
鲂属4种鱼类种间杂交的初步研究
2014-02-15 08:01张大龙杜睿聂竹兰杨振华罗伟易少奎高泽霞王卫民陈柏湘
大连海洋大学学报 2014年2期
张大龙,杜睿,聂竹兰,杨振华,罗伟,易少奎,高泽霞,王卫民,陈柏湘
(1.华中农业大学 水产学院 农业动物遗传育种教育部重点实验室 农业部淡水生物繁育重点实验室,湖北 武汉430070;2.湖北百容水产良种有限公司,湖北 黄冈438800)
鲂属Megalobrama 隶属于鲤形目、鲤科、鲌亚科,主要分布于中国、俄罗斯,越南的一些地区也有分布,是一群较大型的经济鱼类。其中团头鲂Megalobrama amblycephala 性温顺、易起捕、适应性强、疾病少,是中国主要淡水养殖鱼类之一;三角鲂Magalobrama terminalis 食性杂、生长快、体大肉厚、骨刺较少、肉质嫩滑,是淡水鱼类中的珍品;广东鲂Megalobrama hoffmanni 肉细嫩、味鲜美,富含脂肪,为名贵食用鱼;厚颌鲂Megalobrama pellegrini可食比例大、生长较快,尤以1 ~3 龄生长最快,深受人们的喜爱。
杂交是最经典的育种方法之一,可以显著地扩大和丰富鱼类育种的基因库,促进种间基因的交流,引入异种的有利基因,从而培养出新变异品种,特别是远缘杂交。远缘杂交虽有显著的优势,但其杂交后代能形成正常胚胎并顺利达到鱼种阶段的不多。据报道,中国淡水鱼类属间以上的杂交后代有较高的成活率,且有一定生长优势,目前在生产上应用的有4 个杂交组合,即鳊鲂杂种、鲢鳙杂种、鲤鲫杂种和鲮与湘华鲮杂种[1]。有关鲂属及鲂属以上的杂交报道主要有:
(1)目间杂交:团头鲂♀与鳜♂[2];
(2)亚科间杂交:草鱼♀× 团头鲂♂[3-4]、草鱼♀×三角鲂♂[5-7]、青鱼♀×三角鲂♂[8-9]、鲢♀× 团头鲂♂及其反交[10-11]、鲤♀× 团头鲂♂[12-13]、鲢♀×三角鲂♂[14];
(3)属间杂交:长春鳊♀×三角鲂♂[15]、团头鲂♀×长春鳊♂[16-17]、红鳍鲌♀×团头鲂♂[14];
(4)种间杂交:广东鲂♀×团头鲂♂[18-19]、三角鲂♀×团头鲂♂及其反交[20]。
本研究中对鲂属鱼类进行杂交试验,旨在获得生长或其他经济性状上具有杂交优势的后代,以期从中选择合适的杂交组合应用于生产和育种实践。
1 材料与方法
1.1 材料
团头鲂为2009年6月18日从湖北鄂州梁子湖引进的原种;三角鲂为2011年12月13日从浙江钱塘江原种场引进的原种;厚颌鲂为2011年11月4日从泸州龙溪河引进的原种;广东鲂为2011年9月1日从西江肇庆引进的原种。将鲂属4种鱼的原种驯养于湖北鄂州养殖基地,于2012年6月挑选4种鱼的亲本进行人工繁殖。
1.2 方法
1.2.1 催产、人工授精和孵化 采用二针次肌肉注射法:第一针只对雌鱼注射促黄体素释放激素A2(LHRH-A2),剂量为1 μg/kg(体质量),注射时间为20:00;翌日8:00 注射第二针,雌鱼注射LHRH -A24 μg/kg(体质量)、绒毛膜促性腺激素(HCG)500 U/kg(体质量)、地欧酮(DOM)5 mg/kg(体质量),雄鱼减半注射。注射后将雌、雄亲本放在不同水泥池中流水刺激。
采用人工干法授精获得杂交受精卵,全部交配组合设计如表1所示。受精卵用黄泥浆脱黏后,于孵化缸中孵化;另取部分脱黏后的受精卵在玻璃平皿内孵化,以便观察胚胎的发育过程及计算受精率、孵化率和成活率。

表1 鲂属鱼类交配设计Tab.1 Mating design in Megalobrama species
1.2.2 受精率、孵化率、成活率的计算 取每个组合的1000 粒受精卵,分别放入直径为150 mm的培养皿中,每个组合设3 个平行,再放入50 L的塑料槽中静水孵化,用气石微充气,在原肠晚期统计各组合培养皿中的卵总数和受精卵数;剔除未受精的卵,继续孵化受精卵;根据受精卵数、活胚数、破膜鱼苗数和平游鱼苗数,取其平均值,按下式计算受精率、孵化率和成活率:
受精率=受精卵数/总卵数×100%,
孵化率=活胚数/受精卵数×100%,
成活率=平游鱼苗数/破膜鱼苗数×100%。
1.2.3 子代的培育及养殖 各组合子代在室内同一水泥池的不同孵化桶内孵化,水泥池中装有相同型号和功率的水泵,每个水泵出水口分成4 支,分别泵入4 个孵化桶中,孵化桶的上沿加装滤网,形成循环流水,各孵化桶中的水温、pH 均相同,各孵化桶中的流速和水位尽可能相同,孵化水温为26 ~28 ℃。出膜2 ~3 d 后开口摄食,每天9:00、5:00各投喂经50 目纱布过滤的熟蛋黄水,1 个蛋黄喂食10 万尾鱼苗;出膜4 d 后,将鱼苗移至室内循环流水水泥池(长、宽、高分别为5、1、0.5 m)中,进出水口加装滤网,每天9:00、5:00各投喂1次轮虫、枝角类和桡足类等浮游生物,投喂前吸底,并加注曝气后的新水;两星期后,将各组合子代转入室内循环流水的塑料槽(塑料槽直径、高分别为2.0、1.5 m)中,进出水口加装滤网,每天9:00、13:00、17:00、21:00 定时定量投喂麸皮、米糠,随后用麸皮与高龙特种水产饲料混合投喂,期间每3天吸底一次,在蓄水槽中加注曝气后的新水。在子代的培育及养殖过程中,各循环系统水温均为27 ℃。
1.2.4 子代生长性状的测量 当各组合子代生长到45日龄时,从每个组合子代群体中随机取60 尾个体,用游标卡尺测量体长、体高(精确到0.01 mm),用电子天平称量体质量(精确到0.0001 g)。根据Griffing[21]的观点,采用杂种潜力(hybrid potence,hp)判断杂种优势是否产生,计算公式如下:

其中:F1'、F2'是正反交杂种后代表型值;P1、P2是双亲表型值。当hp≥1.0 时,杂交产生了显著的杂种优势;当hp≤-1.0 时,杂交产生了杂种衰退;当-1.0<hp<1.0 时,杂交效果不明显,杂交没有产生显著的杂种优势,也没有产生杂种衰退。
1.3 数据处理
采用Excel 2007 计算平均值、标准差,用SPSS 13.0 软件进行方差分析和多重比较。
2 结果
2.1 受精率、孵化率和成活率
从表2可见:鲂属各组合中,T♀×T♂的受精率和成活率均最高,分别为89.22%、82.67%,S♀×S♂的孵化率最高,为88.51%;而S♀×T♂的受精率最低,为42.12%,H♀×H♂的孵化率最低,为54.64%,T♀× G♂的成活率最低,为66.30%。

表2 鲂属鱼类不同交配组合的受精率、孵化率和成活率Tab.2 Fertilization,hatching,and survival rates in different mating groups of Megalobrama %
2.2 子代早期生长发育
经45 d 人工养殖后,各组合子代的体长、体质量、体高的杂种潜力如图1所示,生长性状如表3所示。从图1可见:团头鲂与厚颌鲂正反杂交子代在体长与体质量方面均表现出明显的杂种潜力,而体高的杂种潜力不明显;团头鲂与三角鲂正反杂交子代在体长、体质量和体高方面杂种潜力均不明显。因广东鲂母本催产失败,未计算团头鲂与广东鲂正交以及三角鲂与广东鲂正交子代的杂种潜力。
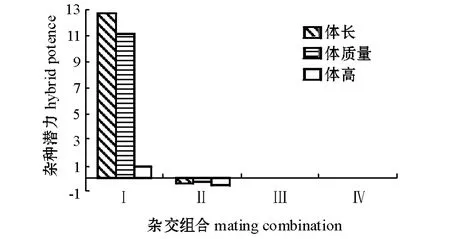
1 鲂属鱼类杂交子代早期生长性状的杂种潜力(n=60)Fig.1 Hybrid potence of the offsprings from the crossbreed of Megalobrama species(n=60)
从表3可见:T ♀× G ♂ F1的体长最大(2.931 cm),H♀× T♂F1的体长次之(2.701 cm),两组间有显著性差异(P<0.05),均显著高于其他组合(P<0.05),而T♀×S♂F1的体长最小(0.597 cm);T♀×G♂F1的体质量最大(0.870 g),H ♀× T♂F1的体质量次之(0.281 g),两组间有显著性差异(P<0.05),均显著高于其他组合(P<0.05),而T♀×S♂F1的体质量最小(0.060 g);H ♀× T ♂F1的体高最大(0.748 cm),显著高于其他组合(P<0.05),T♀×G♂F1的体高次之(0.411 cm),而T♀×S♂F1的体高最小(0.155 cm)。总体上,T♀×G♂、H♀×T♂两组合F1的生长性状均较好,而T♀×S♂、S♀×S♂F1的生长性状较差。
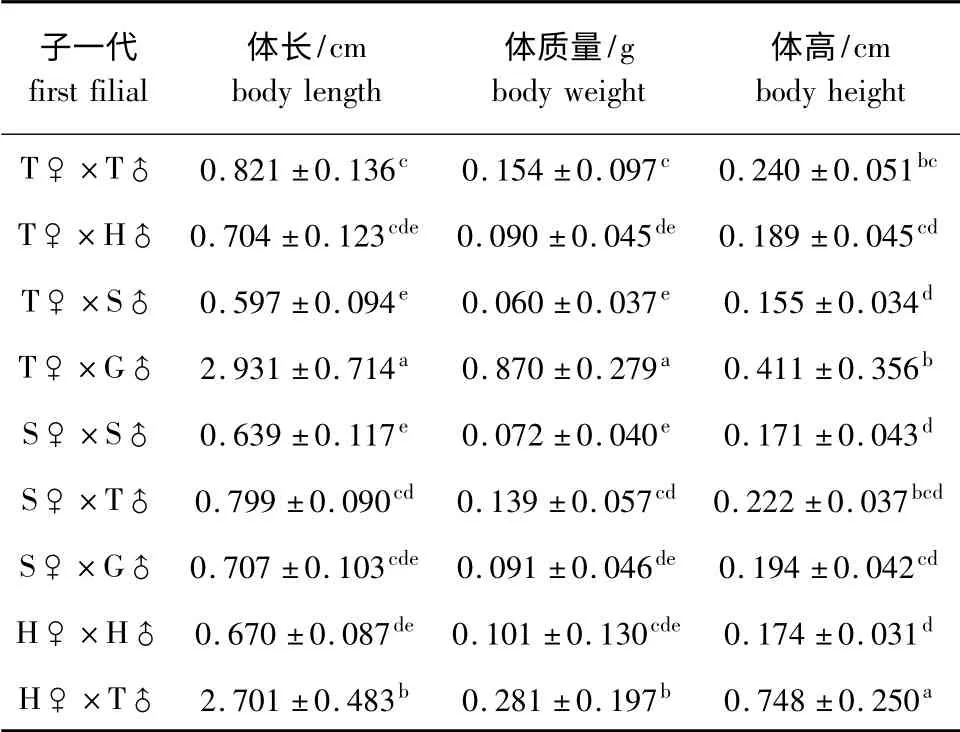
表3 鲂属鱼类杂交子代45日龄时的生长性状(n=60)Tab.3 Growth in 45-day-old F1 of Megalobrama species(n=60)
3 讨论
3.1 影响鲂属鱼类受精率、孵化率和成活率的因素
团头鲂自然分布于长江中游附属湖泊,为本地种。三角鲂分布较广,长江、黄河、黑龙江水系均有分布,而一般广布种鱼类对繁殖生态要求较低,所以,三角鲂和团头鲂的自交及团头鲂♀×三角鲂♂正交的受精率、孵化率、成活率与团头鲂♂×三角鲂♀反交的孵化率和成活率均高于其他组合,但团头鲂♂×三角鲂♀反交的受精率在所有组合中最低,这是由反交组合中三角鲂♀活力低、体质差所致。团头鲂♂×三角鲂♀反交组合中,三角鲂♀催产后立即死亡,卵子质量也较差,人工受精15 h后,受精卵大量滋生水霉。
广东鲂仅雄鱼催熟成功,由雄鱼催产成功,可以排除雌鱼对催产剂不敏感的因素。解剖广东鲂雌鱼发现,其腹壁肌肉厚,性腺呈肉红色带状,切片观察为Ⅱ、Ⅲ时相的卵母细胞,证明性腺发育不成熟,但因肝脏脂肪偏多,外观腹部膨大,被误认为成熟而催产。广东鲂自然分布于韩江、珠江和海南岛水系,水温高,根据生活史策略与环境统一的习性,广东鲂繁殖有效积温相对较高,而湖北省鄂州因受梅雨天气的影响,未能达到广东鲂的有效繁殖积温。
厚颌鲂为长江上游特有鱼类,在进化过程中已经适应长江上游流水生活环境。长江上游地区是西部高原向东部平原过渡的区域,环境条件和下游江段存在差异,因而厚颌鲂生理结构和生态习性表现出对流水环境高度适应和依赖,生活史过程必须在流水环境中完成。本试验中由于条件有限,不能模拟出类似自然流水的产卵条件,这便解释了厚颌鲂自交及厚颌鲂杂交组合受精率、孵化率和成活率都较低的现象。王剑伟等[22]进行了厚颌鲂繁殖试验,结果表明,平均受精率为62.1%,孵化率为44.7%,与本试验结果相近。
虽然本次对鲂属4种鱼类自交与杂交试验均在相同条件下进行人工干法授精,且孵化条件亦相同,但因4种鱼类在自然环境中的生活条件各异,其最适孵化培育条件还有待进一步研究,这也影响鲂属鱼类自交与杂交组合的受精率、孵化率和成活率。
3.2 鲂属鱼类杂交可行性的研究
种间杂交的成功率取决于精卵的亲合能力和精卵的质量。本试验中,4种鲂在分类上为同属不同种,其中团头鲂、三角鲂、广东鲂的染色体数目均为2n=48[17-18],这是三者杂交的细胞学基础,说明在理论上两两杂交是可能的。本试验中属于鲂属种间的远缘杂交,采用正反交的方法均能获得杂交后代,F1胚胎发育、仔鱼开口、鱼苗生长都正常,说明正反交的杂交相容性均较高。从正反交的繁殖生物学数据可见,以团头鲂和三角鲂为母本的杂交组合成功率明显高于以厚颌鲂为母本的组合。李思发等[23]从形态学以及生物遗传学方面指出,团头鲂与三角鲂差异小,亲缘关系最近;广东鲂与团头鲂、三角鲂差异较大,亲缘关系较远。本试验结果也能反映出这方面的差异。
由于异源精子亲合度小于同源配子的亲合度,所以本试验中杂交组合的受精率相比自交组(团头鲂、三角鲂、厚颌鲂自交)的受精率要低,但也能获得相对自交组合而言较高的受精率、孵化率、成活率,这说明4种鲂进行杂交虽然存在繁殖障碍,但是繁殖障碍较小。谢刚等[24]利用广东鲂♀×团头鲂♂杂交获得了成活率高且可育的后代。综上所述,鲂属鱼类种间杂交是可行的。
3.3 鲂属鱼类杂交与自交F1 的早期生长发育
远缘杂交在水产养殖上的应用非常广泛,许多远缘杂交的组合都具有明显的杂种优势。鲂属鱼类的杂交与自交F1,在人工循环流水养殖条件下养殖45 d 时,部分组合F1的生长性状之间表现出一定的差异,如T♀×T♂、T♀×S♂、T♀×G♂、H♀×T♂4 个组合F1的体长、体高、体质量之间均存在显著性差异(P<0.05);部分组合F1的生长性状之间差异不显著,如T♀× H♂、T♀× S♂、S♀×S♂、S♀×G♂、H♀×H♂5 个组合F1的体质量之间差异不显著(P >0.05);团头鲂与厚颌鲂正反交表现出较高的杂种优势(图1)。亲本的优良显性基因的互相作用和群体中杂合子频率的增加,抑制和减弱了不良基因的作用,提高了整个群体的平均显性效应与上位效应。但在极少数情况下,因某些等位基因间的相互作用会产生负的显性效应,在个别性状上表现出杂种群体均值低于双亲均值或双亲中任何一方的现象,即所谓的“劣势”[25]。本试验中,所有杂交组合的子代养殖条件相同,但团头鲂与三角鲂正反交的杂种优势不明显,也未出现所谓的“劣势”,可能是测量其F1时日龄较短、个体较小,以及所投饲料、养殖密度和投饵量均不是最佳等原因所致。因广东鲂母本催产失败,团头鲂与广东鲂正反交及三角鲂与广东鲂正反交F1是否有杂种优势还有待进一步研究。
[1]楼允东,李小勤.中国鱼类远缘杂交研究及其在水产养殖上的应用[J].中国水产科学,2006,13(1):151 -158.
[2]长江水产研究所.家鱼人工繁殖技术[M].北京:农业出版社,1973:83.
[3]长江水产研究所育种室,武汉大学生物系动物教研室.几种经济鱼类及其杂种染色体的初步研究[J].淡水渔业,1975(2):11 -13.
[4]长江水产研究所.草鱼(雌)和武昌鱼(雄)杂交种[J].科技简报,1972(20):39 -40.
[5]刘思阳,李素文.三倍体草鲂杂种及其双亲的红细胞(核)大小和DNA 含量[J].遗传学报,1987,14(2):142 -148.
[6]刘思阳.三倍体草鲂杂种及其双亲的细胞遗传学研究[J].水生生物学报,1987,11(1):52 -58.
[7]刘思阳.草鱼卵子和三角鲂精子杂交的受精细胞学研究[J].水产学报,1987,11(3):225 -232.
[8]陈淑群.青鱼(♀)和三角鲂(♂)不同亚科之间的杂交研究Ⅰ:青鱼(♀)和三角鲂(♂)及其子一代的比较细胞遗传学研究[J].湖南师范大学学报:自然科学版,1984(4):71 -80.
[9]刘筠,陈淑群,王义铣.三角鲂(Megalobrama terminalis)精子与青鱼(Mylopharyngodon piceus)卵子的受精细胞学研究[J].湖南水产科技,1982(4):3 -9.
[10]潘光碧.鲢鲂杂交鱼的初步研究[J].淡水渔业,1987(1):17-19.
[11]朱蓝菲,桂建芳,梁绍昌,等.鲢的远缘杂交子代和人工三倍体的同工酶表达[J].水生生物学报,1993,17(4):293-297.
[12]金万昆,朱振秀,王春英,等.散鳞镜鲤(♀)与团头鲂(♂)亚种间杂交获高成活率杂交后代[J].中国水产科学,2003(2):159.
[13]金万昆,朱振秀,王春英,等.框鳞镜鲤(♀)与团头鲂(♂)杂交及其杂种F1的形态学特征[J].淡水渔业,2003,33(5):16 -18.
[14]长江水产研究所,厦门水产学院.两年来淡水养殖鱼类优良品种选育和基础理论研究简况[J].动物学杂志,1975(1):43 -45.
[15]山西大学生物系,太原市农林水利服务站.鳊鲂人工杂交的初步研究[J].淡水渔业,1973(5):6 -9.
[16]广东省佛山地区水产局.团头鲂长春鳊杂交种[J].水产科技情报,1975(4):18 -19.
[17]林义浩.长春鳊、团头鲂及其杂种染色体组型的比较[J].动物学研究,1984,5(S2):65 -66.
[18]叶星,谢刚,许淑英,等.广东鲂(♀)×团头鲂(♂)杂交子一代及其双亲同工酶的比较[J].上海水产大学学报,2001,10(2):118 -122.
[19]叶星,谢刚,祁宝伦,等.广东鲂(♀)×团头鲂(♂)杂交一代及其双亲染色体组型的分析[J].大连水产学院学报,2002,17(2):102 -107.
[20]杨怀宇,李思发,邹曙明.三角鲂与团头鲂正反交F1的遗传性状[J].上海水产大学学报,2002,11(4):305 -309.
[21]Griffing B.Use of a controlled-nutrient experiment to test heterosis hypotheses[J].Genetics,1990,126:753 -756.
[22]王剑伟,谭德清,李文静.厚颌鲂人工繁殖初报及胚胎发育观察[J].水生生物学报,2005,29(2):130 -136.
[23]李思发,朱泽闻,邹曙明,等.鲂属团头鲂、三角鲂及广东鲂种间遗传关系及种内遗传差异[J].动物学报,2002,48(3):339 -345.
[24]谢刚,叶星,庞世勋,等.杂交(广东鲂♀×团头鲂♂)及其亲本主要遗传性状的比较研究[J].湖北农学院学报,2002,22(4):330 -332.
[25]刘小林,常亚青,相建海,等.栉孔扇贝不同种群杂交效果的初步研究.Ⅰ:中国种群与俄罗斯种群的杂交[J].海洋学报,2003,25(1):93 -99.
猜你喜欢
国外畜牧学(猪与禽)(2022年2期)2022-07-01
中国饲料(2022年5期)2022-04-26
国外畜牧学·猪与禽(2022年1期)2022-04-23
农业科技与装备(2017年9期)2018-03-02
河南水产(2017年2期)2017-06-19
意林(2017年3期)2017-03-03
广东饲料(2016年3期)2016-12-01
现代畜牧科技(2015年12期)2015-10-21
民间故事选刊·下(2015年4期)2015-05-19
小小说月刊·下半月(2015年3期)2015-05-14
