
长江口横沙东滩围垦潮滩内外大型底栖动物功能群研究
2013-12-09 05:58吕巍巍马长安赵云龙
生态学报 2013年21期
吕巍巍,马长安,余 骥,田 伟,袁 晓,赵云龙,*
(1. 华东师范大学生命科学学院, 上海 200062; 2. 上海市野生动植物保护管理站, 上海 200023)
长江口横沙东滩围垦潮滩内外大型底栖动物功能群研究
吕巍巍1,马长安1,余 骥1,田 伟1,袁 晓2,赵云龙1,*
(1. 华东师范大学生命科学学院, 上海 200062; 2. 上海市野生动植物保护管理站, 上海 200023)
2011年4月(春季)、7月(夏季)、10月(秋季)和12月(冬季),在横沙东滩的围垦潮滩和自然潮滩进行底栖动物的采样调查,进行功能群划分及数量特征分析。在围垦潮滩和自然潮滩分别采集到底栖动物19种和22种。依据食性将其划分为浮游生物食者、植食者、肉食者、杂食者和碎屑食者5种功能群,围垦潮滩各功能群物种数分别为4,5,4,3,3;自然潮滩分别为6,3,5,6,2。围垦潮滩丰度由高到低依次为植食者gt;浮游生物食者gt;肉食者gt;杂食者gt;碎屑食者;自然潮滩为浮游生物食者gt;杂食者gt;肉食者gt;碎屑食者gt;植食者。围垦潮滩季节间仅有浮游生物食者和植食者的物种数存在显著性差异,潮位间仅有植食者的丰度存在显著性差异;自然潮滩季节间仅有浮游生物食者的丰度存在显著性差异,潮位间仅有植食者的物种数和丰度无显著性差异。围垦潮滩主导功能群在各潮位均为浮游生物食者和植食者;自然潮滩主导功能群由高潮带到低潮带变化为浮游生物食者→浮游生物食者和杂食者→杂食者。围垦潮滩浮游生物食者和植食者的Shannon-Wiener多样性指数、Margalef丰富度指数、Pielou均匀度指数较高,而杂食者相对较低;自然潮滩杂食者和肉食者的3种多样性指数较高,而植食者相对较低。以上结果表明:围垦潮滩和自然潮滩底栖动物功能群的组成存在一定的差别;各潮位底栖动物功能群组成的不同与生境密切相关;围垦导致底栖动物群落以及功能群发生变化。
大型底栖动物;功能群;围垦;横沙东滩
随着上海社会经济快速发展,人口数量不断增加,城建用地迅猛扩展,土地需求量与日俱增,人多地少成为制约上海市可持续发展的主要因素[1- 2]。而横沙东滩作为上海湿地资源的重要组成部分,目前已在其周缘湿地进行了围垦造陆工程[3- 4]。围垦能改变滩涂湿地的生态系统结构、过程与功能[5- 6]。底栖动物是湿地生态系统的次级生产者,调节食物网中物质循环和能量流动,既可作为捕食者,摄食一些浮游生物、植物、碎屑等,又可作为被捕食者,为鸟类等高营养级动物提供食物[7]。相关研究表明,底栖动物对周围环境还起到一定的指示作用,但考虑到底栖动物的自由活动能力和对环境变化的敏感程度等因素,当时间和空间发生变化时,底栖动物物种的迁徙、更替或消失有时并不能真实反映该区域生态系统的变化。底栖动物功能群是具有相同生态功能的底栖动物的集合[8- 9],采用功能群方法来分析底栖动物群落结构,则可以简化其复杂的群落内部关系,弱化个别物种的作用,有利于整体把握生态系统的结构和功能[10]。
目前,国内外学者对底栖动物功能群已做了大量的研究[11- 14]。而对横沙东滩底栖动物研究大多集中于物种数、丰度、生物量和多样性等基本群落特征[15- 16],没有涉及底栖动物功能群的研究。本文以底栖动物功能群为研究切入点,根据2011年4个季节的横沙东滩围垦潮滩和自然潮滩底栖动物的调查数据,探讨其功能群的特征以及围垦对横沙东滩底栖动物功能群的影响。
1 材料和方法
1.1 研究地概况
横沙东滩位于横沙岛最东端,地理位置为121°52′—122°20′E,31°10′—31°21′N。围垦潮滩位于横沙东滩南大堤以北,自然潮滩位于南大堤以南,2002年至今促淤圈围工程已经完成了五期,累积施工面积约为124.2 km2。围垦潮滩虽受自然潮水影响,但水动力环境与自然潮滩不同,围垦滩面水位较低,且涨落潮水流速较缓,有利于泥沙沉降,潮滩高程明显高于自然潮滩,内部盐沼植被呈规律分布且长势较好。自然潮滩为围垦保留的自然潮滩,南大堤的建立和长江深水航道的影响,使自然潮滩的高程较低,滩面较围垦潮滩要窄,且水流速较快,有稀疏分布的芦苇和海三棱藨草植被。
1.2 调查方法及数据处理
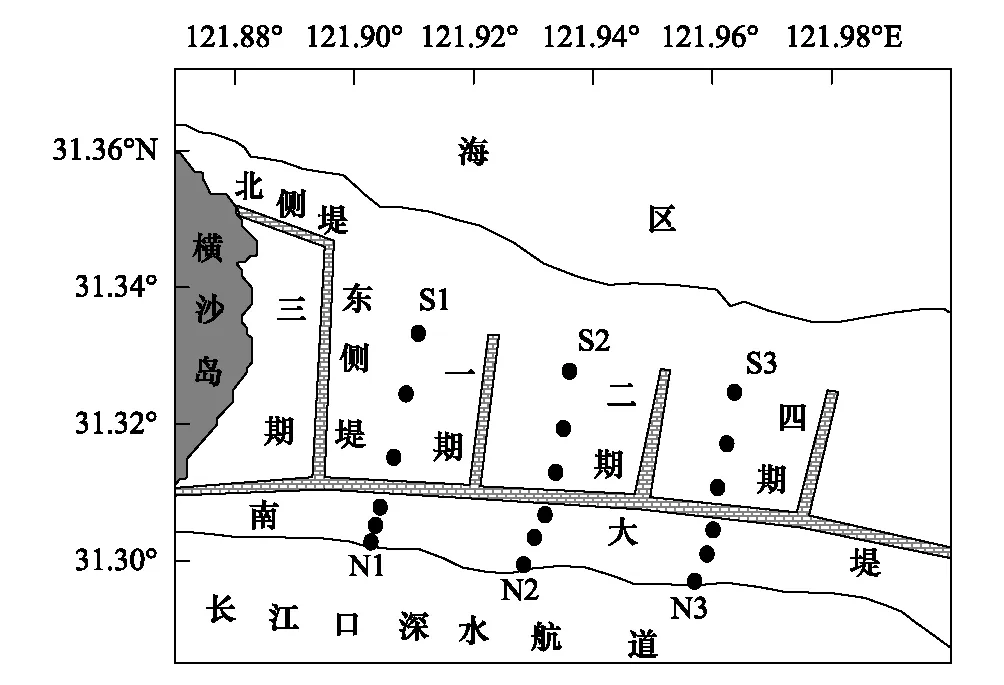
图1 横沙东滩采样断面示意图Fig.1 Map of sampling section in Hengsha East Shoal
根据《海洋调查规范》(GB/T12763.9—2007)底栖动物调查方法及横沙东滩的具体生境状况,于2011年4月(春季)、7月(夏季)、10月(秋季)和12月(冬季)对横沙东滩围垦潮滩和自然潮滩的底栖动物进行调查。围垦潮滩一期、二期和四期工程各设1条样带(S1、S2、S3),自然潮滩设置3条对照样带(N1、N2、N3)。根据高程、植被分布的情况,每条样带分别设置高、中、低3个样点,每个样点定量取样重复4—8次,样方为25 cm×25 cm×30 cm,40目筛网筛洗后拣取底栖动物,同时在周围尽可能地获取定性样本。所获的底栖动物用75%的酒精固定,带回实验室进行鉴定、计数和称量。具体样点设置见图1。
采用Shannon-Wiener多样性指数(H′)、Pielou均匀度指数(J′)、Margalef丰富度指数(d)进行大型底栖动物功能群多样性指数的分析:
H′=-∑(ni/N)log2(ni/N)
J′=H′/log2S
d= (S-1)/log2N
式中,ni为样品中第i种的个体数;N为该水域所有个体总数之和;S为所采样品的种类总数。采用分析软件SPSS16.0对数据进行无重复双因素方差分析。
根据底栖动物食性将其划分为5个功能群,即浮游生物食者(Pl)、植食者(Ph)、肉食者(C)、杂食者(O)、碎屑食者(D)[17- 19]。
2 结果
2.1 底栖动物的功能群组成
功能群的组成和数量见表1。围垦潮滩各功能群种类组成为植食者(5种)gt;浮游生物食者(4种)=肉食者(4种)gt;杂食者(3种)=碎屑食者(3种);自然潮滩则为浮游生物食者(6种)=杂食者(6种)gt;肉食者(5种)gt;植食者(3种)gt;碎屑食者(2种)。围垦潮滩各功能群数量占总数百分比为植食者(70.27%)gt;浮游生物食者(19.26%)gt;肉食者(3.87%)gt;杂食者(3.35%)gt;碎屑食者(3.25%);自然潮滩为浮游生物食者(43.77%)gt;杂食者(26.31%)gt;肉食者(13.76%)gt;碎屑食者(9.98%)gt;植食者(6.18%)。
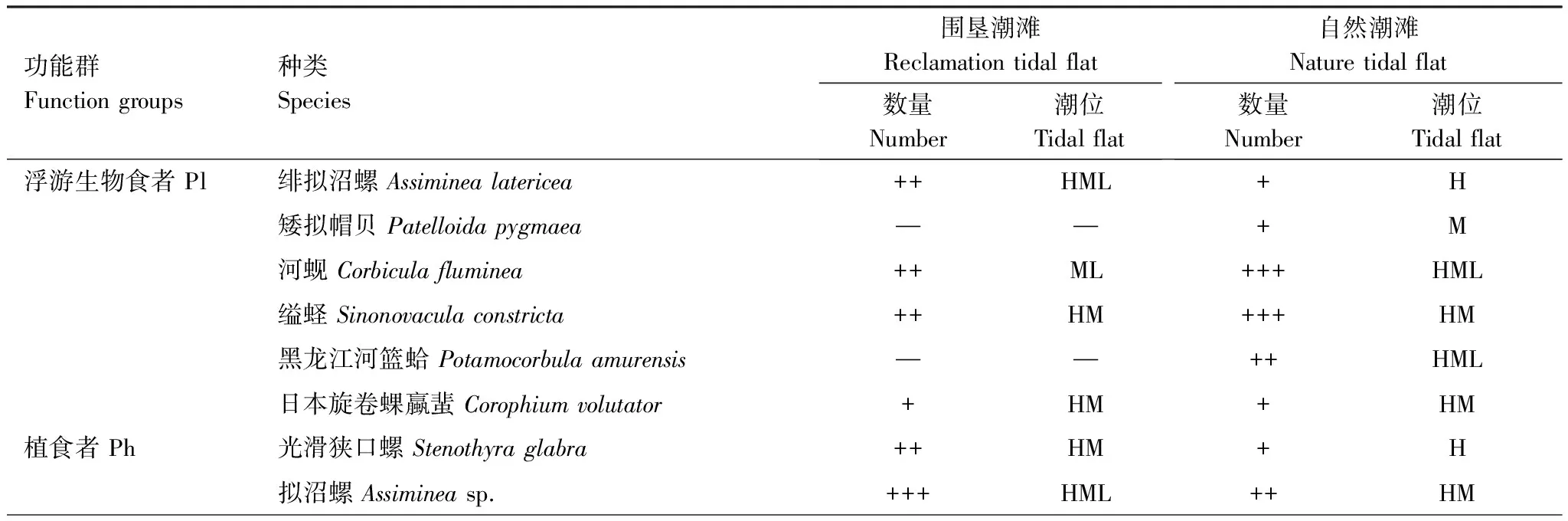
表1 横沙东滩大型底栖动物的物种组成及其功能群
续表
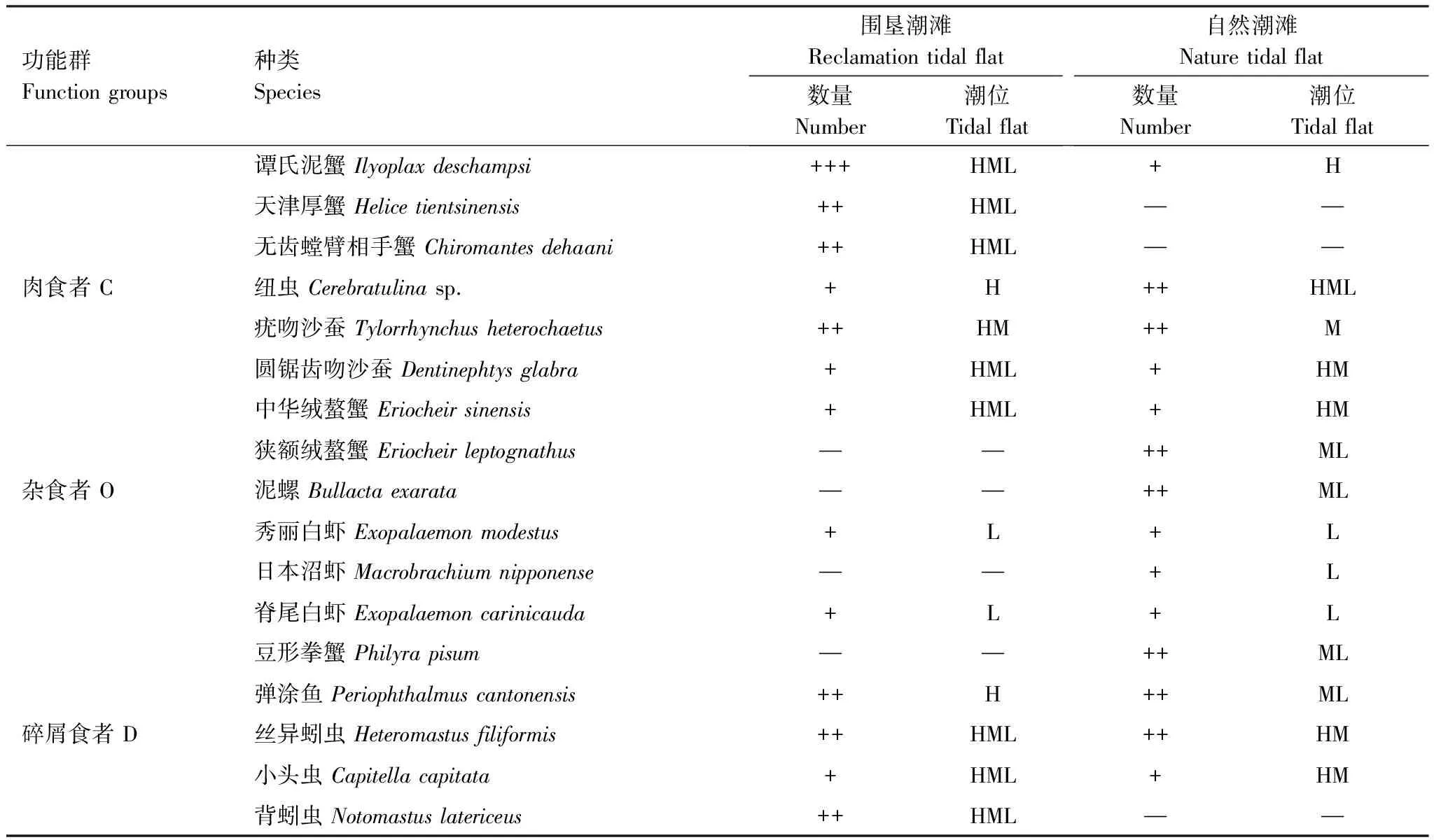
功能群Functiongroups种类Species围垦潮滩Reclamationtidalflat数量Number潮位Tidalflat自然潮滩Naturetidalflat数量Number潮位Tidalflat谭氏泥蟹Ilyoplaxdeschampsi+++HML+H天津厚蟹Helicetientsinensis++HML——无齿螳臂相手蟹Chiromantesdehaani++HML——肉食者C纽虫Cerebratulinasp.+H++HML疣吻沙蚕Tylorrhynchusheterochaetus++HM++M圆锯齿吻沙蚕Dentinephtysglabra+HML+HM中华绒螯蟹Eriocheirsinensis+HML+HM狭额绒螯蟹Eriocheirleptognathus——++ML杂食者O泥螺Bullactaexarata——++ML秀丽白虾Exopalaemonmodestus+L+L日本沼虾Macrobrachiumnipponense——+L脊尾白虾Exopalaemoncarinicauda+L+L豆形拳蟹Philyrapisum——++ML弹涂鱼Periophthalmuscantonensis++H++ML碎屑食者D丝异蚓虫Heteromastusfiliformis++HML++HM小头虫Capitellacapitata+HML+HM背蚓虫Notomastuslatericeus++HML——
Pl: Planktophagous group, Ph: Phytophagous group, C: Camivorous group, O: Omnivorous group, D: Detritivorous group;+: 分别表示物种个体数lt;总个体数的1%; ++: 占总个体数1%—10%; +++:gt;总个体数的10%,—:定量取样未获得;H: 高潮带; M: 中潮带; L: 低潮带
2.2 底栖动物功能群的时空变化
5种功能群在横沙东滩围垦潮滩和自然潮滩的分布各不相同。围垦潮滩的主导功能群为植食者和浮游生物食者,其功能群之和占围垦潮滩全部物种种类和丰度的47.37%和89.34%;而自然潮滩的主导功能群为浮游生物食者和杂食者,其功能群之和占自然潮滩全部物种种类和丰度的54.55%和70.27%。各功能群的物种数和丰度沿高程分布各不相同。具体结果见表2。
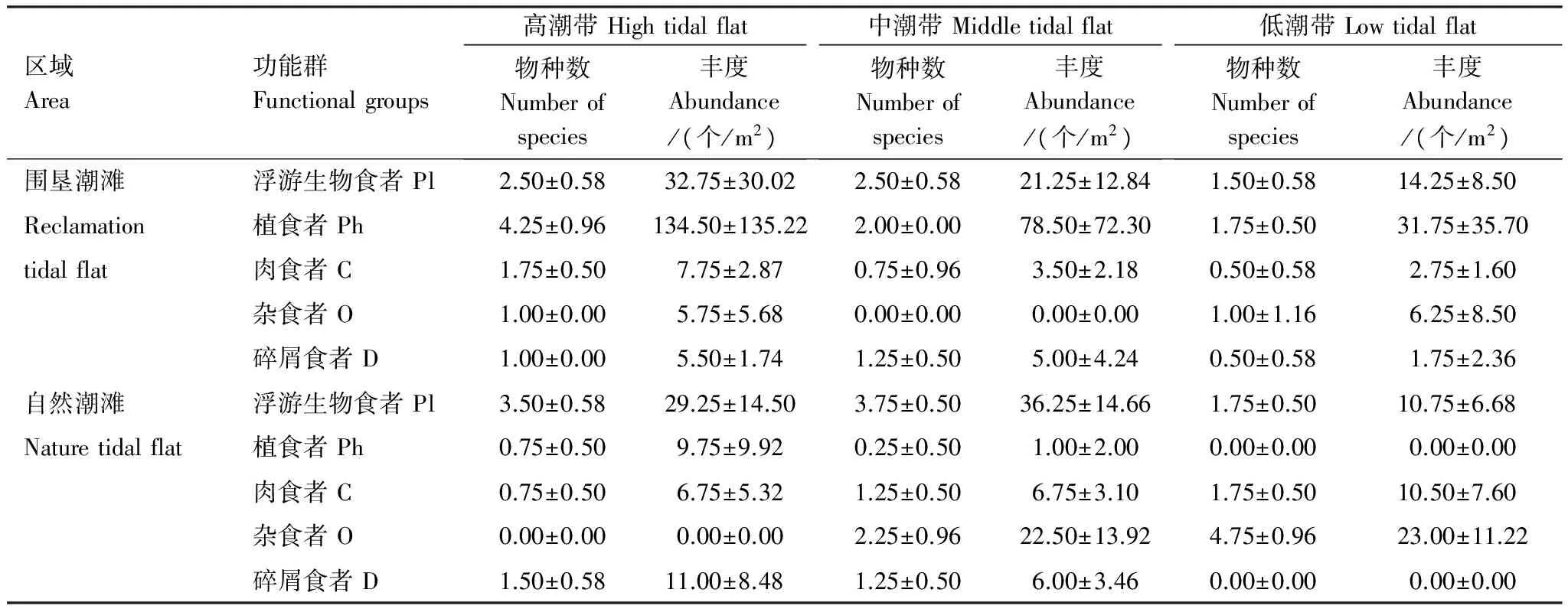
表2 横沙东滩3个潮位大型底栖动物各功能群的物种数和栖息丰度(平均值±标准差)
围垦潮滩和自然潮滩底栖动物功能群的物种数和丰度分别进行季节-潮位无重复双因素方差分析。季节因素来看,围垦潮滩除了浮游生物食者(F3,6=5.68,Plt;0.05)和植食者(F3,6=5.56,Plt;0.05)的丰度在4个季节之间存在显著性差异外,其余无显著性差异,自然潮滩也只有浮游生物食者的丰度(F3,6=4.81,Plt;0.05)在各季节间存在显著性差异,而围垦潮滩和自然潮滩各功能群的物种数在各季节间均无显著性差异。潮位因素来看,围垦潮滩仅有植食者的物种数(F2,6=30.33,Plt;0.01)在不同潮位之间存在极显著性差异,而各功能群的丰度在围垦潮滩不同潮位之间无显著性差异;自然潮滩与围垦潮滩不同,除了植食者的丰度(F2,6=4.13,Pgt;0.05)和物种数(F2,6=4.20,Pgt;0.05)无显著性差异外,其它各功能群的丰度和物种数在各潮位之间具有显著或极显著性差异。具体结果见表3。
自然潮滩和围垦潮滩的主导功能群在各潮位之间呈现不同特点。自然潮滩高潮带的主导功能群为浮游生物食者,中潮带的主导功能群为浮游生物食者和杂食者,而低潮带的主导功能群为杂食者,各潮位主导功能群占所有物种种类和丰度的百分比为:高潮带,40.00%和51.54%;中潮带,61.54%和81.03%;低潮带,60.00%和51.98%。围垦潮滩植食者和浮游生物食者是3个潮位的主导功能群,其功能群之和占每个潮位物种种类和丰度的百分比为:高潮带,53.33%和89.79%;中潮带,57.14%和92.15%;低潮带,50.00%和81.06%。
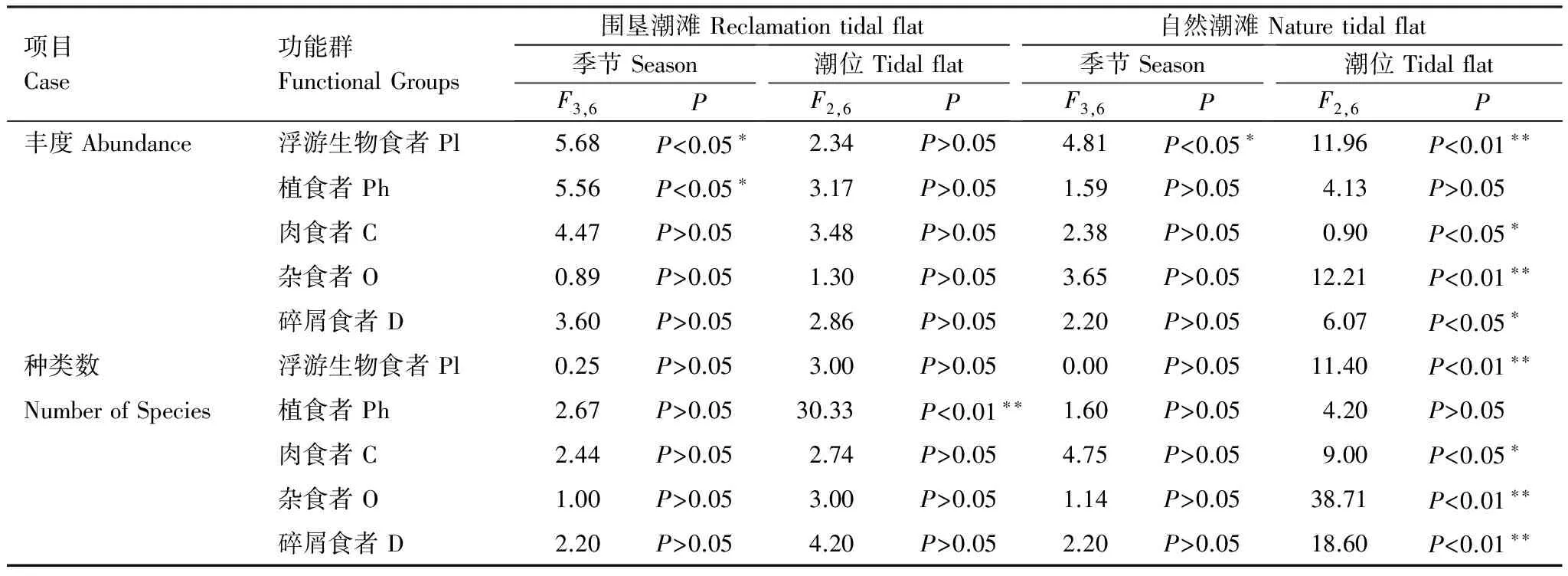
表3 各功能群的季节潮位无重复双因素方差分析
*: 差异显著,Plt;0.05; **: 差异极显著,Plt;0.01
2.3 底栖动物功能群的多样性特征
围垦潮滩和自然潮滩多样性指数变化趋势不同。围垦潮滩浮游生物食者和植食者的Shannon-Wiener多样性指数、Margalef丰富度指数、Pielou均匀度指数较高,而杂食者相对较低;自然潮滩杂食者和肉食者的3种多样性指数较高,而植食者相对较低。
底栖动物各功能群在不同区域呈现出不同的多样性特征。围垦潮滩中植食者和肉食者多样性指数在高潮带最高;浮游生物食者和碎屑食者多样性指数在中潮带最高;杂食者在高潮带和中潮带分别采到1种和0种,而在低潮带多样性指数相对较高。自然潮滩中浮游生物食者和碎屑食者的多样性指数在中潮带最高;肉食者和杂食者在低潮带多样性指数相对较高;植食者在高潮带和中潮带都仅采到1个物种,多样性指数都为0,并且在低潮带无分布。杂食者和碎屑食者也分别在高潮带和低潮带没有物种分布。具体见表4。
3 讨论
3.1 自然潮滩底栖动物功能群分布的时空差异及人为干扰的影响
由于盐度、温度、沉积物类型和水动力条件等环境因子在河口生态系统中呈现一定的梯度变化,使得底栖动物功能群的分布存在时空上的差异[20]。一些研究表明,在潮间带,影响底栖动物功能群时空变化的两个最主要的因素是季节和潮位[21]。从本文的研究结果来看,自然潮滩仅有浮游生物食者的丰度存在季节性的显著性差异,而各功能群的物种数在季节之间无显著性差异。由于高程的改变,导致自然潮滩主导功能群从高潮带到低潮带发生明显的变化为:浮游生物食者→浮游生物食者和杂食者→杂食者,且各功能群中除了植食者的物种数和丰度在各潮位无显著性差异外,另外几种功能群均存在显著性差异。因此,可以判断各潮位生境的不同是导致横沙东滩自然潮滩底栖动物功能群物种数和丰度变化的主导因素。河口潮间带生境的空间异质性直接影响底栖动物功能群分布的区域性特征,沿着高程梯度,自然潮滩生境表现出明显的空间生态系列格局:中潮带属于潮滩湿地演替的高级阶段[22],其底栖动物功能群的物种数、丰度和多样性指数均是自然潮滩最高;低潮带海陆交汇最频繁,较强的潮汐频率使底栖动物栖息环境变得很不稳定,所以功能群的物种数、丰度和多样性指数较中潮带要低,属于潮滩湿地演替的初级阶段;高潮带大部分芦苇和海三棱藨草滩已经被围垦,保留的潮滩仅为高潮带的下缘部分,滩面宽度变得很狭窄,这些因素可能是导致底栖动物功能群的物种数、丰度和多样性指数都为自然潮滩最低的原因。
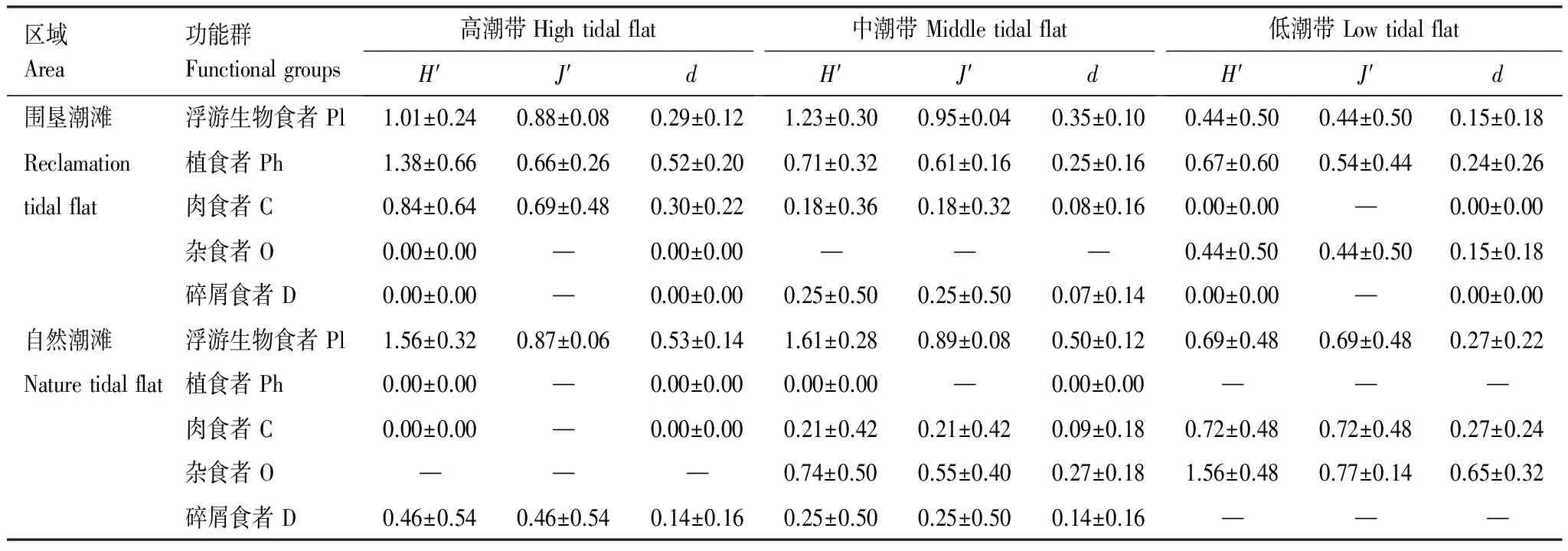
表4 横沙东滩大型底栖动物功能群的多样性指数(平均值±标准差)
—: 无数据;H′: Shannon-Wiener多样性指数 Shannon-Wiener diversity index;J′: Pielou均匀度指数 Pielou evenness index;d: Margalef丰富度指数 Margalef richness index
当生态系统的功能受到损害的时候,功能群的组成和类型也会发生相应的变化[23]。本研究结果表明,高潮带和中潮带主导功能群主要由浮游生物食者和杂食者构成,植食者的物种数和丰度却较少,这与其他学者对长江口的研究结果不同[24]。可能的原因是横沙东滩形成时间较晚,围垦建设的南大堤将高潮带大部分框围入围垦潮滩,保留的潮间带滩面最宽处也仅有两公里左右,为底栖动物提供的栖息空间很有限,底栖动物功能群的构成与绵延数公里、植被呈明显带状分布的崇明东滩相比更为简单[25]。同时,围垦保留的盐沼植被也较为稀疏,导致植食者相对较少。通常在长江口实施的围垦工程初期,对淤积岸段潮滩的框围会加快堤前滩地的淤积[26],但横沙东滩植被覆盖率低,较强的风和浪,流域来沙量减少以及长江深水航道工程都可能影响泥沙沉降,导致自然潮滩高程升高缓慢。潮滩高程直接影响盐沼植被,尤其是海三棱藨草的生活史状况,据相关研究表明,海三棱藨草在高程较低时常采取无性繁殖,不利于种群的快速扩散[27]。因此,这有可能导致横沙东滩自然潮滩的初级生产力较低,且这种状况会持续很长时间,不利于底栖动物功能群的构成和恢复。
3.2 围垦滩涂对底栖动物功能群的影响
滩涂湿地围垦之后,受到一些生物因素和非生物因素的影响,底栖动物功能群的构成会发生相应的变化。围垦潮滩和自然潮滩均有5个功能群,但围垦潮滩主导功能群为浮游生物食者和植食者,且在各潮位之间无变化,而自然潮滩则由高潮带的浮游生物食者向低潮带的杂食者演变。方差分析来看,围垦潮滩仅有植食者的丰度存在显著性差异,而自然潮滩大部分功能群的物种数和丰度都在潮位之间存在显著性差异。这说明围垦导致底栖动物功能群在潮位之间趋同。横沙东滩围垦多年,滩面宽度大约是自然潮滩的2—3倍,这就为底栖动物提供了更多的生存空间。但长时间的促淤使得围垦潮滩的高程逐年升高,各潮位之间高程差异已不明显,盐度、潮汐动力等环境因素的改变使区内底栖动物功能群的分布格局较自然潮滩发生明显的变化,尤其是潮水流速减慢以及围垦后盐沼植被迅速扩张,为底上活动的植食者以及滤食性的浮游生物食者提供了丰富的食物来源和稳定的生存环境,使这两种功能群的丰度和多样性在围垦潮滩均处于较高水平。潮滩围垦之后,原本为光滩的低潮带也开始有芦苇和海三棱藨草植被呈斑块状出现,同时一些植食者由于种群扩张的速度较快,导致高中潮带的生存空间不足而下迁到低潮带,浮游生物食者和植食者比例的增加,必然导致其余几种功能群比例的减少,尤其是低潮带原主导功能群杂食者的生态位已经逐渐被植食者取代。
湿地生态系统食物网构成较为复杂,作为主要的消费者,底栖动物功能群的多样性直接影响着潮间带食物网的结构和功能[28]。自然潮滩虽然受到筑堤工程的影响,但底栖动物功能群的分布较围垦潮滩更具长江口潮间带的特征,尤其是杂食者和肉食者的丰度和多样性要明显高于围垦潮滩;而促淤之后的围垦潮滩仅有植食者的多样性高于自然潮滩,其丰度也达到总丰度的70.27%。这说明围垦导致原始潮滩的生境功能发生改变,更适宜于植食者的生存。根据相关研究,底栖动物内部的物质和能量流动可以简单的归纳为3条路线,即植物→植食者→捕食者;浮游生物→浮游生物食者→捕食者;碎屑→碎屑食者→捕食者[29]。而湿地生态系统中广泛存在着营养级联效应,这种下行效应的强度影响着湿地生态系统食物链中各营养级的变化,进而决定着底栖动物各功能群的发展状况[30]。围垦潮滩一些杂食性和肉食性底栖动物由于环境的改变数量下降直至消失,而捕食者的减少使得植食者数量相对增加,这种营养级联效应也会间接作用于潮间带的初级生产者,最终可能导致围垦潮滩初级生产力不断下降。因此,围垦滩涂生态功能的不平衡,人为的生态修复干预十分必要。
3.3 生态修复
底栖动物构成了湿地生态系统中的底栖亚系统,其功能群可作为生态系统受损时的压力指标来反映生态系统的基本动态[31]。研究湿地生态系统中底栖动物功能群的构成对生态修复有重要的意义。根据横沙东滩的实际情况,增加消费者特别是食物链中等级较高的消费者的多样性有利于控制围垦潮滩中植食者的数量,避免单一主导功能群造成群落结构的不平衡,使围垦潮滩的初级生产力保持在一个较高的水平。同时,适当降低围垦高程的幅度,恢复低潮带光滩原貌,重新投放泥螺等低潮带经济物种,使狭额绒螯蟹等活动能力较强的低潮带常见物种迁移返回重新利用新的沉积环境。综上所述,湿地围垦对底栖动物功能群的影响是十分明显的,在进行湿地开发的同时,注重对底栖动物特别是生态经济价值较高的物种的保护,做到经济发展与维持生态环境永续相结合,这是需要长期进行研究的课题。
[1] Ma C A, Xu L L, Tian W, Lv W W, Zhao Y L. The influence of a reclamation project on the macrobenthos of an East Nanhui tidal flat. Acta Ecologica Sinica, 2012, 32(4): 1007- 1015.
[2] Zhang B, Yuan X, Pei E L, Niu J Y, Heng N N, Wang T H. Change of waterbird community structure after the intertidal mudflat reclamation in the Yangtze River Mouth: a case study of Nanhui Dongtan area. Acta Ecologica Sinica, 2011, 31(16): 4599- 4608.
[3] Sang Y Y, Yu Z Y, Jin L. Natural evolvement and effect of project in the Hengsha East Shoal of the Changjiang River estuarine. Donghai Marine Science, 2003, 21(3): 14- 23.
[4] Du J L, Jiang L P, Yang S L. GIS analysis of the natural evolvement of the Hengsha East Shoal of the Yangtze River and the infects of projects in the last thirty years. Marine Science Bulletin, 2007, 26(5): 43- 48.
[5] Niu J Y, Heng N N, Zhang B, Yuan X, Wang T H. Waterbird habitat-selection during winter and spring in reclaimed coastal wetlands in Nanhui Dongtan, Shanghai. Zoological Research, 2011, 32(6): 624- 630.
[6] Ge B M, Bao Y X, Zheng X. Structure of macrobenthic communities in different habitats at the diked tidal flat in winter. Zoological Research, 2005, 26(1): 47- 54.
[7] Zhang C L, Xu B D, Ren Y P, Xue Y, Ji Y P. The spatio-temporal change in the secondary production of macrozoobenthos in the intertidal zone of Jiaozhou Bay. Acta Ecologica Sinica, 2011, 31(17): 5071- 5080.
[8] Li Y, Li G Y, Mu J P, Sun S C. Effects of consumer diversity on food web structure and ecosystem functioning: current knowledge and perspectives. Acta Ecologica Sinica, 2008, 28(1): 388- 398.
[9] Jeanette L S, Mike A K, Anthony J S H, John I S. Can functional groups be used to indicate estuarine ecological status? Hydrobiologia, 2007, 588(1): 45- 58.
[10] Ge B M, Bao Y X, Cheng H Y, Li H H, Hu Z Y. The trophic functional groups and trophic levels of the macrobenthic community at the east tidal flat of Lingkun Island. Acta Ecologica Sinica, 2008, 28(10): 4796- 4804.
[11] Mucha A P, Costa M H. Macrozoobenthic community structure in two Portuguese estuaries: relationship with organic enrichment and nutrient gradients. Acta Oecologica, 1999, 20(4): 363- 376.
[12] Talley T S, Dayton P K, Ibarra-Obando S E. Tidal flat macrofaunal communities and their associated environments in estuaries of southern California and northern Baja California, Mexico. Estuaries and Coasts, 2000, 23(1): 97- 114.
[13] Xin J H, Ren Y P, Xu B D, Zhang C L, Xue Y, Ji Y P. Macrozoobenthos functional groups in intertidal flat of northwest Jiaozhou Bay. Chinese Journal of Applied Ecology, 2011, 22(7): 1885- 1892.
[14] Bao Y X, Hu Z Y, Li H H, Ge B M, Cheng H Y. Seasonal variation and functional groups of macrobenthic communities at diked and natural tidal flat, Lingkun Island, China. Acta Zoologica Sinica, 2008, 54(3): 416- 427.
[15] Yuan X Z, Lu J J. Studies on zoobenthos resources in the islands of the Changjiang Estuary. Journal of Natural Resources, 2001, 16(1): 37- 41.
[16] Tao S R, Jiang L F, Wu J H, Zhao B, Li B. Community characteristics and seasonal changes of macrozoobenthos in intertidal zones of Hengsha and Changxing islands at Yangtze River Estuary. Chinese Journal of Ecology, 2009, 28(7): 1345- 1350.
[17] Yuan X Z, Lu J J, Liu H. Distribution pattern and variation in the functional groups of zoobenthos in the Changjiang Estuary. Acta Ecologica Sinica, 2002, 22(12): 2054- 2062.
[18] Steneck R S, Watling L. Feeding capabilities and limitation of herbivorous molluscs: a functional group approach. Marine Biology, 1982, 68(3): 299- 319.
[19] Fauchald K, Jumars P A. The diet of worms: a study of polychaete feeding guilds. Oceanography and Marine Biology, An Annual Review, 1979, 17: 193- 284.
[20] Xu H F, Zhao Y L. The Report of Scientific Survey of the Chongming Dong Tan Migratory Bird Nature Reserve of Shanghai. Beijing: China Forestry Publishing House, 2005: 75- 115.
[21] You Z J, Tao L, Jiao H F, Shi H X, Lou D. A survey of macrobenthos in the Xiangshan Bay. Oceanologia et Limnologia Sinica, 2011, 42(3): 431- 435.
[22] Li H H, Bao Y X, Hu Z Y, Ge B M. Seasonal dynamics of macrobenthic functional groups and trophic levels in the bridge construction zone at the South Bank of Hangzhou Bay, China. Acta Zoologica Sinica, 2007, 53(6): 1011- 1023.
[23] Tilman D, Knops J, Wedin D, Reich P, Ritchie M, Siemann E. The influence of functional diversity and composition on ecosystem processes. Science, 1997, 277(5330): 1300- 1302.
[24] Zhu X J, Lu J J. Functional groups of zoobenthos in the intertidal zone of Jiuduansha, the Yangtze River Estuary. Zoological Research, 2003, 24(5): 355- 361.
[25] An C G, Zhao Y L, Lin L, Li J Y, Cui L L, Ma C Y. The biodiversity of macrobenthos of intertidal zone on Chongming Island in summer. Acta Ecologica Sinica, 2008, 28(2): 577- 586.
[26] Chen C J. Changes in mud flat after tideland being enclosed. Marine Science Bulletin, 1990, 9(3): 69- 74.
[27] Sun S C, Cai Y L, Liu H. Biomass allocation ofScirpusmariqueteralong an elevational gradient in a salt marsh of the Yangtse River Estuary. Acta Botanica Sinica, 2001, 43(2): 178- 185.
[28] Simanonok M P, Anderson C B, Pastur G M, Lencinas M V, Kennedy J H. A comparison of impacts from silviculture practices and North American beaver invasion on stream benthic macroinvertebrate community structure and function inNothofagusforests of Tierra del Fuego. Forest Ecology and Management, 2011, 262(2): 263- 269.
[29] Zhang Q T, Hu G K, Ni R, Hu Y, Guo C. A preliminary study on the trophic structure of macrobenthos in the Tanggu littoral zone. Transactions of Oceanology and Limnology, 2005, 17(3): 73- 78.
[30] Halpern B S, Borer E T, Seabloom E W, Shurin J B. Predator effects on herbivore and plant stability. Ecology Letters, 2005, 8(2): 189- 194.
[31] Mouquet N, Moore J L, Loreau M. Plant species richness and community productivity: why the mechanism that promotes coexistence matters. Ecology Letters, 2002, 5(1): 56- 65.
参考文献:
[1] 马长安, 徐霖林, 田伟, 吕巍巍, 赵云龙. 围垦对南汇东滩湿地大型底栖动物的影响. 生态学报, 2012, 32(4): 1007- 1015.
[2] 张斌, 袁晓, 裴恩乐, 牛俊英, 衡楠楠, 王天厚.长江口滩涂围垦后水鸟群落结构的变化——以南汇东滩为例. 生态学报, 2011, 31(16):4599- 4608.
[3] 桑永尧, 虞志英, 金寥. 长江河口横沙东滩自然演变及工程影响. 东海海洋, 2003, 21(3): 14- 23.
[4] 杜景龙, 姜俐平, 杨世伦. 长江口横沙东滩近30年来自然演变及工程影响的GIS分析. 海洋通报, 2007, 26(5): 43- 48.
[5] 牛俊英, 衡楠楠, 张斌, 袁晓, 王天厚. 上海市南汇东滩围垦后海岸带湿地冬春季水鸟生境选择. 动物学研究, 2011, 32(6): 624- 630.
[6] 葛宝明, 鲍毅新, 郑祥. 围垦滩涂不同生境冬季大型底栖动物群落结构. 动物学研究, 2005, 26(1): 47- 54.
[7] 张崇良, 徐宾铎, 任一平, 薛莹, 纪毓鹏. 胶州湾潮间带大型底栖动物次级生产力的时空变化. 生态学报, 2011, 31(17): 5071- 5080.
[8] 李妍, 李国勇, 慕军鹏, 孙书存. 消费者多样性对食物网结构和生态系统功能的影响. 生态学报, 2008, 28(1): 388- 398.
[10] 葛宝明, 鲍毅新, 程宏毅, 李欢欢, 胡知渊. 灵昆岛东滩潮间带大型底栖动物功能群及营养等级构成. 生态学报, 2008, 28(10): 4796- 4804.
[13] 辛俊宏, 任一平, 徐宾铎, 张崇良, 薛莹, 纪毓鹏. 胶州湾西北部潮滩湿地大型底栖动物功能群. 应用生态学报, 2011, 22(7): 1885- 1892.
[14] 鲍毅新, 胡知渊, 李欢欢, 葛宝明, 程宏毅. 灵昆东滩围垦区内外大型底栖动物季节变化和功能群的比较. 动物学报, 2008, 54(3): 416- 427.
[15] 袁兴中, 陆健健. 长江口岛屿湿地的底栖动物资源研究. 自然资源学报, 2001, 16(1): 37- 41.
[16] 陶世如, 姜丽芬, 吴纪华, 赵斌, 李博. 长江口横沙岛、长兴岛潮间带大型底栖动物群落特征及其季节变化. 生态学杂志, 2009, 28(7): 1345- 1350.
[17] 袁兴中, 陆健健, 刘红. 长江口底栖动物功能群分布格局及其变化. 生态学报, 2002, 22(12): 2054- 2062.
[20] 徐宏发, 赵云龙. 上海市崇明东滩鸟类自然保护区科学考察集. 北京: 中国林业出版社, 2005: 75- 115.
[21] 尤仲杰, 陶磊, 焦海峰, 施慧雄, 楼丹. 象山港大型底栖动物功能群研究. 海洋与湖沼, 2011, 42(3): 431- 435.
[22] 李欢欢, 鲍毅新, 胡知渊, 葛宝明. 杭州湾南岸大桥建设区域潮间带大型底栖动物功能群及营养等级的季节动态. 动物学报, 2007, 53(6): 1011- 1023.
[24] 朱晓君, 陆健健. 长江口九段沙潮间带底栖动物的功能群. 动物学研究, 2003, 24(5): 355- 361.
[25] 安传光, 赵云龙, 林凌, 李嘉尧, 崔丽丽, 马春艳. 崇明岛潮间带夏季大型底栖动物多样性. 生态学报, 2008, 28(2): 577- 586.
[26] 陈才俊. 围滩造田与淤泥质潮滩的发育. 海洋通报, 1990, 9(3): 69- 74.
[27] 孙书存, 蔡永立, 刘红. 长江口盐沼海三棱藨草在高程梯度上的生物量分配. 植物学报, 2001, 43(2): 178- 185.
[29] 张青田, 胡桂坤, 倪蕊, 胡洋, 郭宠. 塘沽潮间带大型底栖动物营养结构的初步分析. 海洋湖沼通报, 2005, 17(3): 73- 78.
MacrobenthicfunctionalgroupsatthereclamationandnaturaltidalflatsofHengshaEastShoal,theEstuaryofChangjiangRiver
LV Weiwei1, MA Chang′an1, YU Ji1, TIAN Wei1, YUAN Xiao2, ZHAO Yunlong1,*
1SchoolofLifeScience,EastChinaNormalUniversity,Shanghai200062,China2ShanghaiWildlifeConservationManagementStation,Shanghai200023,China
Functional groups of macrobenthos are vital for the assessment of the changes of ecological functions of wetland driven by natural and human factors. In order to get a better understanding of the functional groups of macrobenthos, the surveys were conducted in the reclamation tidal and the natural tidal flats of Hengsha East Shoal in April, July, October and December 2011. The results demonstrated that 19 species and 22 species were found in and outside the dyke, respectively. Based on their food preferences, the macrobenthos were classified into 5 functional groups: Planktophagous group, Phytophagous group, Camivorous group, Omnivorous group and Detritivorous group. The number of species in each functional group was 4, 5, 4, 3, 3 in the reclamation tidal flat, and 6, 3, 5, 6, 2 in the natural tidal flat. The abundance of each functional group in proper order was Phytophagous gt; Planktophagous gt; Camivorous gt; Omnivorous gt; Detritivorous in the reclamation tidal flat, and Planktophagous gt; Omnivorous gt; Camivorous gt; Detritivorous gt; Phytophagous in the natural tidal flat. The species number and abundance of each functional group were analyzed by the two-way ANOVA method. For the reclamation tidal flat, there was no significant difference among seasons for all of the species number and abundance except for the species number of Planktophagous group and Phytophagous group. Similarly, no significant difference was found among the tidal flats for the species number and abundance of each functional group except for the abundance of Phytophagous group. For the natural tidal flat, the difference was significant for the species number of Planktophagous group among the seasons, but not for others. However, most of the functional groups had significant difference among the tidal flats except for the species number and abundance of Phytophagous group. The Planktophagous group and Phytophagous group were the dominant functional groups at three tidal flats of the reclamation tidal flat. The dominant functional groups of the natural tidal flat, however, were Planktophagous at high tidal flat, Planktophagous and Omnivorous at middle tidal flat, Omnivorous at low tidal flat, respectively. In the reclamation tidal flat, Planktophagous group and Phytophagous group had the higher diversity indices (Shannon-Wiener index, Margalef index, Pielou index), while Omnivorous group had the lower diversity indices. In the natural tidal flat, Camivorous group and Omnivorous group had the higher diversity indices, while Phytophagous group had the lower diversity indices.
In conclusion, the composition of functional groups between the reclamation tidal flat and the natural tidal flat was distinctly different. And the changes of the functional groups among the tidal flats were often associated with habitats. The reclamation had resulted in the variations of the communities and functional groups in macrobenthos.
macrobenthos; functional groups; reclamation; Hengsha East Shoal
国家科技支撑计划资助项目(2010BAK69B14);上海市科委重大资助项目(09DZ120010A,10DZ1200700);国家公益性行业(农业)科研专项资助项目(201203065)
2012- 07- 10;
2013- 06- 21
*通讯作者Corresponding author.E-mail: ylzhao@ bio.ecnu.edu.cn
10.5846/stxb201207100969
吕巍巍,马长安,余骥,田伟,袁晓,赵云龙.长江口横沙东滩围垦潮滩内外大型底栖动物功能群研究.生态学报,2013,33(21):6825- 6833.
Lv W W, Ma C A, Yu J, Tian W, Yuan X, Zhao Y L.Macrobenthic functional groups at the reclamation and natural tidal flats of Hengsha East Shoal, the Estuary of Changjiang River.Acta Ecologica Sinica,2013,33(21):6825- 6833.
猜你喜欢
海洋通报(2022年5期)2022-11-30
小主人报(2022年18期)2022-11-17
海洋通报(2021年1期)2021-07-23
科学之谜(2021年4期)2021-07-09
海洋通报(2020年3期)2020-10-20
西湖(2020年8期)2020-08-13
山东煤炭科技(2020年4期)2020-05-12
简帛(2019年2期)2019-11-03
科学之友(2017年3期)2017-03-24
中国工程咨询(2015年5期)2015-02-16
