
长链非编码RNA NEAT1对小鼠精子发生的影响
2014-01-20 08:38:38曾文先
家畜生态学报 2014年12期
胡 源,曾文先
(西北农林科技大学动物科技学院,陕西杨凌712100)
长链非编码RNA(lncRNA)具有在细胞特异性表达的特性[1],参与形成细胞核中的亚结构并调节多种细胞活动和生物学过程,包括调控基因表达和蛋白质的活性、干细胞自我更新与分化等[2]。但关于lncRNA 在小鼠精子发生过程中的作用及其调节机理目前尚未见报道。lncRNA NEAT1作为核心组分参与形成旁斑(paraspeckle)结构[3]。敲除NEAT1影响DBHS(Drosophila melanogaster behavior,human splicing)蛋白在细胞中的定位,从而影响旁斑结构的形成[4-5]。而旁斑可以阻滞成熟mRNA 的出核与后续翻译[6],参与调控细胞的分化和生长[7,8]。在细胞分化的过程中NEAT1的表达升高[9-11]。本研究旨在探索NEAT1对维持小鼠精子发生所起的作用,为雄性动物的精子发生过程和生殖研究提供试验基础。
1 材料与方法
1.1 试验动物及细胞系
试验动物:C57BL/6J小鼠,昆明种小白鼠。细胞系:GC-1精原细胞系,293T 细胞系。病毒包装系统:CD513B-U6表达质粒及包装质粒(pVSV-G、pGag-Pol、pRsv-rev)。
1.2 主要试剂与仪器
PCR 扩增仪;凝胶电泳成像分析系统(Biorad);二氧化碳培养箱(Thermo);超净工作台(苏州);台式低温离心机(Sigma);超纯水制备系统(Mill-Q);立式压力蒸汽灭菌锅;荧光倒置显微镜(Olympus),二氧化碳培养箱(Thermo)。
1.3 NEAT1表达检测
RNA 提取:使用Trizol法提取细胞总RNA;反转录PCR:使用TaKaRa 公司PrimeScriptTMRT reagent Kit with gDNA Eraser试剂盒;PCR:使用TaKaRa公司Ex Taq®试剂盒,引物信息如下:
上游引物:TACATGGGGGCAGTGTCCTA
下游引物:ACCACAGAAGAGGAAGCACG
1.4 干扰载体构建
包括设计shRNA;CD513B-U6质粒双酶切:使用BamHI和EcoRI限制性内切酶进行双酶切;构建CD513B-U6-shRNA 载体及测序。

表1 NEAT1的shRNA 序列信息Table 1 The shRNA sequence information of NEAT1
1.5 慢病毒包装与滴度测定
使用 CD513B-U6-shRNA 载体、pVSV-G1、pGag-Pol、pRev ug包装载体,转染293T 细胞系进行病毒包装;采用倍比稀释法测定慢病毒的滴度,用荧光显微镜观察每孔中发出绿色荧光的细胞的数量,对最后两个有绿色荧光细胞的孔进行细胞计数,数目依次记为X,Y,公式为:
滴度(TU/mL)=(X+Y×10)×1000/2X 孔的病毒溶液的含量(μL)。
1.6 有精子发生的曲细精管比例统计
使用多聚甲醛固定小鼠的睾丸,石蜡包埋切片后进行 H.&E.染色。依据Kanatsu-Shinohara等[12]采用的方法,当生殖细胞在曲细精管的管基底膜呈圆周状分布,并且生殖细胞有两层或者两层以上时,判定该曲细精管有精子发生;曲细精管内生殖细胞没有达到两层的,则判定为无精子发生。精子发生比例:有精子发生精细管占总体曲细精管的比例。每个处理组设置三个重复,每个重复观察平均约50个曲细精管。
2 结果与分析
2.1 NEAT1在小鼠睾丸及细胞系中的表达
提取C57BL/6J小鼠的睾丸组织和GC-1细胞的RNA,反转录PCR 后用cDNA 模板进行PCR 检测NEAT1的表达分布,结果表明NEAT1 在成年小鼠睾丸以及GC-1细胞系中均表达(图1)。
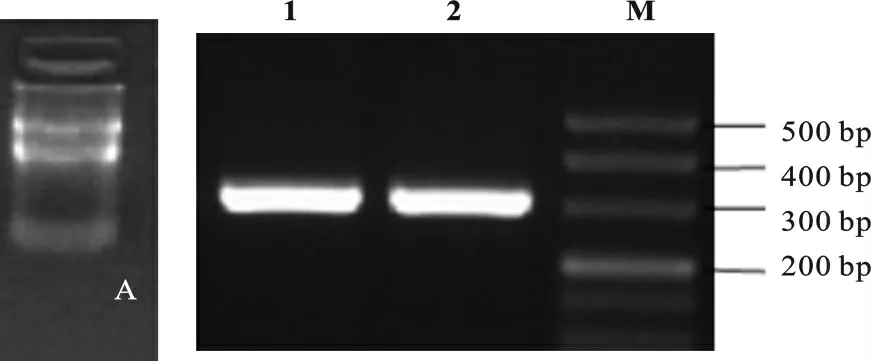
图1 NEAT1在睾丸组织和GC-1细胞中的表达A.提取的RNA 电泳图片;B.RT-PCR 结果;1.GC-1细胞系;2.睾丸组织Fig.1 NEAT1expression in testicular tissue and GC-1cellsA.Bxtracted RNA electrophoresis;B.RT-PCR resuts;1.GC-1cells;2.Testicular tissues
2.2 干扰载体构建结果与分析
将干扰shRNA 和双酶切后的CD513B-U6 载体连接构建干扰载体。质粒送公司测序,将结果和设计的shRNA 进行比对,连接成功且未出现碱基突变(图2),表明NEAT1干扰载体构建成功。
2.3 慢病毒包装及病毒滴度测定
利用SBI的第三代慢病毒包装系统,将四个质粒共转染293T细胞系包装病毒,16h后进行观察并换成病毒包装缓冲液,结果发现有约70%的293T 细胞能够自发荧光,表明转染成功(图3)。48h后收毒,并进行滴度测定,测定结果为1.72×107TU/mL。
图2 测序结果分析Fig.2 The analysis of sequencing results
2.4 干扰效果检测结果及分析
将构建的慢病毒感染GC-1 细胞,荧光显微镜下观察发现病毒感染细胞效果良好(图4)。
提取GC-1细胞RNA,反转录PCR 后实时定量PCR 检测NEAT1的表达,结果显示包装的干扰慢病毒能够将NEAT1转录本水平下调69%,干扰效果良好(图4)。

图3 四质粒共转染293T 细胞,16h后荧光图左侧为NEAT1-shRNA 组,右侧为NC组Fig.3 The Fluorescent images at 16hours after the transfection of four plasmids into 293Tcell lineLeft means CD513B-U6-NEAT1-shRNA,Right means CD513B-U6-NC

图4 慢病毒感染GC-1细胞效果A,B,C分别为明场图,荧光图和叠加图Fig.4 The efficiency of lentivirus infect GC-1cellsA,B,C are BFI,fluorography and overlay chart
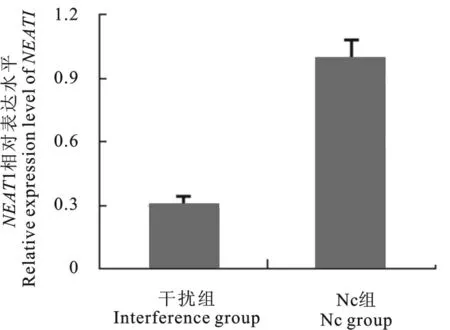
图5 在GC-1细胞中检测RNAi干扰效果Fig.5 RNA interference of NEAT1in GC-1cells
2.5 睾丸指数检测结果及分析
统计并对比干扰组,NC 组和空白组(未注射病毒)小鼠睾丸指数,结果显示干扰病毒注射组睾丸指数大于另外两组,表明睾丸的重量有轻微增加的趋势,但统计结果不显著(P=0.11)(图6)。
2.6 NEAT1干扰病毒注射对精子发生的影响
将干扰和NC组病毒通过睾丸输出管注射到曲细精管中,35d后固定干扰病毒注射组和NC 组小鼠睾丸组织并进行H.&E.染色。对比不同组的H&E 染色图发现注射NEAT1 干扰病毒后小鼠(相对于NC 组)睾丸中有精子发生的曲细精管减少,部分管腔内生殖细胞变少,表明干扰NEAT1可能影响小鼠精子发生(图7)。
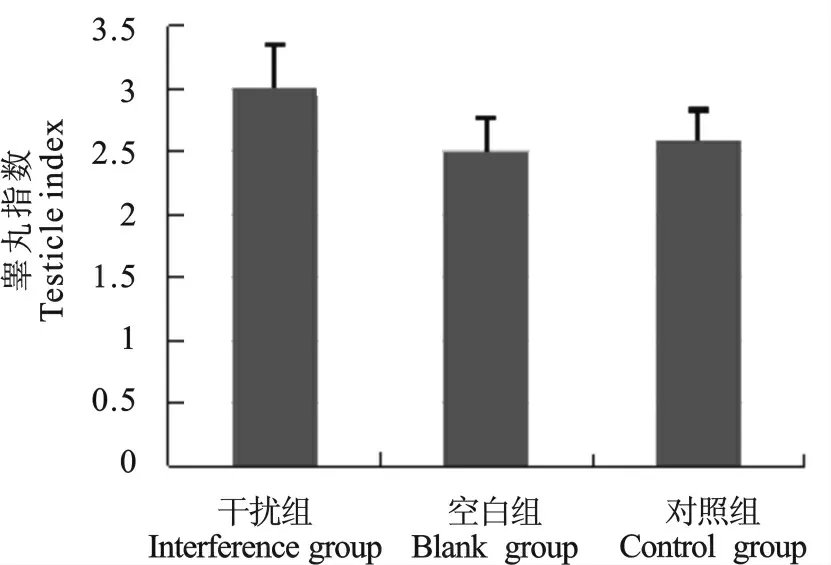
图6 干扰组、空白组和对照组的睾丸指数Fig.6 The ratio of testicles in interfering group,blank group and control group
2.7 有精子发生的曲细精管统计及分析
使用Kanatsu-shinohara的标准统计干扰组和NC组的小鼠睾丸内有精子发生的曲细精管比例,结果显示NC 组有精子发生的曲细精管的比例为97%,而NEAT1干扰组有精子发生的曲细精管比例为86%,明显减少(P<0.05)(图8),小鼠睾丸曲细精管内精子发生过程受到影响。上述结果表明NEAT1参与维持小鼠睾丸的精子发生。
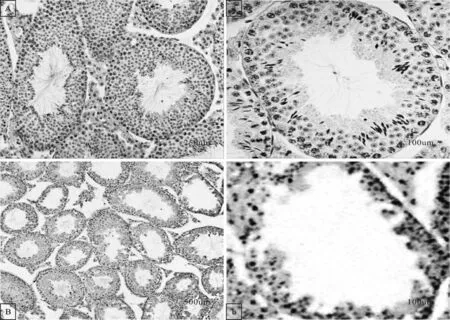
图7 H&E染色图A,a为NC组,B,b为干扰组Fig.7 The images of H&E stainingA,a are NC group,B,b are interference group
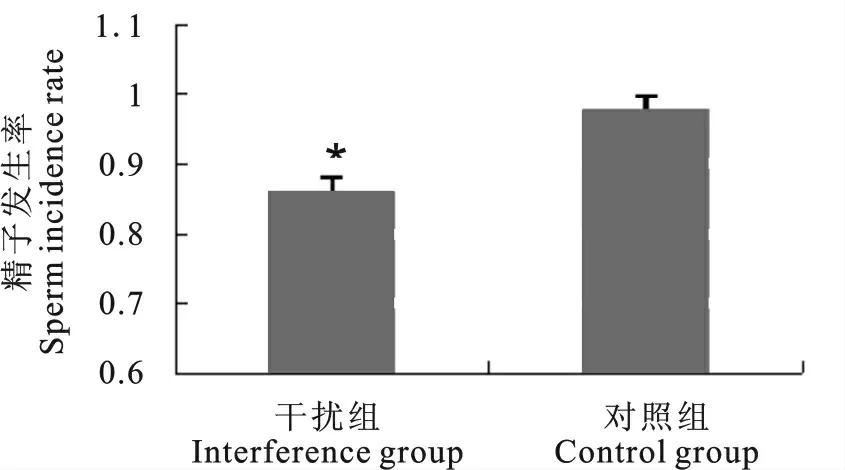
图8 干扰组与对照组精子发生比例Fig.8 The spermatogenesis ratio comparison between two groups
3 讨论
在2010年,Archa等[13]研究结果表明NEAT1在人类胚胎干细胞中表达,且能作为核心组分形成旁斑。在人类疾病研究方面,NEAT1 参与了一些重大疾病的发病过程,如卵巢癌,银屑病等[14-15]。而NEAT1在小鼠精子发生过程中的作用对于目前尚未见报道。本试验则验证了小鼠NEAT1在睾丸和GC-1细胞系中表达并采用干扰的方法探究在小鼠中NEAT1对精子发生的影响。
试验结果表明,NEAT1参与调节小鼠精子发生这一重要生物学过程。在本试验中,干扰NEAT1后试验组小鼠的睾丸指数轻微增加(相对与注射NC病毒和空白对照睾丸的重量),但统计数据分析发现差异并不显著,不足以证明NEAT1对小鼠的睾丸重量有明显的影响。2011年,Nakagawa等[16]研究发现,NEAT1敲除小鼠可以在试验条件下生存且具有繁殖能力,除了无法形成旁斑结构外,并未表现出明显的表型。这与本研究得到的试验结果基本一致。小鼠精子发生数据统计结果表明,注射了干扰NEAT1病毒组(与NC病毒注射组相比)的精子发生比例有下调,其分别是86%和97%。该结果暗示了NEAT1有可能具有维持小鼠睾丸中精子发生的作用,NEAT1 对小鼠的生殖过程有一定的影响,但小鼠仍然具有生殖能力。
目前,在人的胚胎干细胞里的研究结果显示,NEAT1发挥功能的机制主要是依赖RNA-蛋白复合体结构。研究表明,在未分化的人源胚胎干细胞中,由于缺乏NEAT1 和paraspeckles,一些对于干细胞多能性维持具有重要作用的mRNA,例如Lin28可以在转录之后快速出核,翻译产生干细胞维持所需要的LIN28蛋白质,在分化过程中,其通过旁斑这一亚核结构将Lin28 mRNA 滞留在亚核区,阻碍其翻译表达,表明NEAT1具有调控人源胚胎干细胞的命运决定的重要作用[6]。而在小鼠的精子发生过程中,包括SSCs的自我更新与分化和后期的精原细胞分裂等过程都涉及到很多相关基因的时序性表达。比如在SSCs自我更新过程中起重要作用的Lin28(维持干细胞特性)、PLZF(SSCs中重要干细胞转录因子,不受GDNF 通路调控)、Bcl6b(维持SSCs自我更新周期,干扰Bcl6b会促进小鼠SSCs的凋亡,敲除则会造成Bcl6b小鼠其曲细精管内不同阶段生殖细胞的比例出现异常)、Etv5(敲除Etv5会导致SSCs功能受损,曲细精管内只含有支持细胞,雄性不育)等[17-19]。这些重要基因是否会被NEAT1构成的旁斑结构进行表达水平调控或者通过旁斑对mRNA 的滞留效应进行时序调控,有待进一步试验验证。另外,Nakagawa在2012年的研究表明,另一个lncRNA-Malat1可能调节NEAT1的表达,当在特定细胞系中敲除Malat1 后,NEAT1表达明显下调,表明Malat1 可以调控NEAT1 表达,影响旁斑形成[20]。本试验结果显示NEAT1可能具有维持小鼠精子发生的作用,从而一定程度参与小鼠的生殖过程,NEAT1 在该过程发挥功能的具体机制是什么以及Malat1 是否参与了调控NEAT1的表达进而影响后续的旁斑形成和功能发挥则需要进一步实验进行验证。
4 结论
NEAT1在小鼠睾丸组织、GC-1细胞系中均表达;构建的NEAT1干扰病毒可使NEAT1表达水平下调69%,表明该慢病毒效果良好;干扰NEAT1后小鼠睾丸里的精子发生受到一定影响,表明NEAT1参与维持雄性小鼠的精子发生。
[1]Pandey R R,Mondal T,Mohammad F,et al.Evolution and functions of long noncoding RNAs[J].Cell,2009,136:629-641.
[2]Mitchell Guttman,Julie Donaghey,Bryce W.Carey,et al.lincRNAs act in the circuitry controlling pluripotency and differentiation[J].Nature,2011,10:1 038-1 048.
[3]Christine M Clemson,John N Hutchinson,Sergio A Sara,et al.An Architectural Role for a Nuclear Noncoding RNA:NEAT1RNA Is Essential for the Structure of Paraspeckles[J].Molecular Cell,2009,33:717-726.
[4]Bernard D,Prasanth K V,Tripathi V,et al.A long nuclearretained non-coding RNA regulates synaptogenesis by modulating gene expression[J].The EMBO Journal,2010,29:3 082-3 093.
[5]Bond C S,Fox A H.Paraspeckles:nuclear bodies built on long noncoding RNA[J].The Journal of Cell Biology,2009,186:637-644.
[6]Ling-Ling Chen,Gordon G Carmichael.Altered Nuclear Retention of mRNAs Containing Inverted Repeats in Human Embryonic Stem Cells:Functional Role of a Nuclear Noncoding RNA[J].Molecular Cell,2009,35:467-478.
[7]Popovici C,Fallet M,Marguet D,et al.Intracellular trafficking of LET-756,a fibroblast growth factor of C.elegans,is controlled by a balance of export and nuclear signals[J].Exp Cell Res,2006,312:1 484-1 495.
[8]Ulfig N,Briese V.Immunolabelling of spliceosomes in sections and cultured astrocytes of human fetal brain tissue[J].Brain Dev,1999,21:51-58.
[9]Sunwoo H,Dinger M E,Wilusz J E,et al.MEN epsilon/beta nuclear-retained non-coding RNAs are up-regulated upon muscle differentiation and are essential components of paraspeckles[J].Genome Res,2009,19:347-359.
[10]Clemson C M,Hutchinson J N,Sara S A,et al.An architectural role for a nuclear noncoding RNA:NEAT1RNA is essential for the structure of paraspeckles[J].Mol Cell,2009,33:717-726.
[11]Sasaki Y T,Ideue T,Sano M,et al.MENepsilon/beta noncod
ing RNAs are essential for structural integrity of nuclear pa
raspeckles[J].Proc Natl Acad Sci USA,2009,106:2 525-2 530.
[12]Kanatsu-Shinohara M,Ogonuki N,Inoue K,et al.Longterm proliferation in culture and germLine transmission of mouse male germLine stem cells[J].Biology of Reproduction,2003,69:612-616.
[13]Archa H Fox,Angus I Lamond.Paraspeckles[J].Cold Spring Harb Perspect Biol,2010,2:687.
[14]李新华,安 鹏,李俊琴,等.银屑病患者外周血T 细胞SCL及NEATl的表达[J].中国药物与临床,2013(13):280-282.
[15]Yun-Sook Kim,Jin Hwan Do,Sumi Bae,et al.Identification of differentially expressed genes using an annealing control primer system instage III serous ovarian carcinoma[J].BMC Cancer,2010,10:576.
[16]Nakagawa S,Naganuma T,Shioi G,et al.Paraspeckles are subpopulation specific nuclear bodies that are not essential in mice[J].J Cell Biol,2011,193(1):31-39.
[17]Buaas F W,Kirsh A L,Sharma M,et al.Plzf is required in adult male germ cells for stem cell self-renewal[J].Nature Genetics,2004,36:647-652.
[18]Oatley J M,Avarbock M R,Telaranta A I,et al.Identifying genes important for spermatogonial stem cell self-renewal and survival[J].Proceedings of the National Academy of Sciences,2006,103:9 524-9 529.
[19]Chen C,Ouyang W,Grigura V,et al.ERM is required for transcriptional control of the spermatogonial stem cell niche[J].Nature,2005,436:1 030-1 034.
[20]Nakagawa S,LP J Y,Shioi G,et al.Malat1is not an essential component of nuclear speckles in mice[J].RNA,2012,18(8):1 487-1499.
猜你喜欢
中国临床医学影像杂志(2022年5期)2022-07-26 07:11:58
右江医学(2021年9期)2021-10-22 05:15:28
昆明医科大学学报(2021年1期)2021-02-07 01:06:12
中国生殖健康(2019年7期)2019-01-06 09:27:30
癌变·畸变·突变(2016年5期)2016-08-22 05:55:20
山东医药(2015年14期)2016-01-12 00:39:43
江苏大学学报(医学版)(2015年2期)2015-04-17 06:49:51
中国医药导报(2015年26期)2015-02-28 22:07:44
安徽医药(2014年9期)2014-03-20 13:14:09
遗传(2014年3期)2014-02-28 20:59:01
