
S-亚硝基-N-乙酰半胱氨酸对绿脓菌素感染鼠体内的病理形态及氧化作用的影响
2014-02-08 06:26柴文戍
中国全科医学 2014年9期
刘 伟,柴文戍
铜绿假单胞菌是机体免疫力低下者急性院内感染的重要原因,也是肺纤维化或支气管扩张患者慢性或反复肺感染的最常见原因[1]。绿脓菌素(PCN)是铜绿假单胞菌产生的毒力因子,对哺乳类细胞有很多破坏性作用。研究证明了PCN通过产生活性氧(ROS)对内皮细胞[2]和上皮细胞[3]产生氧化损伤作用。一氧化氮(NO)是哺乳类动物宿主防御的重要成分,是独特的抗菌剂,NO抑制各种微生物的生长,包括病毒、细菌、寄生虫和真菌[4]。NO衍生的活性分子,如过氧亚硝基(ONOO-),可以诱导病原体氧化损伤。研究证明NO在缺血再灌注损伤时能够防止氧化损伤,并减少哺乳动物宿主细胞的氧化压力[5]。S-亚硝基-N-乙酰半胱氨酸(SNAC)是NO供体,可以改善细胞氧化损伤。本实验主要探索PCN对感染鼠体内ROS、超氧化物歧化酶(SOD)、白介素8(IL-8)的作用及其机制,进一步探讨SNAC对此作用的影响。
1 材料与方法
1.1 材料 PCN购买于Sigma公司。实验所用动物均为无病原体的8~12周龄、体质量230~250 g的健康雄性Sprague-Dawley(SD)大鼠(辽宁医学院动物实验中心提供)。试剂盒购自南京建成生物工程研究所,N-乙酰半胱氨酸(NAC)与亚硝酸钠(NaNO2)购买于生工生物工程(上海)股份有限公司。
1.2 方法
1.2.1 动物模型制备及分组 采用随机数字表法将90只健康雄性SD大鼠分为对照组、PCN组和SNAC组,每组30只。对照组和PCN组大鼠分别腹腔注射1 ml/100 g 0.9%氯化钠溶液,SNAC组大鼠腹腔注射SNAC(1 ml/100 g)。用1%戊巴比妥钠溶液(40 mg/kg)行大鼠腹腔注射麻醉,对照组气管内接种0.9%氯化钠溶液0.2 ml,PCN组和SNAC组分别气管内接种PCN 0.2 ml(6×108CFU/ml)。
1.2.2 SNAC准备 NAC和NaNO2用于SNAC准备[6]。由等摩尔的NAC和NaNO2混合生成SNAC水溶液,室温条件下避光混合搅拌20 min。0.9%氯化钠溶液稀释SNAC到终浓度100 nmol/L并立即使用。
1.2.3 病理标本取材及处理 分别于PCN感染后6、18、30、48、72 h,用1%戊巴比妥钠溶液(40 mg/kg)进行腹腔麻醉,处死大鼠。然后分离气管,经气管注入5 ml无菌磷酸盐缓冲液(PBS)行右侧支气管肺泡灌洗,反复5次,回收后注入无菌试管,回收率≥80%。回收的支气管肺泡灌洗液(BALF)离心(离心半径 13.5 cm,2 000 r/min,10 min),取上清液保存于-20 ℃,统一进行BALF细胞总数和中性粒细胞计数。取左上肺浸泡于10%甲醛溶液中,做苏木素-伊红(HE)染色;取左肺余肺1 g加入9 ml 0.86%冷氯化钠溶液制成肺组织匀浆,离心,取上清液保存于-70 ℃,统一进行生化指标测定。
1.2.4 大体观察 分别于PCN感染后6、18、30、48、72 h观察大鼠精神状态、进食及饮水情况及肺炎症状,并观察肺组织外观颜色、水肿、出血等炎性表现及镜下观察相应时间点肺脏病理组织形态学变化。
1.2.5 肺组织ROS含量、SOD活性的测定 肺组织匀浆ROS含量及SOD活性测定按试剂盒说明书操作步骤进行。
1.2.6 肺组织IL-8含量的测定 使用酶联免疫吸附法(ELISA)测定肺组织匀浆IL-8含量,按试剂盒说明书操作步骤进行。

2 结果
2.1 大鼠的一般状态及肺组织外观 PCN感染6 h后大鼠出现精神萎靡、食欲不振、寒颤、发热、呼吸困难,感染30 h最明显。对照组大鼠肺外观呈粉红色,无明显异常改变;PCN组PCN感染后6 h时可见肺组织表面散在点状出血;18~72 h肺体积增大,肺组织暗红色,呈片状出血;SNAC组肺组织病变较PCN组明显减轻。
2.2 肺组织形态学变化 光镜下可见对照组肺组织结构清晰,肺泡间隔无增厚及炎性细胞浸润;PCN组肺组织肺泡壁结构完整性被破坏,肺泡间隔增厚,明显充血、水肿,肺泡内大量炎性细胞浸润,30 h出现典型肺炎表现;与PCN组相比,SNAC组肺组织炎性表现明显减轻,肺泡壁结构完整性较好,充血、水肿减轻,炎性细胞浸润减少(见图1)。
2.3 3组大鼠不同时间BALF细胞总数及中性粒细胞计数比较 3组大鼠PCN感染后6、18、30、48、72 h BALF细胞总数及中性粒细胞计数比较,差异均有统计学意义(P<0.05);其中PCN组和SNAC组各时间点BALF细胞总数及中性粒细胞计数较对照组均增高,SNAC组各时间点BALF细胞总数及中性粒细胞计数较PCN组均降低,差异有统计学意义(P<0.05,见表1、2)。
2.4 3组大鼠不同时间ROS含量比较 3组大鼠PCN感染后6、18、30、48、72 h ROS含量比较,差异均有统计学意义(P<0.05);其中PCN组和SNAC组各时间点ROS含量较对照组均增高,SNAC组各时间点ROS含量较PCN组均降低,差异有统计学意义(P<0.05,见表3)。
2.5 3组大鼠不同时间SOD活性比较 3组大鼠PCN感染后6、18、30、48、72 h SOD活性比较,差异均有统计学意义(P<0.05);其中PCN组和SNAC组各时间点SOD活性较对照组均降低,SNAC组各时间点SOD活性较PCN组均增高,差异有统计学意义(P<0.05,见表4)。
2.6 3组大鼠不同时间IL-8含量比较 3组大鼠PCN感染后6、18、30、48、72 h IL-8含量比较,差异均有统计学意义(P<0.05);其中PCN组和SNAC组各时间点IL-8含量较对照组均增高,SNAC组各时间点IL-8含量较PCN组均降低,差异有统计学意义(P<0.05,见表5)。

注:a对照组肺组织结构清晰,肺泡间隔无增厚及炎性细胞浸润;b PCN组肺泡壁结构完整性被破坏,肺泡间隔增厚,明显充血、水肿,肺泡内大量炎性细胞浸润;c SNAC组肺组织炎性表现减轻,肺泡壁结构完整性好,充血、水肿减轻,炎性细胞浸润减少

图1 PCN感染后30 h肺组织形态学变化(HE染色,×200)
注:与对照组比较,*P<0.05;与PCN组比较,△P<0.05
表2 3组大鼠不同时间BALF中性粒细胞计数比较
注:与对照组比较,*P<0.05;与PCN组比较,△P<0.05
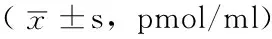
表3 3组大鼠不同时间ROS含量比较
注:与对照组比较,*P<0.05;与PCN组比较,△P<0.05

表4 3组大鼠不同时间SOD活性比较
注:与对照组比较,*P<0.05;与PCN组比较,△P<0.05

表5 3组大鼠不同时间IL-8含量比较
注:与对照组比较,*P<0.05;与PCN组比较,△P<0.05
3 讨论
本研究结果表明PCN感染后,PCN组肺组织明显充血、水肿,大量炎性细胞浸润,BALF检测到炎性细胞显著增加,以中性粒细胞为主,ROS含量增高,SOD活性降低,IL-8含量增加,说明PCN发挥毒性作用与氧化损伤有关。而SNAC干预后,SNAC组大鼠一般状态较好,肺组织炎性表现减轻,BALF检测到的炎性细胞明显减少,ROS含量减少,SOD活性增加,IL-8含量减少,表明SNAC对小鼠体内的PCN氧化损伤具有保护作用。研究还表明大鼠的一般状态、肺组织病理炎性表现在PCN感染后30 h时表现最明显。
上述表现其原因考虑为,SNAC是NO供体,NO不仅具有免疫调节作用而且直接作用于微生物,NO既可以杀死病原体又可以抑制其复制。许多机制参与氮氧化物的抗菌作用,包括改变蛋白质硫醇和金属中心的相互作用[7-8],阻塞微生物基本生理过程[9-10],破坏细菌的蛋白质铁硫集群而导致催化毒性氧化反应的游离铁的释放[11]。NO和相关物质通过扰乱参与DNA复制的锌蛋白和与酪氨酰激酶反应抑制核苷酸还原酶,进而抑制细菌DNA复制[12]。此外,线粒体是ROS的重要来源,其损伤可能导致细胞氧化损伤,SNAC能改善线粒体的功能,因此导致ROS产生减少[6],发挥其保护氧化损伤的作用。而且有报道认为在假单胞菌肺炎动物模型中,吸入NO可以减少细菌的负荷[13]。本实验结果表明PCN通过氧化应激导致细胞核组织损伤。SNAC作为NO供体,不仅具有抗菌作用,而且对氧化损伤具有一定的保护作用。
研究表明PCN主要通过介导ROS的产生造成氧化应激而发挥其毒性作用,PCN可以被NAD(P)H 或硫醇直接还原,通过有氧氧化循环产生超氧化物(O2-)和过氧化氢(H2O2)[14]。氧化损伤是氧化和抗氧化过程失衡的结果,抗氧化防御系统(如SOD)在消除体内氧化循环中起着重要作用。SOD是机体内重要的抗氧化剂,可消除过剩的氧自由基及其衍生物,保护细胞免受损伤[15-17]。IL-8为重要的中性粒细胞趋化因子,是宿主防御病原菌感染机制中起重要作用的前炎性细胞因子,对中性粒细胞具有较强的趋化作用和激活作用[18-19],IL-8释放的增加是导致感染肺部中性粒细胞增多最主要的原因。研究证据表明,氧化应激可以引发炎症,反过来炎症加剧了活性氧的产生[20];肺炎性疾病[21]、暴露于环境微粒肺细胞[22]的氧化应激诱导IL-8表达并乙酰化;氧化应激能硝基化人组蛋白脱乙酰化酶2(HDAC2),导致IL-8表达增加[23];Ivison等[24]研究表明H2O2通过磷酸化p-38 MAP酶和延长I-κB降解与核因子(NF)-κB激活的协同作用使IL-8产生增加。因此氧化应激引发IL-8表达上调可能是联系氧化应激和炎症之间的一个重要环节。
总之,本研究结果提示,作为NO供体的SNAC不仅具有抗菌作用,并可能通过影响ROS、SOD的表达而发挥一定抗氧化作用,但本实验检测的炎性因子及抗氧化酶有限,仍需实验进一步研究。
1 Muller M.Premature cellular senescence induced by pyocyanin,a redox-active Pseudomonas aeruginosa toxin[J].Free Radic Biol Med,2006,41(11):1670-1677.
2 Muller M.Pyocyanin induces oxidative stress in human endothelial cells and modulates the glutathione redox cycle[J].Free Radic Biol Med,2002,33(11):1527-1533.
3 O′Malley YQ,Abdalla MY,McCormick ML,et al.Subcellular localization of Pseudomonas pyocyanin cytotoxicity in human lung epithelial cells[J].Am J Physiol Lung Cell Mol Physiol,2003,284(2):L420-430.
4 Fang FC.Antimicrobial reactive oxygen and nitrogen species:concepts and controversies[J].Nat Rev Microbiol,2004,2(10):820-832.
5 Husain M,Bourret TJ,McCollister BD,et al.Nitric oxide evokes an adaptive response to oxidative stress by arresting respiration[J].J Biol Chem,2008,283(12):7682-7689.
6 Andraus W,Souza GF,Oliveira MG,et al.S-nitroso-N-acetylcysteine ameliorates ischemia-reperfusion injury in the steatotic liver[J].Clinics,2010,65(7):715-721.
7 Foster MW,Hess DT,Stamler JS.Protein S-nitrosylation in health and disease:current perspective[J].Trends in Molecular Medicine,2009,15(9):391-404.
8 Han G,Zippin JH,Friedman A.From bench to bedside:the therapeutic potential of nitric oxide in dermatology[J].J Drugs Dermatol,2009,8(6):586-594.
9 Gusarov I,Shatalin K,Starodubtseva M,et al.Endogenous nitric oxide protects bacteria against a wide spectrum of antibiotics[J].Science,2009,325(5946):1380-1384.
10 Akaike T,Okamoto T,Zaki MH,et al.New paradigm of host defense against intracellular pathogens by nitric oxide[J].Nihon Hansenbyo Gakkai Zasshi,2009,78(1):41-47.
11 Keyer K,Imlay JA.Superoxide accelerates DNA damage by elevating free-iron levels[J].Proc Natl Acad Sci U S A,1996,93(24):13635-13640.
12 Schapiro JM,Libby SJ,Fang FC.Inhibition of bacterial DNA replication by zinc mobilization during nitrosative stress[J].Proc Natl Acad Sci U S A,2003,100(14):8496-8501.
13 Webert KE,Vanderzwan J,Duggan M,et al.Effects of inhaled nitric oxide in a rat model of Pseudomonas aeruginosa pneumonia[J].Crit Care Med,2000,28(7):2397-2405.
14 Rada B,Jendrysik MA,Pang L,et al.Pyocyanin-enhanced neutrophil extracellular trap formation requires the NADPH oxidase[J].PLoS One,2013,8(1):e54205.
15 罗招凡,劳伟思,梁伟南,等.血清同型半胱氨酸和SOD水平与急性脑梗死的相关性研究[J].中国卫生检验杂志,2011,21(10):2463-2464.
16 高天.颅内复杂动脉瘤患者围术期血清基质金属蛋白酶-9、超氧化物歧化酶、丙二醛及一氧化氮水平变化研究[J].实用心脑肺血管病杂志,2012,20(1):52.
17 苏显明,智晓文,刘景委,等.丹红注射液对老年冠心病患者血中对氧磷酶1、超氧化物歧化酶活性及丙二醛水平的影响研究[J].中国全科医学,2011,14(11):3804.
18 Kunkel SL,Standiford T,Kasahara K,et al.Interleukin-8(IL-8):the major neutrophil chemotactic factor in the lung[J].Exp Lung Res,1991,17(1):17-23.
19 程刚,陆士奇,李岩.急性胰腺炎患者血清降钙素原和肿瘤坏死因子α及白介素8的动态变化及临床意义[J].中国全科医学,2012,15(9):3180.
20 Zhou J,Jang YP,Kim SR,et al.Complement activation by photooxidation products of A2E,a lipofuscin constituent of the retinal pigment epithelium[J].Proc Natl Acad Sci U S A,2006,103(44):16182-16187.
21 Ito K,Ito M,Elliott WM,et al.Decreased histone deacetylase activity in chronic obstructive pulmonary disease[J].N Engl J Med,2005,352(19):1967-1976.
22 Gilmour PS,Rahman I,Donaldson K,et al.Histone acetylation regulates epithelial IL-8 release mediated by oxidative stress from environmental particles[J].Am J Physiol Lung Cell Mol Physiol,2003,284(3):L533-540.
23 Ito K,Hanazawa T,Tomita K,et al.Oxidative stress reduces histone deacetylase 2 activity and enhances IL-8 gene expression:role of tyrosine nitration[J].Biochem Biophys Res Commun,2004,315(1):240-245.
24 Ivison SM,Wang C,Himmel ME,et al.Oxidative stress enhances IL-8 and inhibits CCL20 production from intestinal epithelial cells in response to bacterial flagellin[J].Am J Physiol Gastrointest Liver Physiol,2010,299(3):G733-741.
本文链接
铜绿假单胞菌是一种定植于支气管扩张、肺囊性纤维化、慢性阻塞性肺疾病患者下呼吸道的机会致病菌,可持续刺激宿主免疫反应而致进展性的气道破坏,具有多重耐药的特点。一旦感染,临床治疗十分困难,现已成为院内感染的主要致病菌。
绿脓菌素是铜绿假单胞菌分泌的重要的细菌毒力因子之一,可降低支气管黏液流动速度,有利于铜绿假单胞菌定植于呼吸道;同时,它还逐步降低呼吸道纤毛摆动,最后可发展为广泛的纤毛停滞,进而导致上皮损伤。
猜你喜欢
江苏卫生保健(2022年7期)2022-07-26
昆明医科大学学报(2022年1期)2022-02-28
现代临床医学(2021年6期)2021-11-20
天津医科大学学报(2021年4期)2021-08-21
现代临床医学(2021年4期)2021-07-31
皮肤病与性病(2021年3期)2021-07-30
中华养生保健(2020年10期)2021-01-18
家庭影院技术(2018年9期)2018-11-02
中国制笔(2017年2期)2017-07-18
中成药(2017年3期)2017-05-17
