
α-松油醇对意大利青霉的抑制作用
2014-01-18 08:33欧阳秋丽陶能国何湘丽
食品科学 2014年11期
欧阳秋丽,贾 雷,陶能国*,何湘丽
(湘潭大学化工学院,湖南 湘潭 411105)
α-松油醇对意大利青霉的抑制作用
欧阳秋丽,贾 雷,陶能国*,何湘丽
(湘潭大学化工学院,湖南 湘潭 411105)
测定α-松油醇对意大利青霉菌丝体形态及菌丝体生长的影响,并对其抑菌机制进行初步探讨。结果表明:α-松油醇能明显抑制意大利青霉菌丝体生长,最小抑菌浓度(minimal inhibitory concentration,MIC)和最小杀菌浓度(minimal fungicidal concentration,MFC)分别为2.00 μL/mL和8.00 μL/mL;经MIC和MFC α-松油醇处理后,意大利青霉菌丝体胞外pH值、胞外电导率及260 nm条件下吸光度显著增加,总脂质含量有显著下降。提示α-松油醇能改变细胞膜的通透性、破坏细胞膜结构的完整性,导致胞内成分泄露,从而抑制意大利青霉菌丝体生长。
α-松油醇;意大利 青霉;细胞膜;抑制作用
意大利青霉(Penicillium italicum)引起的青霉病是柑橘主要贮藏期病害之一,每年造成大量的经济损失[1]。由于化学杀菌剂易带来食品安全,以及病原菌对化学杀菌剂抗性不断增强等问题,植物精油等生物防治法逐渐成为采后柑橘病害的替代方案[2]。Caccioni等[3]发现向液体培养基中添加250~5 000 μL/L甜橙、酸橙、宽皮柑橘、葡萄柚和枳橙精油能不同程度地抑制意大利青霉菌丝体生长,其中枳橙精油的抑菌效果最好。向固体培养基中添加3 000 μL/L柠檬醛可完全抑制意大利青霉孢子的萌发和生长[4]。Tripathi等[5]研究表明,100 μg/L薄荷、500 μg/L灰罗勒和200 μg/L生姜精油可使接种意大利青霉的甜橙贮藏时间分别延长6、6、4 d;延长酸橙的贮藏时间8、6、8 d。40 μL/平板的宽皮柑橘和甜橙精油、0.5 μL/平板柠檬醛和5 μL/平板芳樟醇能完全抑制意大利青霉孢子萌发和芽管生长[6]。Yahyazadeh等[7]研究指出,熏蒸400 μL/L麝香草和丁香精油可有效杀死汤姆森脐橙和伏令夏橙果实表面接种的意大利青霉。含质量分数0.5%~3%佛手柑精油和壳聚糖的复合物能不同程度抑制意大利青霉生长[8]。前期研究表明,5.0 μL/mL不同月份(7—10月份)的椪柑果皮精油能完全抑制意大利青霉生长,且椪柑果皮精油中的单一组分,如0.5 μL/mL柠檬醛、辛醛、1.0 μL/mL壬醛、癸醛以及2.0 μL/mLβ-芳樟醇、α-松油醇均能完全抑制意大利青霉菌丝体生长[9]。
α-松油醇又名α-萜品醇,是一种具有丁香味的萜类化合物,广泛存在于植物精油中[2-9]。研究表明,α-松油醇对多种革兰氏菌和多种真菌均有较强的抑制作用,是一种潜在的抑菌物质[9-11]。本实验拟探讨α-松油醇抑制意大利青霉菌丝体生长的作用机制,为柑橘采后病害生物防治提供理论依据。
1 材料与方法
1.1 材料与试剂
α-松油醇(纯度90%) 西格玛奥德里奇(上海)贸易有限公司;香草醛(分析纯) 梯希爱(上海)化成工业发展有限公司。
意大利青霉(P. italicum)分离自腐烂柑橘果实,现保存于湘潭大学生物与食品工程专业实验室。
1.2 仪器与设备
SPX-250B-D生化培养箱 上海博迅实业有限公司;Ecl ipse600光学显微镜 尼康仪器(上海)有限公司;UV2450紫外分光光度计、BL320H电子天平 日本Shimadzu公司;TDL5A 大容量冷冻离心机 长沙英泰仪器有限公司;FA1004N分析天平 上海精密科学仪器厂;PHS-W系列微机型pH/mV计、DDS-W系列微机型电导率仪 上海般特仪器有限公司;CoolSafe 110-4L真空冷冻干燥机 丹麦Labo Gene公司。
1.3.1 α-松油醇对意大利青霉菌丝体的作用
采用琼脂稀释培养法[12]研究α-松油醇对意大利青霉菌丝体生长的影响。α-松油醇的添加量设定采用倍半稀释法,实验终量为0.00、0.25、0.50、1.00、2.00、4.00 μL/mL和8.00 μL/mL。每个培养皿接入3个生长一致的菌饼(d=6.0 mm),置于(25±2)℃恒温培养箱培养4 d。以生长2 d时完全没有菌丝生长的最低添加量作为α-松油醇的最小抑菌浓度(MIC),以生长4 d时杀死99.9%的菌丝生长的最低添加量作为最小杀菌浓度(MFC)[14],采用十字交叉法每天测量菌丝体生长直径,用相应体积的0.5%吐温-80为对照(CK),结果取平均值。挑取CK、MIC、MFC处理4 d后 的菌丝体,光学显微镜下(4(目镜)×40(物镜)倍)观察形态变化并拍照。
1.3.2 α-松油醇对意大利青霉胞外胞外pH值的影响
采用微型pH/mV计测定α-松油醇对意大利青霉胞外pH值的影响。将意大利青霉孢子洗脱于马铃薯葡萄糖液体培养基(potato dextrose broth,PDB),(25±2)℃,160 r/min振荡培养48h后,4 000 r/min离心15 min,水洗3 次,收集菌丝体,重新悬浮于磷酸缓冲液(pH 7.0)中。加入α-松油醇添加量为0、MIC和MFC时处理0、30、60、120 min后,测定胞外pH值,用无菌水作对照。
1.3.3 α-松油醇对意大利青霉胞外电导率的影响
采用微型电导仪测定α-松油醇 对意大利青霉胞外电导率的影响。培养及处理方法如1.3.2节,测定其胞外电导率,用无菌水作对照。
1.3.4 α-松油醇对意大利青霉菌丝体细胞成分释放的影响
20世纪80年代,北京与台北两地的流行音乐个人演唱会市场都处于起步及初始发展阶段,因此个人演唱会的数量有限(表2、表4),但仍有香港地区、韩国、日本以及欧美国家的歌手(表3、表5),前往两地举办个人演唱会。
参照Paul等[13]的方法,培养及处理方法如1.3.2节,加入α-松油醇添加量为0、MIC、MFC,处理0、30、60、120 min后取样, 将样品置于12 000r/min离心2min收集上清液,用紫外分光光度计在波长260 nm处测吸光度,对照组用PBS(pH 7.0)进行校正。
1.3.5 α-松油醇对意大利青霉总脂质含量的影响
采用香草醛硫酸比色法[14]测定处理后的菌丝体。在加入α-松油醇添加量为0、MIC、MFC处理120 min后,用真空冷冻干燥机干燥2 h,称取一定量干燥菌丝体,利用液氮充分研磨,按照体积比1∶1∶1加入甲醇、氯仿、水剧烈振荡充分萃取脂质,将含有脂质的混合液与浓硫酸沸水浴加热10 min,室温下冷却后加入磷光体香草醛素充分混匀,在520 nm波长处测其吸光度,最后利用胆固醇标准曲 线计算其脂质含量,脂质含量以mg/g表示。
1.4 数 据分析
每组有3 次重复,采用常规的分析方法标注标准偏差,数据分析采用SPSS16.0统计分析软件,ANOVA来分析显著性差异(P<0.05)。
2 结果与分析
2.1 α-松油醇对意大利青霉菌丝体的作用
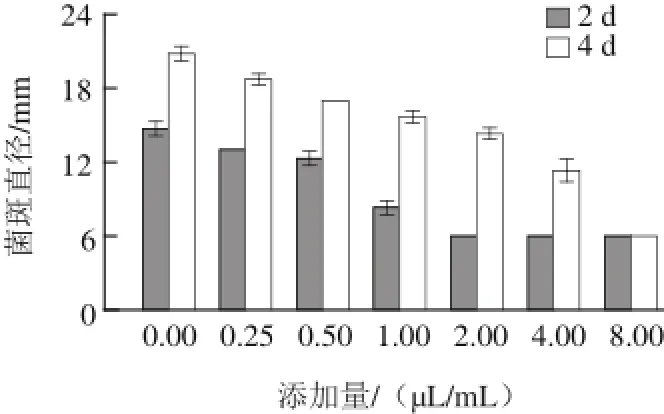
图1 1 α-松油醇对意大利青霉菌丝体生长的影响Fig. 1 Effect of α- terpineol on mycelial growth of P. italicum
由图1可知,随着添加量增加,α-松油醇对意大利青霉的抑制作用逐渐增强。培养2 d后,当α-松油醇的添加量≥2.00 μL/mL时,意大利青霉基本无生长,说明α-松油醇对意大利青霉的最小抑菌浓度MIC为2.00 μL/mL。培养4 d后,8.00 μL/mL α-松油醇能完全抑制意大利青霉菌丝体生长,0.25~4.00 μL/mL α-松油醇也能不同程度抑制意大利青霉菌丝体生长(P<0.05),因而α-松油醇对意大利青霉的最小杀菌浓度MFC为8.00 μL/mL。
由图2可知,α-松油醇对意大利青霉菌丝体形态存在明显影响,未处理的菌丝体表面形状规则,粗细均匀,结构规则,处于正常生长状态(图2a);经MIC浓度处理的菌丝体虽较为饱满,但细胞中 的内容物减少(图2b);经MFC处理后,菌丝体出现了干瘪、细小等形态,细胞内容 物泄漏更为严重(图2c)。

图2 2 α-松油醇对意大利青霉菌丝体形态的影响Fig.2 Effect of α-terpineol on morphology of P. italicum
2.2 α-松油醇对意大利青霉胞外pH值的影响
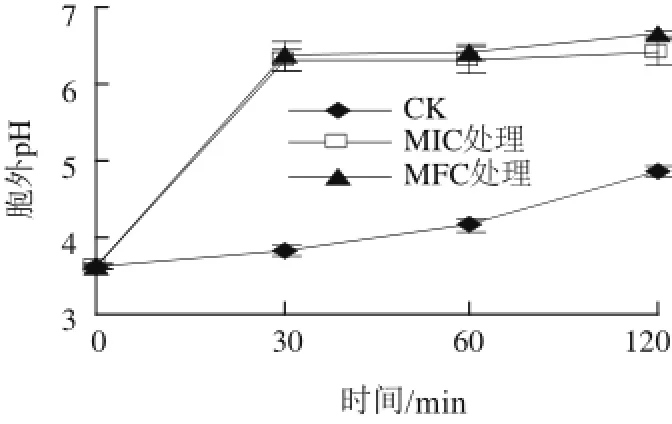
图 33 α-松油醇对意大利青霉胞外pH值的影响Fig.3 Effect of α-terpineol on extracellular pH of P. italicum
由图3可知,处理30 min时,MIC和MFC处理组的胞外pH值显著上升,但两者之间无显著差异,分别为6.30±0.14和6.37±0.18,显著高于对照的3.82±0.08(P<0.05)。随着处理时间延长到120 min,对照组CK胞外pH值显著升高,但低于处理组;而MIC与MFC组胞外pH值趋于平缓,无显著性差异。
2.3 α-松油醇对意大利青霉胞外电导率的影响
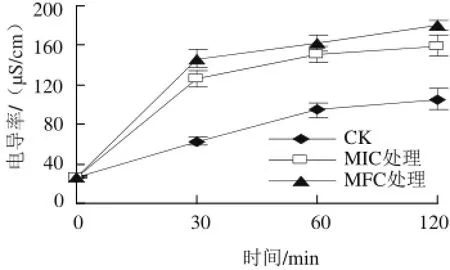
图4 4 α-松油醇对意大利青霉胞外电导率的影响Fig.4 Effect of α-terpineol on extracellular conductivity of P. italicum
由图 4可知,经α-松油醇处理后,意大利青霉的胞外电导率随处理时间和添加量的增加不断上升,以0~30 min内变化最为明显。处理30 min时, MIC和MFC处理组的电导率分别为(126.5±8.1)和(146.6±9.7)μS/cm,显著高于CK组的(63.0±4.4)μS/cm(P<0.05),之后仍有不同幅度增加。处理120 min时,CK、MIC和MFC处理组电导率比初始电导率分别增加了78.9、132.9、153.2 μS/cm。
2.4 α-松油醇对意大利青霉菌丝体细胞成分释放的影响

图5 5 α-松油醇对意大利青霉260 nm细胞成分释放的影响Fig.5 Effect of α-terpineol on absorbance at 260 nm of P. italicum
由图5可知,α-松油醇显著增加意大利青霉260 nm细胞成分释放,以处理前30 min内变化最为明显。处理30 min时,CK组的OD260nm值为(0.024±0.017),显著低于MIC处理组(0.129±0.002)以及MFC处理组(0.161±0.013)(P<0.05)。随着时间延长,CK组OD260nm值基本保持不变。对MIC和MFC处理组而言,30~60 min内OD260nm值仍有显著增加,但60 min后OD260nm值基本保持不变。
2.5 α-松油醇对意大利青霉总脂质含量的影响

图6 6 α-松油醇对意大利青霉总脂质含量的影响Fig. 6 Effect of α-terpineol on total lipids of P. italicum
由图6可知,处理120 min后,意大利青霉的总脂质含量明显下降,且处理与未处理的意大利青霉总脂质含量有显著性差异(P<0.05),CK、MIC、MFC的脂质含量分别为(238.8±45.6)、(138.0±16.8)和(112.4±14.5)mg/g,但MIC和MFC处理组之间差异不显著。
3 讨 论
本实验探讨了α-松油醇对意大利青霉菌丝体生长的影响,结果表明,α-松油醇也能不同程度抑制意大利青霉菌丝体生长,最小抑菌浓度(MIC)和最小杀菌浓度(MFC)分别为2.00 μL/mL和8.00 μL/mL,这一结果与前期研究结果一致,但优于不同发育时期的椪柑精油[9]。
关于萜类化合物的抑菌机制,目前了解的不是很透彻。通常认为,萜类化合物能破坏并穿透细菌及真菌细胞的脂质结构,增加膜流动性,破坏细胞膜的完整性,提高膜通透性,细胞内容物发生泄漏,最终抑制细菌及真菌生长[15-22]。本实验中也存在类似现象,当添加量为MIC和MFC的α-松油醇加入后,意大利青霉菌丝体胞外pH值、胞外电导率以及OD260nm都有显著上升,且在0~30 min时变化最快。胞外pH值的增加意味着胞内pH值的下降,细胞因H+积累而发生酸化,易造成细胞内生理生化过程发生不可逆的损伤[18]。电导率的增加与胞内K+、Mg2+、Ca2+等外泄有关[18-20],30 min时MIC、MFC组的电导率分别达到了(150.3±8.1)和(161.7±10.3)μS/cm,显著高于CK组(P<0.05),直至处理120 min,各处理组电导率仍存在显著性差异,说明细胞膜通透性发生了变化。260 nm是 核酸的特征波长,在处理过程中,0~120 min间各组OD260nm值差异显著,且在30 min时处理组就已经明显高于CK组,表明意大利青霉胞内核酸等物质泄漏量增多,暗示着细胞内的遗传过程受到影响[13,21-22]。脂质占细胞干质量的6 0%~80%,是细胞膜的结构物质[14],在120 min时MIC、MFC处理组脂质含量分别下降100.8、126.4 mg/g(P<0.05),说明细胞膜的组成发生变化,对细胞膜的通透性及细胞活性产生了潜在的伤害。
4 结 论
α-松油醇是一种极具潜力的抑菌物质,可有效破坏意大利青霉菌丝体细胞膜结构的完整性,增加细胞膜通透性,致使胞内物质如金属离子、核酸等外泄以及胞内H+积累,从而导致意大利青霉的菌丝体死亡。
[1] 龙超安, 邓伯勋, 何秀娟. 柑橘青、绿霉病高效拮抗菌34-9的筛选及其特性研究[J]. 中国农业科学, 2005, 38(12): 2434-2439.
[2] SHARMAN N, TRIPATHI A. Fungitoxicity of the essential oil of Citrus sinensis on post-harvest pathogens[J]. World Journal of Microbiology and Biotechnology, 2006, 22(6): 587-593.
[3] CACCIONI D R L, GUIZZARDI M, BIONDI D M, et al. Relationship between volatile components of citrus fruit essential oils and antimicrobial action on Penicillium digitatum and Penicillium italicum[J]. International Journal of Food Microbiology, 1998, 43(1/2): 73-79.
[4] KLIEBER A, SCOTT E, WURYATMO E. Effect of method of application on antifungal eff cacy of citral against postharvest spoilage fungi of citrus in culture[J]. Australasian Plant Pathology, 2002, 31(4): 329-332.
[5] TRIPATHI P, DUBEY N K, BANERJI R, et al. Evaluation of some essential oils as botanical fungitoxicants in management of postharvest rotting of citrus fruits[J]. World Journal of Microbiology and Biotechnology, 2004, 20(3): 317-321.
[6] DROBY S, EICK A, MACARISIN D, et al. Role of citrus volatiles in host recognition, germination and growth of Penicillium digitatum and Penicillium italicum[J]. Postharvest Biology and Technology, 2008, 49(3): 386-396.
[7] YAHYAZADEH R, OMIDBAIGI Z R, FAGHIH-NASIRI M, et al. Control of Penicilliu m decay on citrus fruit using essential oil vapours of thyme or clove inside polyethylene and nano-clay polyethylene films[J]. The Journal of Horticultural Science and Biotechnology, 2009, 84(4): 403-409.
[8] SÁNCHEZ-GONZÁLEZ l, CHÁFER M, CHIRALT A, et al. Physical properties of edible chitosan f lms containing bergamot essential oil and their inhibitory action on Penicillium italicum[J]. Carbohydrate Polymers, 2010, 82(2): 277-283
[9] 贾雷, 何湘丽, 陶能国, 等. 不同发育期槛柑精油对意大利青霉和指状青霉的抑制作用[J]. 食品工业科技, 2013, 34(7): 68-72.
[10] PARK M J, GWAK K S, YANG I, et al. Effect of citral, eugenol, nerolidol and α-terpineol on the ultrastructural changes of Trichophyton mentagrophytes[J]. Fitoterapia, 2009, 80(5): 290-296.
[11] SCORA K M, SCORA R W. Effect of volatiles on mycelium growth of Penicillium digitatum, P. italicum, and P. ulaiense[J]. Journal of Basic Microbiology, 1998, 38 (5/6): 405-413.
[12] SHARMA N, TRIPATHI A. Effects of Citrus sinensis (L.) Osbeck epicarp essential oil on growth and morphogenesis of Aspergillus niger (L.) Van Tieghem[J]. Microbiological Research, 2008, 163(3): 337-344.
[13] PAUL S, DUBEY R C, MAESWARI D K, et al. Trachyspermum ammi (L.) fruit essential oil influencing on membrane permeability and surface characteristics in inhibiting food-borne pathogens[J]. Food Control, 2011, 22(5):725-731.
[14] HETAL G A, SARAN M M, ABU S A N K, et al. Effects of Cymbopogon citratus L.essential oil on the growth ,morphogenesis and af atoxin production of Aspergillus f avus ML2-strain[J]. Journal of Basic Microbiology, 2007, 47(1): 5-15.
[15] HADA T, INOUE Y, SHIRAISI A, et al. Leakage of K+ions from Staphylococcus aureus in response to tea tree oil[J]. Journal of Microbiological Methods, 2003, 53 (3): 309-312.
[16] SHAO X F, CHENG S, WANG H F, et al. The possible mechanism of antifungal action of tea tre oil on Botrytis cinerea[J]. Journal of Applied Microbiology, 2013, 114(6): 1642-1649.
[17] BENNIS S, CHAMI F, Chami N, et al. Surface alteration of Saccharomyces cerevisiae induced by thymol and eugenol[J]. Letters in Applied Microbiology, 2004, 38(6): 454-458.
[18] RAYBAUDI-MASSILIA R M, MOSQUEDA-MELGAR J, MARTINBELLOSO O. Antimicrobial activity of essential oils on Salmonella enteritidis, Escherichia coli, and Listeria innocua in fruit juices[J]. Journal of Food Protection, 2006, 69(7): 1579-1586.
[19] BARD M, ALBRECHT M R, GUPTA N, et al. Geraniol interferes with membrane functions in strains of Candida and Saccharomyces[J]. Lipids, 1988, 23(6): 534-538.
[20] BURT S. Essential oils: their antibacterial properties and potential applicati ons in foods: a review[J]. International Journal of Food Microbiology, 2004, 94(3): 223-253.
[21] DEVI K P, NISHA S A, SAKTHIVEL R, et al. Eugenol (an essential oil of clove) acts as an antibacterial agent against Salmonella typhi by disrupting the cellular membrane[J]. Journal of Ethnopharmacology, 2010, 130(1): 107-115.
[22] de SOUZA E L, de BARROS J C, de OLIVEIRA C E V, et al. Influence of Origanum vulgare L. essential oil on enterotoxin production, membrane permeability and surface characteris tics of Staphylococcus aureus[J]. International Journal of Food Microbiology, 2010, 137(2): 308-311.
Inhibitory Effect of α-Terpineol on Penicillium italicum
OUYANG Qiu-li, JIA Lei, TAO Neng-guo*, HE Xiang-li
(College of Chemical Engineering, Xiangtan University, Xiangtan 411105, China)
The effect of α-terpineol at various concentrations on the morphology and mycelial growth of Penicillium italicum was evaluate d, and the corresponding mechanism was explored. Res ults showed that α-terpineol had a significant inhibitory effect on the mycelial growth of P. italicum, with minimal inhibitory concentr ation (MIC) and minimal fungicidal concentration (MFC) of 2.00 and 8.00 øL/mL, respectively. After being treated with α-t erpineol a t MIC or MFC, marked increases in the extracellular pH, extracellular conductivity and absorbance at 260 nm were observed. Moreover, a decrease in total lip id content was also observed. Our research indicated that α-terpineol can increase the membrane permeability and destroy the integr ity of the membrane of P. italicum, thus resulting in the leakage of cellular materials and inhibiting the mycelial growth of P. italicum.
α-terpineol; Penicillium italicum; cell membrane; inhibitory effect
S436.66
A
1002-6630(2014)11-0032-04
10.7506/spkx1002-6630-201411007
2013-07-13
国家自然科学基金面上项目(31271964);湖南省教育厅青年项目(12B126);国家级大学生创新训练计划项目(201210530012)
欧阳秋丽(1990—),女,硕士研究生,研究方向为柑橘贮藏与保鲜。E-mail:ouyang199006@126.com
*通信作者:陶能国(1979—),男,教授,博士,研究方向为柑橘贮藏与保鲜。E-mail:nengguotao@126.com
猜你喜欢
医疗装备(2023年3期)2023-03-22
林产工业(2021年7期)2021-07-28
昆明医科大学学报(2021年4期)2021-07-23
小学生导刊(2018年18期)2018-05-23
环境保护与循环经济(2017年3期)2017-09-26
中国酿造(2016年12期)2016-03-01
安全(2015年7期)2016-01-19
大连工业大学学报(2015年4期)2015-12-11
信阳师范学院学报(自然科学版)(2015年4期)2015-08-09
武夷学院学报(2015年3期)2015-07-18
