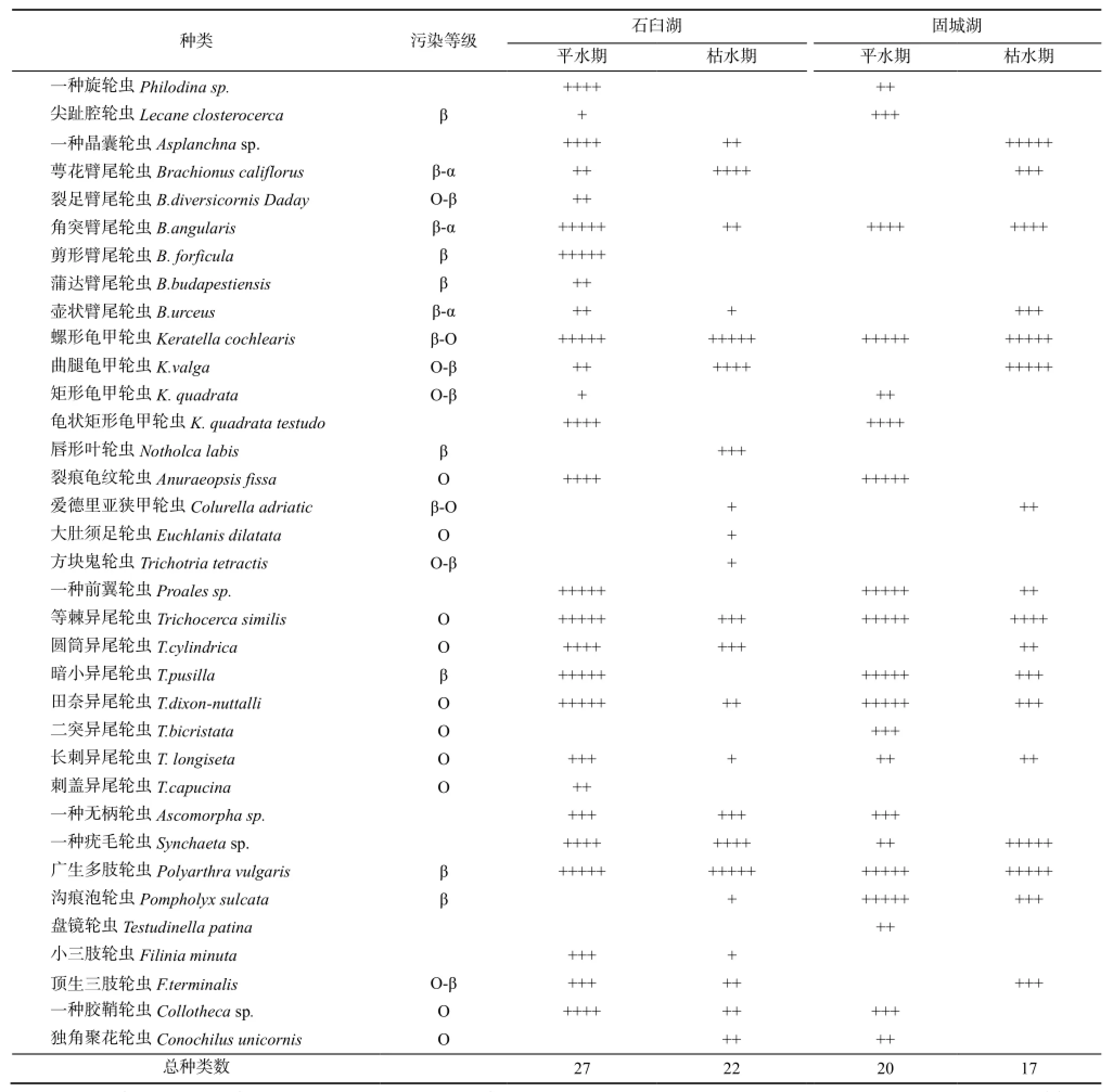
表2 两个湖泊轮虫种类组成Table 2 Rotifer species composition in two lakes
2.3轮虫现存量变化
2次调查轮虫密度变动在50~13566.67 ind·L-1,平均3023.19 ind·L-1,生物量变动在8.50~4912.75 µg·L-1,平均745.72 µg·L-1。密度最大值出现在平水期固城湖G2站点,为13566.67 ind·L-1,生物量最大值出现在枯水期石臼湖S1站点,为4912.75 µg·L-1。石臼湖平均密度为2576.04 ind·L-1,平均生物量为955.93 µg·L-1;固城湖平均密度为3917.50 ind·L-1,平均生物量为325.29 µg·L-1。两个湖泊轮虫现存量季节变动较明显,大部分站点平水期现存量大于枯水期,只有站点S1和S2枯水期大于平水期(图4)。另外站点S4和G4生物量为枯水期大于平水期。这与不同季节出现的轮虫种类有关,这些站点在枯水期出现了较多大个体的轮虫,如疣毛轮虫和萼花臂尾轮虫。固城湖轮虫密度大于石臼湖,而生物量却小于石臼湖,也与两个湖泊出现的优势种有关,固城湖优势种主要为螺形龟甲轮虫、暗小异尾轮虫、沟痕泡轮虫等,个体较小,而石臼湖优势种则包括了疣毛轮虫和臂尾轮虫等较大个体的轮虫。
2.4轮虫多样性指数与营养状态关系
轮虫在不同季节表现出不同的多样性。平水期
石臼湖Shannon-wiener指数(H)在1.57~3.14之间变动,平均2.48;Margalef指数(D)在0.52~2.14之间变动,平均1.40。枯水期石臼湖H值在1.00~2.35之间变动,平均1.85;D值在0.26~1.00之间变动,平均0.67。平水期固城湖H值在1.36~2.52之间变动,平均1.97;D值在1.14~1.42之间,平均1.26。枯水期固城湖H值在0.94~2.17之间,平均1.64;D值在0.33~1.05之间,平均0.64。轮虫多样性指数平水期高于枯水期,石臼湖的轮虫多样性高于固城湖(图5)。

图4 轮虫现存量变化Fig. 4 variation of rotifer density and biomass
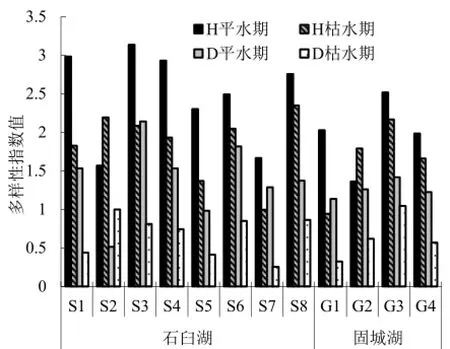
图5 轮虫多样性指数Fig. 5 variation of rotifers diversity index
我们通过综合营养状态指数(TLI)与轮虫多样性指数及轮虫种类数的pearson相关性分析得到,水体富营养状态与轮虫种类数和多样性指数成负相关,其中TLI与种类数和Margalef指数(D)相关性显著(表3)。说明营养增加,轮虫种类减少,多样性降低。
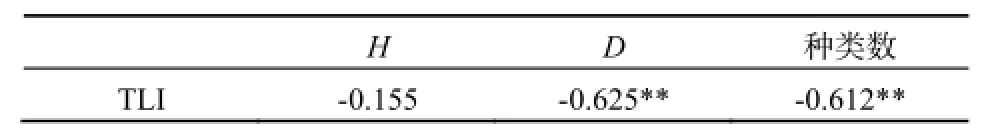
表3 TLI与轮虫多样性指数和种类数的相关性Table 3 Correlation analysis of TLI and biodiversity index and species
2.5轮虫分布与水质的关系
为检验轮虫种群结构与水质参数的关系,将各站点轮虫丰度与水质参数做CCA分析,可得到图6结果。统计分析结果显示,CCA轴的轴1解释率为0.491,轴2解释率为0.274,轴3解释率为0.180。轴1代表了大部分环境因子参量信息,与环境因子相关性显著(r=0.975,P=0.002),各环境因子与3个轴之间的关系见表4。可以看出,轴1与温度T成显著正相关,与TP、CODMn、TN和NH4+-N成负相关,轴2主要与NH4+-N成正相关。影响轮虫分布的主要水环境因子为温度T、CODMn、TP、TN和NH4+-N。
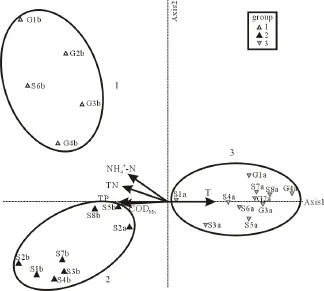
图6 轮虫站点分布与水环境变量的CCA排序图Fig. 6 CCA ordination biplots of site scores and selected environmental variables
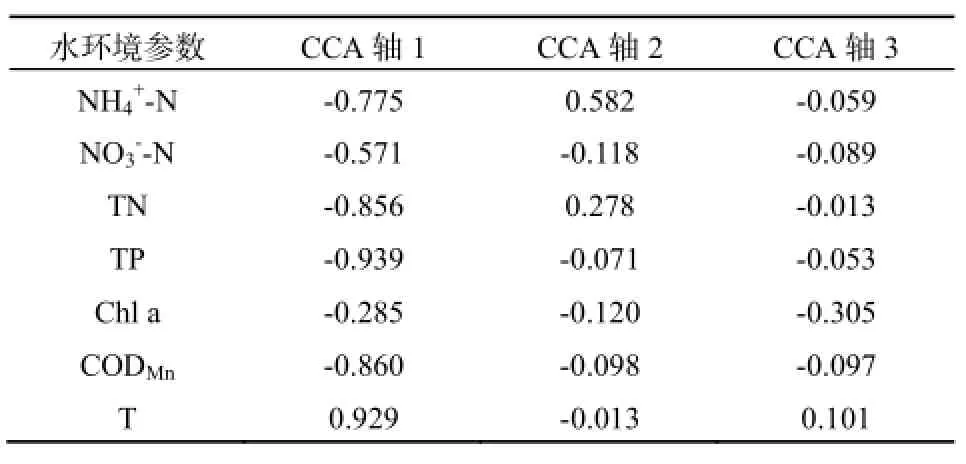
表4 水环境参数与CCA 3轴之间的关系Table 4 Relationship between the environmental parameters and three axis
按轮虫丰度分布与水环境因子的关系排序,2次调查的24个站点可大致分成3个组(如图6)。经非参数程序多响应置换过程(multi response permutation procedures,MRPP)检验,3个组之间差异显著(统计量T=-9.46,A=0.21,P<0.01),说明依照环境因子排序划分的3个组是合理的。
3 讨论
3.1环境因子对轮虫分布的影响
轮虫的种群动态与诸多环境因子密切相关(Herzig,1987;Duggan等,2002),其中,温度对轮虫的生长繁殖和种群分布起了关键作用(王家楫,1961;Armengol等,1998;Mikschi,1989;杨宇峰等,2006)。诸多学者研究了轮虫群落结构与温度的关系(都雪等,2014;温利新等,2006),认为温度是影响轮虫密度季节变化的影响因素之一。本研究中,分布于CCA轴2两侧的站点分别代表了平水期站点和枯水期站点,也说明温度是决定轮虫季节分布的主要因素。从优势种组成变化来看,除了螺形龟甲轮虫和广生多肢轮虫为全年优势种,裂痕龟纹轮虫、前翼轮虫、暗小异尾轮虫及角突臂尾轮虫、剪形臂尾轮虫等主要出现在平水期,而曲腿龟甲轮虫、萼花臂尾轮虫则主要出现在枯水期。这主要与食物资源的季节变化有关(Pourriot,1997),相比于枯水期,平水期温度较高,藻类繁殖较快,而浮游动物中广温性种类较多,其中以藻类为食的臂尾轮虫和暗小异尾轮虫,及以碎屑为主要食物的裂痕龟纹轮虫就得到相应的增长(温利新等,2006;刘红等,2006)。
营养状态参数同样影响着轮虫的种群结构(Yoshida等,2003)。本研究中,总氮、总磷、高锰酸盐和氨氮与轮虫种类数呈显著负相关(P<0.05),这些营养盐综合影响了轮虫的种群分布。TP含量是判定水体富营养化程度的一个重要指标,水体的富营养化程度随着TP含量的升高而加剧,从而影响轮虫的结构组成(都雪等,2014)。Jeppesen等(2000)认为,轮虫的种类数随着TP的增加而下降,这与本研究结果一致。Tasevska等人(2012)研究表明,轮虫密度随营养盐的增加而增加。Wang等(2010)研究显示,轮虫密度与水体无机氮含量呈显著正相关。本研究中,轮虫密度与TN呈正相关,但相关性并不显著。这可能与石臼湖和固城湖所处的营养状态有关,也与通江湖泊较复杂的生境变化有关。在湖泊富营养化过程中,轮虫的种类和密度都将发生演变,一般认为,水体富营养化,将使一些生物敏感种消失,使生物群落结构单一化,而耐受种的大量繁殖,使轮虫密度增加(钱方平等,2007)。从TLI值来看,石臼湖和固城湖分别处于轻度富营养化和中营养-轻度富营养化阶段,其影响首先体现在种类的变化上,其次才影响到轮虫的密度。除了水体理化因子,轮虫的现存量还受食物资源、生物之间的竞争捕食等诸多因素的影响(Devetter,1998;William,1984)。由于养殖和围湖造田等人类活动干扰,石臼湖和固城湖氨氮含量和高锰酸盐指数含量较高,一些站点超过III类水质标准,这些污染物都会对轮虫结构产生影响,污染物过多也将抑制轮虫的生长繁殖(朱玮阁等,2009;Garcia-Garcia等,2007)。
3.2富营养化对轮虫多样性的影响
水体富营养化对水生生物群落结构及其物种多样性的影响已有较多报道(胡菊香等,2007;郭凯等,2010;Dodson,1992)。综合营养状态指数(TLI)与轮虫多样性指数及种类数的相关性分析(表3)表明,水体富营养状态与轮虫种类数和多样性指数成负相关,随着水体富营养化程度的增加,轮虫种类减少,多样性降低。这与熊金林等(钱方平等,2007;熊金林等,2003)研究结果一致。湖泊发生富营养化的过程中,物种多样性一般先升高,后下降。其间,在中营养水平时生物群落结构复杂,物种多样性较高,而后逐渐下降(日本生态学会环境问题专门委员会,1987)。石臼湖和固城湖处于不同的富营养化阶段,但都超过了中营养水平,因此,随着TLI的升高轮虫多样性呈下降趋势。轮虫多样性指数与水体污染程度有一定关系,许多学者研究(陈立婧等,2008;饶小珍等,2000)认为轮虫可作为水质监测的指标,开展水质状况评价。按轮虫多样性指数的评价结果,石臼湖和固城湖为富营养化状态,与综合营养状态指数评价结果一致,但富营养化程度按轮虫多样性指数评价为石臼湖<固城湖,这与综合营养状态指数评价结果相反。可见,在确定富营养化污染等级上,单纯以轮虫多样性指数为标准是不够的,还应探讨轮虫的种类组成,或与其他参数综合分析。这与赵红雪等人的研究结论一致(赵红雪等,2011;李共国和虞左明,2003)。
3.3富营养化对轮虫种群结构的影响
石臼湖和固城湖的轮虫分布与水体富营养状态密切相关。通过CCA分析将两个湖泊的站点大致分为三组,其依据是水环境参数的排序。然而,分析这三组出现的轮虫种类,我们同样可以发现,其区别于另外两组的代表性种类对水体营养状态也有很好的指示作用。用PC-ORD做指示种分析(indicator species analysis,ISA),可以得出表5结果。
参照Sladecek(1983)划分的轮虫指示种类别,组1的代表性种类曲腿龟甲轮虫,所指示的营养级别为O-β;组2代表性种类萼花臂尾轮虫,指示的营养级别为β-α;组3种类较复杂,但以O营养级种类为主。以此来划分,则表示组2是富营养状态较高的水域,组1其次,组3相对营养状态低些。这与调查的水质状况和轮虫结构是相符的。组1站
点组成以固城湖枯水期站点为主,组2除了S2之外,全是石臼湖枯水期站点,而组3则是石臼湖和固城湖平水期站点。调查结果已显示,两个湖泊平水期水质优于枯水期,平水期营养状态也较枯水期低。

表5 指示种分析结果Table 5 The results of ISA
计算石臼湖好固城湖各类型指示种所占比例,两个湖泊不同季节出现的种类中,平水期O和O-β类型轮虫所占比例(石臼湖80%,固城湖53%)都高于枯水期(石臼湖56%,固城湖36%),这再次说明了轮虫种类演变与富营养化有一定关系。从平水期石的中营养-轻富营养状态至枯水期的轻度富营养-中度富营养状态,轮虫种类不仅随营养水平上升而下降,而且出现的种类也与营养水平相适应。
这种演变除了与季节和食物资源有关,也体现了优势种对污染的耐受性及生物之间的竞争关系。如得到较大认同的10种富营养化轮虫指示种(胡菊香等,2007),在石臼湖和固城湖的不同季节都有个别种类成为优势种。而同一属的不同种往往出现在不同季节。如臂尾轮虫,在平水期主要出现角突臂尾轮虫和剪形臂尾轮虫,而枯水期则出现萼花臂尾轮虫,除了萼花臂尾轮虫对污染的耐受力和对低温适应性较强外(耿红,2006;王爱民等,2014),这种现象也应该和种间竞争机制有关(谢钦铭等,1997),同样的,曲腿龟甲轮虫和矩形龟甲轮虫也分别出现在不同的时期,而螺形龟甲轮虫则成为全年优势种,这说明了螺形龟甲轮虫对富营养化有较强的适能力。
ARMENGOL X, ESPARCIA A, MIRACLE M R. 1998. Rotifer vertical distribution in a strongly stratified lake:a multivariate analysis[J]. Hydrobiologia, 387/388: 161-170.
BERZINS B and PEJLER B. 1989. Rotifer occurrence and trophic degree[J]. Hydrobiologia, 182: 171-180.
CHEN L J, LIU Q, PENG Z, et al. 2012. Rotifer community structure and assessment of water quality in Yangcheng Lake[J]. Chinese Journal of Oceanology and Limnology, 30(1): 47-58.
DODSON S I. 1992. Predicting crustacean zooplankton species richness[J]. Limnol Oceanogr, 37: 848-856.
DUGGAN I C, GREEN J D and SHIEl R J, 2002. Distribution of rotifer assemblages in North Island, New Zealand, Lakes:relationships to environmental and historical factors[J]. Freshwater Biology, 47: 195-206.
MIKSCHI E. 1989. Rotifer distribution in relation to temperature and oxygen content[J]. Hydrobiologia, 186/187: 209-214.
GARCIA-GARCIA G, PICAZO-PAEZ E A, NANDINI S, et al. 2007. Combined effects of sediment and lead(PbCl2) on the demography of Brachionus patulus (Rotifera: Brachionidae)[J]. Hydrobiologia, 593: 209-218.
HERZIG A. 1987. The analysis of planktonic rotifer populations: A plea for long-term investigations[J]. Hydrobiologia, 147: 163-180.
JEPPESEN E, JENSEN J P, KRISTENSEN P, et al. 2000. Trophic structure,species richness and biodiversity in Danish lakes: changes along a phosphorus gradient[J]. Freshwater Biology, 45: 201-218.
LESLIE E HOLLAND, C F BRYAN, J PRESTON NEWMAN JR. 1983. Water quality and the rotifer populations in the Atchafalaya River Basin, Louisiana[J]. Hydrobiologia, 98(1): 55-59.
DEVETTER M. 1998. Influence of environmental factors on the rotifer assemblage in an artificial lake[J]. Hydrobiologia, 387/388: 171-178.
TASEVSKA O, JERSABEK C D, KOSTOSKI G, et al. 2012. Differences in rotifer communities in two freshwater bodies of different trophic degree (Lake Ohrid and Lake Dojran, Macedonia)[J]. Biologia, 67/3: 55-572.
POURRIOT R. 1977. Food and feeding habits of Rotifera[J]. Arch Hydrobiol Beih Ergebn Limnol, 8: 243-260.
SLADECK V. 1983. Rotifers as indicators of water quality[J]. Hydrobiologia, 100: 169-201.
YOSHIDA T, URABE J, ELSER J J. 2003. Assessment of ‘top-downʼ and‘bottom-upʼ forces as determinants of rotifer distribution among lakes in Ontario, Canada[J]. Ecological Research, 18: 639-650.
OBERTEGGER U and MANCA M. 2011. Response of rotifer functional groups to changing trophic state and crustacean community[J]. Limnol, 70(2): 231-238.
WEN X L, XI Y L, QIAN F P, et al. 2011. Comparative analysis of rotifer community structure in five subtropical shallow lakes in East China:role of physical and chemical conditions[J]. Hydrobiologia, 661: 303-316.
WANG S B, XIE P, GENG H. 2010. The relative importance of physicochemical factors and crustacean zooplankton as determinants of rotifer density and species distribution in lakes adjacent to the Yangtze River, China[J]. Limnologica, 40: 1-7.
WILLIAM E N. 1984. Regulation of rotifer densities by crustacean zooplankton in an oligotrophic montane lake in British Columbia[J]. Oecologia(Berlin), 61: 175-181.
陈立婧, 顾静, 彭自然, 等. 2008. 滆湖轮虫群落结构与水质生态学评价[J]. 动物学杂志, 43(3): 7-16.
都雪, 王齐东, 张超文, 等. 2014. 洪泽湖轮虫群落结构及其与环境因子的关系[J]. 湖泊科学, 26(2): 269-276.
耿红. 2006. 水体富营养化和蓝藻对轮虫影响的生态毒理学研究[D]. 武汉: 中国科学院水生生物研究所: p99-108.
郭凯, 赵文, 殷守仁, 等. 2010. 北京官厅水库轮虫群落结构与水体富营养化状况[J]. 湖泊科学, 22(2): 256-264.
国家环境保护总局《水和废水监测分析方法》编委会. 2002. 水和废水监测分析方法(第4版)[M]. 北京: 中国环境科学出版社.
胡菊香, 吴生桂, 陈金生, 等. 2007. 巢湖富营养化对轮虫的影响研究[J].环境科学与技术, 30(12): 16-18.
金相灿, 刘树坤, 章宗涉. 1995. 中国湖泊环境[M]. 北京: 海洋出版社.
李共国, 虞左明. 2003. 千岛湖轮虫群落结构及水质生态学评价. 湖泊科学, 15(2): 169-178.
刘红, 马徐发, 刘瑾. 2006. 武汉南湖浮游轮虫的初步研究[J]. 水利渔业, 26(2): 60-62.
毛春梅, 张文锦. 2007. 固城湖富营养化评价及防治对策[J]. 安徽农业科学, 3(28): 9005-9006, 9030.
钱方平, 席贻龙, 温新利, 等. 2007. 湖泊富营养化对轮虫群落结构及物种多样性的影响[J]. 生物多样性, 15(4): 344-355.
饶小珍, 许友勤, 陈寅山. 2000. 福州内河的轮虫与水质污染评价[J]. 福建师范大学学报(自然科学版), 16(1): 71-75.
日本生态学会环境问题专门委员会编, 卢全章译. 1987. 环境指示生物·水域分册[M]. 北京: 中国环境科学出版社.
史小丽, 秦伯强. 2007. 长江中下游地区湖泊的演化及生态特性[J]. 宁波大学学报(理工版), 20(2): 221-226.
王爱民, 席贻龙, 牛翔翔, 等. 2014. 汀棠湖冬季出现的萼花臂尾轮虫对水温的适应[J]. 生态学报, 34(16): 4588-4595.
王家楫. 1961. 中国淡水轮虫志[M]. 北京: 科学出版社.
王庆, 刘丽君, 杨宇峰. 2010. 南方某水库轮虫群落特征及其对环境因子的响应[J]. 生态学报, 30(13): 3385-3395.
王荣娟, 张金池. 2011. 石臼湖湿地水环境质量评价及富营养化状况研究[J]. 湿地科学与管理, 7(2): 26-28.
温新利, 席贻龙, 张雷, 等. 2006. 芜湖市镜湖轮虫群落结构分析及水质的生态学评价[J]. 水生生物学报, 30(2): 152-158.
谢钦铭, 李云, 李长春. 1997. 鄱阳湖轮虫种类组成与现存量季节变动的初步研究[J]. 江西科学, 15(4): 235-242.
熊金林, 梅兴国, 胡传林. 2003. 湖北四湖泊营养类型与轮虫群落的关系[J]. 动物学杂志, 38(6): 8-14.
杨宇峰, 王庆, 陈菊芳, 等. 2006. 河口浮游动物生态学研究进展[J]. 生态学报, 26(2): 576-585.
赵红雪, 邱小琮, 李力, 等. 2011. 宁夏沙湖轮虫种类与生物量及多样性变化[J]. 湖北农业科学, 50(6): 1242-1246.
朱玮阁, 李孙根, 吕林兰, 等. 2009. 环境激素氰戊菊酯和有机锡对轮虫生活史特征的影响[J]. 湖泊科学, 21(5): 687-692.
诸葛燕. 1997. 中国典型地带轮虫的研究[D]. 武汉: 中国科学院水生生物研究所.
Impact of Eutrophication on Rotifer Community Structure in Shallow Lakes
ZHENG Jinxiu, CHI Shiyun, LI Dan, WANG Hongjun, HU Juxiang*
Key Laboratory of Ecological Impacts of Hydraulic-Projects and Restoration of Aquatic Ecosystem of Ministry of Water Resources, Institute of Hydroecology, Ministry of Water Resources and Chinese Academy of Sciences, Wuhan 430079, China
Shijiu Lake and Gucheng Lake are two shallow lakes in the middle and downstream areas of the Yangtze River. The lake eutrophication in the lakes has attracted wide attention. To explore the influence of eutrophication on rotifers, an investigation on rotifer community structure were carried out in normal season and dry season in 2012. At the same time water quality parameters were tested to study the relationship between the rotifers distribution and water quality including the eutrophication status. The results showed that almost of sites in Shijiu Lake were mesotrophic (Trophic Level Index, TLI<50) except that few sites in mesotrophication (TLI>50). However in dry season, both lakes were mesotrophic (TLI>50). The investigation of aquatic organism showed that 35 species of rotifers were detected in total. There were 33 species in Shijiu lake and 27 species in Gucheng lake. the species were more in normal season. The frequency of Brachionus and Trichocerca was the highest. The dominant species were Keratella cochlearis, Polyarthra vulgaris, Trichocerca sp, Pompholyx sulcata, Trichocerca sp and Brachionus sp. The average density of the rotifers was 3023.19 ind·L-1and the average biomass was 745.72 µg·L-1. The seasonal changes were obvious in the standing crop. The standing crop in the normal season was higher than in the dry season in almost of the sites. Rotifers diversity index was higher in Shijiu lake. TLI was significantly negatively related to the rotifers species number and diversity. With the increasing of nutrient loading, rotifer species and diversity decreased. Further, Canonical Correspondence Analysis(CCA) showed that the main water qualtiy factors affecting the rotifer species distribution were NH4+-N, TN,TP and CODMn. According to the sort of the relationships between the rotifer abundance and the water environment factors, 24 sites can be roughly divided into three groups. The results showed that Rotifer community structure was affected by eutrophication, and the representative species in each group had a good indication on the trophic status. The representative species in Group 1 was Keratella valga, indicating the trophic level of oligosaprobic-β mesosaprobic; the representative species of Group 2 was Brachionus calyciflorus, indicating the nutrition level of β-α mesosaprobic; the representative species of Group 3 complicated, mainly as the indicators for oligosaprobic.
rotifers; community structure; eutrophication; shallow lakes
X171.5
A
1674-5906(2014)12-1964-08
国家自然科学基金项目(51279112);中央分成水资源费项目(1261420237007);水利部948项目(201408)
郑金秀(1979年生),女,高级工程师,主要研究方向为水生态监测与评价。E-mai:surexxzh@126.com
*通信作者:E-mail:hujuxiang2005@163.com
2014-09-01
郑金秀,池仕运,李聃,汪红军,胡菊香. 富营养化对浅水湖泊轮虫种群结构影响研究[J]. 生态环境学报, 2014, 23(12): 1964-1971.
ZHENG Jinxiu, CHI Shiyun, LI Dan, WANG Hongjun, HU Juxiang. Impact of Eutrophication on Rotifer Community Structure in Shallow Lakes [J]. Ecology and Environmental Sciences, 2014, 23(12): 1964-1971.
轮虫是淡水浮游动物的重要组成部分,在水生态系统食物链中起着承上启下的作用,对维持水生态系统结构和功能有重要意义。由于轮虫个体小,世代周期短,对环境敏感,被视为水体污染的重要指示生物,常常用于水质生态学评价(Chen等,2012;Leslie等,1983)。目前,对湖泊中轮虫的研究,除了探讨种群结构与水环境因子关系(都雪等,2014;Wen等,2011),更多关注其对富营养化的响应(Tasevska等,2012;OBERTEGGER和MANCA,2011;BerzinS和Pejler,1989)。研究表明,轮虫在浮游动物中的比例随水体富营养化程度的增加而上升(Duggan等,2002)。随着湖泊营养水平的上升,轮虫的数量和生物量有逐渐增加的趋势(王庆等,2010;Yoshida等,2003)。通过分析轮虫群落结构的动态变化,可以掌握水生态系统的结构和功能状况,因此,调查研究湖泊中轮虫分布与水环境因子和富营养化状态的关系,可为水生态系统健康评价和水质监测及污染防治提供技术支撑。
长江中下游地区是我国淡水湖泊比较集中的地区,大多数为浅水湖泊,富营养化问题较突出(史小丽和秦伯强,2007)。石臼湖和固城湖位于南京市高淳县,石臼湖是安徽省当涂县与江苏省溧水县、高淳县三县间的界湖,总面积210.4 km2。固城湖与石臼湖原系湖泊洼地,古为丹阳湖,由于长期洪水挟带泥沙沉积和围垦,后分割为石臼湖、固城湖及其它诸小湖泊,其余形成河网圩区。因此,该地区水系比较发达,石臼湖分别与长江、青弋江和水阳江三大水系相连通;以长江一级支流——姑溪河与长江相连,成为国内为数不多的通江淡水湖,是长江丰水期重要的调洪储水淡水湖泊之一。固城湖则是水阳江下游一座滞蓄山洪的天然调节湖泊,面积39 km2。近几十年来,随着社会经济的快速发展,废水排放、围湖造田及过度养殖等人类活动严重威胁了石臼湖和固城湖的生态环境。这两座湖泊都有不同程度的富营养化现象(毛春梅和张文锦,2007;王荣娟和张金池,2011)。水环境恶化,富营养化加剧,将使水生生物群落结构发生变化,但目前对这两个湖泊的轮虫种群动态及其与水质关系的研究尚未见报道。因此,本研究于2012年平水期和枯水期对两个湖泊轮虫进行采集调查,分析轮虫的种类组成、现存量及多样性变化,探讨轮虫分布与水环境因子和水体营养化状态的关系,以期为湖泊的污染控制和富营养化综合治理提供科学依据。
猜你喜欢
玩具世界(2022年3期)2022-09-20 01:46:48
中外文摘(2021年9期)2021-11-11 21:23:08
江苏水利(2021年5期)2021-05-28 08:43:42
润·文摘(2021年4期)2021-05-10 02:35:28
银幕内外(2020年5期)2020-11-25 02:42:59
故事会(2020年11期)2020-06-04 12:43:51
陶山(2019年2期)2019-12-15 20:52:00
江苏水利(2018年10期)2018-10-25 02:01:56
史志学刊(2018年4期)2018-09-04 09:22:14
中国水利(2015年16期)2015-02-28 15:14:46
