
红外相机技术在鼠类密度估算中的应用
2013-12-16 08:19章书声鲍毅新王艳妮方平福
生态学报 2013年10期
章书声,鲍毅新,王艳妮,方平福,叶 彬
(浙江师范大学生态研究所,金华 321004)
野外调查小型兽类密度时,根据不同种类的生活习性而采用不同的方法,对旱獭、黄鼠等昼行性种类常采用堵洞盗开法、弓形夹法、挖洞法、计洞法等;对跳鼠等夜行性种类常采用夹夜法;鼢鼠等地下生活的种类一般采用土丘系数法和切洞堵洞法;田鼠和绒鼠等地面生活种类可采用夹日法[1]。
传统的小型兽类密度调查方法对动物的生境会产生一定的干扰。红外线触发自动数码相机陷阱技术(ITCT),简称红外相机技术,已成为现代生物资源调查中的重要方法之一[2]。采用该方法研究兽类种群密度时,对于一些大型兽类,如孟加拉虎(Panthera tigris tigris)[3-4]、雪豹(Uncia uncia)[5-6]可以根据其斑纹、体型、毛色等“自然标记”进行个体识别,并估算其种群密度。但对于夜行性小型兽类,因“自然标记”不明显,难以通过夜间所摄相片进行个体识别。Rowcliffe等[7]在采用红外相机技术估算Whipsnade野生动物园内中国水鹿(Cervus unicolor)、小麂(Muntiacus reevesi)等动物的种群密度时,将气体分子碰撞率原理应用于相机与动物之间接触率的计算,该方法无需对动物进行个体识别。本文尝试采用红外相机技术,对地面上活动的啮齿目动物(鼠类)在不对物种进行个体识别的情况下,估算古田山自然保护区样地中的鼠类密度,并将估算结果与标志重捕法获得的结果进行对比,来验证此模型准确度和可靠性。
1 研究方法
1.1 研究地区
古田山位于浙江省开化县境内西北部,与江西省婺源县、德兴市接壤,地理范围为29°10′19.4″—29°17′41.4″N,118°03′49.7″—118°11′12.2″E,属典型中亚热带季风气候区,四季分明。区内保存着完好的以甜槠(Castanopsis eyrei)和木荷(Schima superba)等为主要优势种的常绿阔叶林,保护区总面积81 km2,其中核心区面积为15.5 km2,为森林的长期生物多样性动态监测和机理研究提供了很好的研究基地[8]。本项研究所选用的监测样地是参照(CTFS)标准所建设的1个24 hm2常绿阔叶林(东西长600 m、南北宽400 m)常绿阔叶林样地[9]。
1.2 红外相机技术
2009年5月至2011年7月,将24 hm2的研究样地分成18个栅格利用分层随机抽样法原理均匀布置视保嘉SG550红外相机20台(图1,比例尺为1∶20)以监测样地的鼠类密度。相机分布密度为0.83台/hm2,与其他类似研究比较,本研究相机分布密度较高(表1)。
1.2.1 气体分子碰撞率原理
根据物理学的经典理想气体微观模型,每个分子被模型化为具有一定质量、不计体积、无气体间粘滞力的小球。所以,理想气体就是数量巨大的理想弹性小球的无规则运动[13]。该模型的使用条件包括:(1)气体分子线度远小于与分子间的距离,可忽略不计(鼠类体型远小于鼠类与相机的距离);(2)遵循牛顿运动定律,每个分子的碰撞为完全弹性碰撞(鼠类被相机拍摄本身不存在能量的损耗);(3)分子间的相互作用力、重力可忽略不计(模型所依据的是鼠类与相机的碰撞率,并非鼠类之间的相遇率,满足此要求)(4)模型的统计假设为:对大量气体分子来说,气体分子处于热动态平衡时,分子可沿各个方向无规则运动(鼠类在样地内可以沿任何方向无规则活动)

图1 古田山24 hm2样地红外相机分布图(黑点即相机位置)Fig.1 The camera distribution map in the 24 hm2plot of Gutianshan National Natural Reserve(black spots present camera sites)

表1 本研究及相关文献中相机分布密度Table 1 The distributed density of camera in this study and the literatures
1.2.2 估算方法
本次调查对象为地面上活动的啮齿目动物(鼠类),活动区域属于二维空间。根据上述原理,分子在二维封闭空间中移动,在一定时间内移动覆盖的面积是由分子线度(鼠类体型,在原理中已被忽视)和移动距离组成的,而在此时间范围内分子之间的期望碰撞率是指全部分子移动覆盖的面积占总面积的比例值[14]。应用到生物学当中就是一个相对静止分子(鼠类)与一个圆形的检测区域内(相机的视角)碰撞数y(鼠类个体独立照片总数),由分子(鼠类)的移动速度v,移动时间t(相机监测日),分子移动的距离r(相机检测区域的半径),分子数密度D(鼠类密度)组成,如公式(1)。

然而,相机检测区域并不是圆形的,而是以半径为r和角度θ组成的扇形面积,故动物所覆盖路径的宽度已经不再是单纯2 r,而是应该是所有可能的角度组成的扇形面积。其中,(π-θ)/2≤γ≤π/2,由Rowcliffe等[7]推导出:

对相近地点或时间段连续拍摄的物种照片采用相同侧面、部位的斑纹、体型、毛色等特征进行比对,来判断是否属同一次数,定义为独立照片数(IP)。其次是取样时间,Rowcliffe等[7]研究中国水鹿的种群密度时统计t值是以累积6周相机白天(12h)工作作为有效时数的,而鼠类属于夜行动物且需长年观察,故本文以月为单位计算其相机工作日。以下列公式计算:

式中,MCV为月拍摄值,y为当月鼠类个体独立照片总数,t为相机布放当月的天数。

所用红外相机的监测角度θ为0.872 rad(50°),相机的监测半径r为0.01 km(10 m)。研究发现样地内鼠类一般日活动距离为100—300 m(根据样地内号桩标号计算),布笼时间为24 h,设置鼠类日移动速度v1(0.1 km/d),v2(0.2 km/d),v3(0.3 km/d)3 个参数。
1.3 标志重捕法
2009—2011年每年的5月和10月(其中2009年为7月),分别在同一样地内采用传统的网格布笼法[15]对样地内的鼠类进行标志重捕,具体布笼地点如图1当中的小实线方框,以10 m×10 m方式布笼,共置布笼点160个。在每个布笼点放置1个鼠笼,以红枣为诱饵进行重捕。傍晚开笼,次日上午检查后关笼。对所捕鼠类进行物种鉴定,首次捕获的个体以切趾法进行个体标记,并记录捕获日期、鼠号和笼号等,每次连续重捕7d。采用Jolly-sober法和最小存活数量法来估算样地内的鼠类密度[16]。
2 结果
2.1 红外相机技术估算鼠类密度
由3个鼠类移动速度(0.1、0.2、0.3 km/d)参数得出的鼠类密度分别为D1,D2,D3(表2),相互之间存在极显著差异(F=17.326,df=2,P <0.01),用LSD法两两比较,D1与D2之间不存在显著差异(P >0.05),D3与D1、D3与D2之间却都存在极显著差异(P <0.01)。

表2 运用红外相机技术估算鼠类密度Table 2 Rodent population density estimated by infrared triggered camera technique

续表
2.2 标志重捕法估算鼠类密度
自2009年到2011年,对24 hm2大样地内利用标志重捕法捕鼠调查结果表明,样地内有社鼠(Niviventer confucianus)、刺毛鼠(N.fulvescens)和白腹巨鼠(Leopoldamys edwardsi),其中社鼠占90%以上。由于获取鼠类的样本数量较少,重捕率小于50%,不满足J-S模型的前提条件,故采用最小存活数量法(枚举法)估算样地内的鼠类密度,结果见表3。

表3 用标志重捕法估计的鼠类密度Table 3 Rodent population density estimated by capture mark-recapture method
2.3 两种方法比较
由红外相机技术估计得出的鼠类密度D1、D2与D3和标志重捕法在相对应的5个月份(2011年10月无相机拍摄除外)所得鼠类密度D进行对比,D与D1存在极显著差异(t=3.910,df=8,P <0.01),D与D2(t=1.931,df=8,P=0.090)以及与D3(t=0.430,df=8,P=0.679)之间皆无显著差异,其中D 与 D3之间的差异更小,契合程度更好。根据每月所拍鼠类独立照片(IP)获得月拍摄值(MCV),以D3为准,得出2009年5月至2011年7月每月样地内的鼠类密度(图2),平均密度为(6.57±0.81)只/hm2,鼠类密度符合正态分布(P >0.05)。
3 讨论
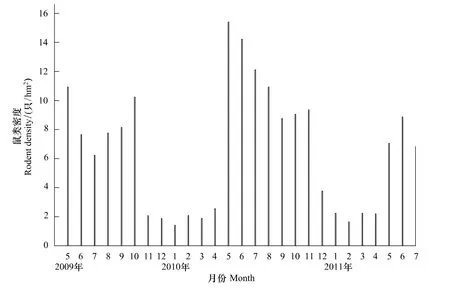
图2 红外相机技术估计的鼠类密度季节消长(以D3为标准)Fig.2 The seasonal changes in the rodent density(inds/hm2)which obtained from infrared triggered cameras technology
鼠类的日活动距离随种类、性别、年龄、体重与巢区面积等而异。在与古田山同纬度的千岛湖对社鼠的调查结果显示,雄鼠平均活动距离为(111.47±6.80)m(22.35—275 m),雌鼠为(84.82±4.69)m(14.14—290.49 m)[15]。根据本文研究样地中社鼠占绝对优势的情况,我们设置了鼠类日移动速度v1、v2、v33个参数。当鼠类日活动距离为0.1 km(v1)时,得出的密度D1过大,与标志重捕法获得的鼠类密度D存在着显著差异。D2与D3则均与D值不存在显著差异,说明红外相机技术是可以估计样地内鼠类密度的,且假设鼠类日移动距离为0.2—0.3 km是合理的。但D与D2的差异(P=0.090)比与D3(P=0.679)的差异更大,说明D3与传统的标志重捕法获得的鼠类密度契合程度较高,故可以认为样地内鼠类日移动平均距离准确值为0.3 km。
相机分布的密度往往与动物的家域或种群密度成反比例关系(表1),如大型猫科动物华南虎、雪豹家域面积约达到100 km2,相机分布的密度约为0.1台/km2[5-6,10-11];对家域面积相对较小的小麂台湾亚种将相机分布密度增高至0.3台/km2[12];而本模型样地内鼠科动物的家域范围更小,故将相机密度增加至0.83台/hm2,虽不及样地内布笼密度1.29只/hm2,但鼠类在样地内一般日平均活动距离不超过300 m,近乎1台/hm2已经属于高密度的布置,具有较高的准确性和精确度。
相机监测时间也可影响密度估算的精确度。在常用的标志重捕法中,要提高重捕率则须增加野外调查取样的强度和难度,在捕捉努力保持不变的情况下单方面提高重捕率必然导致取样面积的减少,这样就有可能陷入调查动物样本量不足的困境,而单纯地延长重捕时间也可能增加对种群的干扰而影响标记个体的重捕率[17]。红外相机捕捉技术在调查样本时间方面具有更大的优越性,如本文求得2009年5月至2011年7月鼠类月平均密度(图2),即假设忽略动物的迁入、迁出、出生、死亡等参数,将1个月内鼠类种群默认为是一个闭合的种群。通过监测此闭合的种群,发现2009、2010和2011年都反映出5月份伊始持续到夏季末属于鼠类密度的高峰期。其结果与杨春文[18]对东北主要林区森林5种啮齿动物共存机制的研究结论相似,夏季食物资源丰富,种间竞争较小;而11月到初春时分由于冬季的严寒、捕食压力(红外拍摄同时发现黄腹鼬Mudtela kathiah、鼬獾Melogale moschata和豹猫Prionailurus bengalensis)及食物匮乏导致鼠类数量降低,通过减少种内及种间食物竞争压力,使群落结构保持相对稳定。
因此,采用红外相机技术无需个体识别即可估计出鼠类密度,对比传统鼠类密度调查法,该技术具有可昼夜连续工作,劳动力成本低、无创伤、环境干扰小等优点。在开展小型兽类数量监测中,可采用红外相机技术为主,抽样捕捉调查为辅的方法,以提高调查的可靠性和精确度。
致谢:浙江大学丁平教授为本文提供了红外相机数据及启迪理想模型的构建方法,特此致谢。
[1] Zhao T B,Tao B E,Dong X C,Tang M C.Rodent population survey methodology and its evaluation.Chinese Journal of Vector Biology and Control,2007,18(4):332-334.
[2] He B S,Yuan Z H,Zhang X M,Li X Z,Li J.Infrared triggered camera trap survey technique pand monitoring.Science Journal of Northwest University Online,2009,7(2):1-5.
[3] Karanth K U,Nichols J D,Kumar N S,Hines J E.Assessing tiger population dynamics using photographic capture-recapture sampling.The Ecological Society of America,2006,87(11):2925-2937.
[4] Karanth K U.Estimating tiger Panthera tigirs populations from camera-trap data using capture-recapture models.Biological Conservation,1995,71(3):333-338.
[5] Ma M,Xu F,Chundawat R S,Kubanych J,Wu Y Q,Aizezi,Zhu M H.Camera trapping of snow leopards for the photo capture rate and population size in the Muzat Valley of Tianshan Mountains.Journal of Animal,2006,52(4):788-793.
[6] Ma M,Xu F,Bariushaa M,Wu YQ,Tomas M,Kyle M.Monitoring of population density of snow leopard in Xinjiang.Journal of Ecology and Rural Environment,2011,27(1):79-83.
[7] Rowcliffe J,Field J,Turvey S,and Carbone C.Estimating animal density using camera traps without the need for individual recognition,Journal of Applied Ecology,2008,45(4):1228-1236.
[8] Li L,Chen J H,Ren H B,Mi X C,Yu M J,Yang B.Spatial patterns of Castanopsis eyrei and Schima superba in mid-subtropical broad-leaved evergreen forest in Gutianshan National Reserve,China.Chinese Journal of Plant Ecology,2010,34(3):241-252.
[9] Zhu Y,Zhao G F,Zhang L W,Shen G C,Mi X C,Ren H B,Yu M J,Chen J H,Chen S W,Fang T,Ma K P.Community composition and structure of Gu TianShan Forest dynamic plot in a mid-subtropical evergreen broad-leaved forest,east china.Chinese Journal of Plant Ecology,2008,32(2):262-273.
[10] Huang X Y,Hu D F,Liu W S,Jeff M.Field survey on South China tigers and their habitat evaluation in Yihuang Provincial Reserve in Jiangxi Province.Journal of Beijing Forestry University,2003,25(4):54-58.
[11] Huang X Y,Hu D F,Liu W S,Wang Z C,Liu W S.Field survey of south China tigers and their habitat evaluation in Hupingshan National Reserve.Journal of Zhejiang Forestry College,2004,21(2):180-184.
[12] Huang M X,Jian Y X.Monitoring larger mammals in the Nanzihshian logging road area,Yushan National Park,Taiwan.Taiwan Forestry Science,2007,22(2):135-147.
[13] Wang D M,Zeng D L,Zhang X M,Liu J F.Comparison of microscopic behavior of molecules for real gas and its ideal gas model.Journal of Thermal Science and Technology,2003,2(4):288-291.
[14] Hutchinson,J M C & Waser,P M.Use,misuse and extensions of“ideal gas”models of animal encounter.Biological Reviews,2007,82,335-359.
[15] Shen L L.The research on home range and field of Niviventer confucianus on islands at Thousand Island Lake.Zhejiang Normal University Master Dissertation,2011.
[16] Liu W,Zhong W Q,Wan X R.Advance in the research of home range in rodent.Chinese Journal of Ecology,2002,21(4):36-40.
[17] Wan X R,Zhong W Q.Discussion on the survivorship estimation for the Jolly-Seber model.Chinese Journal of Zoology,2001,36(5):36-39.
[18] Yang C W.Institute on northeast forest five rodent coexistence mechanism.Northeast Forestery University Doctoral Dissertation,2007.
参考文献:
[1] 赵天飙,陶波尔,董希超,唐蒙昌.啮齿动物种群数量调查方法及其评价.中国媒介生物学及控制杂志,2007,18(4):332-334.
[2] 何佰锁,袁朝晖,张希明,李小枝,李健.红外线触发数码相机陷阱技术在大熊猫监测中的应用.西北大学学报(自然科学网络版),2009,7(2):1-5.
[5] 马鸣,徐峰,Chundawat R S,Kubanych J,吴逸群,艾则孜,朱玛洪.利用自动照相术获得天山雪豹拍摄率与个体数量.动物学报,2006,52(4):788-793.
[6] 马鸣,徐峰,Bariushaa M,吴逸群,Tomas M,Kyle M.新疆雪豹种群密度监测方法探讨.生态与农村环境学报,2011,27(1):79-83.
[8] 李立,陈建华,任海保,米湘成,于明坚,杨波.古田山常绿阔叶林优势树种甜槠和木荷的空间格局分析.植物生态学报,2010,34(3):241-252.
[9] 祝燕,赵谷风,张俪文,沈国春,米湘成,任海保,于明坚,陈建华,陈声文,方腾,马克平.古田山中亚热带常绿阔叶林动态监测样地——群落组成与结构.植物生态学报,2008,32(2):262-273.
[10] 黄祥云,胡德夫,刘伟石,Jeff M.宜黄自然保护区华南虎野生种群调查及栖息地评价.北京林业大学学报,2003,25(4):54-58.
[11] 黄祥云,胡德夫,唐小平,王志臣,刘伟石.壶瓶山自然保护区华南虎野外调查及栖息地评价.浙江林学院学报,2004,21(2):180-184.
[12] 黄美秀,简荧云.玉山国家公园楠溪林道较大型哺乳动物之监测.台湾林业科学,2007,22(2):135-147.
[13] 王德明,曾丹苓,张新铭,刘娟芳.实际气体与其对应的理想气体模型微观性质的比较.热科学与技术,2003,2(4):288-291.
[15] 沈良良.千岛湖岛屿社鼠的巢区和领域研究.浙江师范大学硕士学位论文,2011.
[16] 刘伟,钟文勤,宛新荣.啮齿动物巢区研究进展.生态学杂志,2002,21(4):36-40.
[17] 宛新荣,钟文勤.Jolly-Seber法中种群存活率估算的探讨.动物学杂志,2001,36(5):36-39.
[18] 杨春文.东北主要林区森林五种啮齿动物共存机制研究.东北林业大学博士学位论文,2007.
猜你喜欢
环球时报(2022-05-23)2022-05-23
现代园艺(2021年23期)2021-12-01
林业科技(2021年2期)2021-07-08
金桥(2021年4期)2021-05-21
林业勘查设计(2020年1期)2021-01-18
新农业(2020年18期)2021-01-07
电子制作(2019年7期)2019-04-25
野生动物学报(2018年4期)2018-11-23
光学精密工程(2016年3期)2016-11-07
科技视界(2016年23期)2016-11-04
