
七叶一枝花种子萌发特性研究
2013-12-09 08:36张旺凡沈素贞梁文斌
中国野生植物资源 2013年5期
张旺凡,沈素贞,梁文斌,张 睿
(1.湖南中医药高等专科学校,湖南株洲412012;2.中南林业科技大学,湖南长沙410004)
七叶一枝花[Paris polyphylla Smith var.chinensis(Franch.)Hara]又名华重楼,属延龄草科重楼属多年生草本植物,是《中国药典》2005年版收载的药用重楼品种之一[1],分布于湖南、江西、浙江等地,多生于海拔700~1 300 m的山坡、灌木林或溪边、湿地肥沃的沙壤土和腐殖质土壤[2]。现代药理研究表明该中草药具有抗肿瘤、止血、止咳平喘、免疫调节、抗菌消炎、镇静、镇痛及调节心脑血管等作用[3],是云南白药、宫血宁片、三七血伤宁胶囊、莲花片、止痛抗癌丸、季德胜蛇药片和抗病毒冲剂等中成药的主要原料,具有较高的经济效益和社会效益。
随着研究的深入,七叶一枝花的药用价值越来越受到人们的重视,应用范围不断拓展,市场需求量越来越大。因其生长周期长,对生态环境要求高,供求矛盾日益突出,地下茎价格由2008年每公斤140元上升到2012年每公斤800元以上,4年翻了近6倍[3],受利益驱动,人们对其无节制地采挖,已造成野生资源枯竭,因而人工栽培成为解决供需矛盾的主要途径。常规采用根茎繁殖,但种源稀缺且用量大,难以满足市场需求;种子繁殖具有繁殖系数大、生产成本低等优点,但其休眠期长,发芽率低[4],因此人工打破种子休眠,提高发芽率,缩短休眠周期才是目前解决七叶一枝花种苗奇缺的有效途径。目前对七叶一枝花的研究主要集中在药理活性等方面而对其种子萌发特性的研究尚未见报道。为此本文对七叶一枝花的种子采取变温层积与激素处理相结合打破休眠并对其萌发温度、培养基质、光照等方面进行了系统研究,以期为七叶一枝花的繁殖及栽培提供技术参考,解决供求矛盾。
1 材料与方法
1.1 试验材料
供试种子采自井冈山区野生七叶一枝花。
1.2 研究方法
1.2.1 种子吸水
随机选取100粒饱满种子,用蒸馏水清洗干净,吸干表水后用0.000 1 g电子天平称重。然后放在20℃蒸馏水中浸泡,第一天每隔2 h称重1次,第二天每隔6 h称重1次,计算平均值。
1.2.2 萌发过程观察
在种子萌发的不同阶段用解剖镜观察其形态变化。
1.2.3 变温层积与激素处理
种子采回后,去外果皮,用蒸馏水清洗干净后凉干,用1 000倍多菌灵液浸泡消毒半小时,然后用细沙分层贮藏,在人工气候箱内进行“冷→热→冷”变温处理,冷处理温度为5℃,热处理为20℃。设置3种变温处理,即“60 d冷→30 d热→60 d冷”(T1)、“60 d冷→30 d热→30 d冷”(T2)和“30 d冷→30 d热→30 d冷”(T3),置于人工气候箱内在20 ℃、周期性光照(1 500 lx,12 h·d-1)和以3 层湿润滤纸为基质的培养条件下进行种子萌发试验;选取最佳的层积处理,采用不同浓度的GA3浸泡处理24 h,用蒸馏水作对照,进行萌发试验,每隔5 d计算一次发芽数。每处理为30粒种子,3次重复。
1.2.4 温度、基质与光照处理
经过变温层积和GA3处理的种子,分别放在10℃、20℃、30℃条件下催芽试验;在最合适的萌发温度下设置珍珠岩、细砂、细黄土和滤纸4种基质进行萌发试验;选择最合适的温度和基质进行光照(1 500 lx,12 h·d-1)与黑暗对照试验。每处理为30粒种子,3次重复。
1.3 数据统计与分析
种子着床后每天观察正常萌发数,直至不再出现萌发种子为止,试验过程每天补充蒸馏水以保持滤纸湿润,以胚根从种孔伸出2 mm视为萌发[5],计算种子萌发率。采用SPSS19.0和Excel2007对数据进行统计分析与比较。
2 结果与分析
2.1 种子吸水特性
从图1可以看出,前4 h内为七叶一枝花种子的急剧吸水阶段,此阶段是由于细胞内的亲水物质进行吸胀吸水;4 h之后是停止吸水阶段,细胞利用已吸收的水分进行代谢作用[6]。研究发现种子浸泡48 h,重量从 4.75 g 增加到 5.66 g,增加了19.16%,说明种子不存在种皮吸水障碍。虽然七叶一枝花种皮较硬,但种子具有种脐和种孔,并不影响种子对水分的吸收,说明种皮不是影响胚发育的原因,影响种子萌发的因素可能是种子内源激素对胚发育的抑制而造成胚的生理后熟与形态后熟[7]。
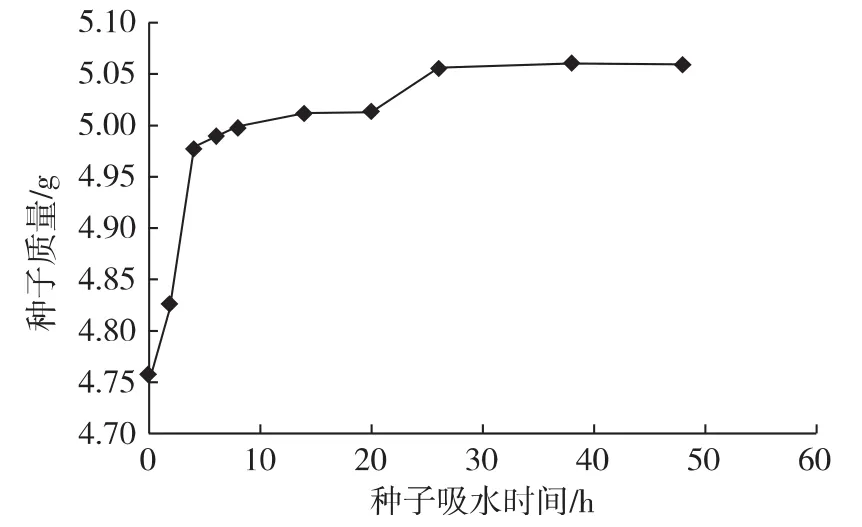
图1 七叶一枝花种子吸水曲线
2.2 种子萌发过程的观察
七叶一枝花种子萌发过程(如图2)大致可分为4个过程,即吸胀吸水、胚根伸出、胚根伸长、子叶出土。七叶一枝花种子在适宜的生长环境中,当胚生长发育到一定程度,胚根突破种皮,从胚珠珠孔伸出,当胚根长至1 cm左右时,种子基部开始膨大,形成一个小突起,胚根继续生长,但胚芽并不继续伸长,而是在种子内休眠,此为二次休眠期。当胚芽抑制物被解除,二次休眠打破后,胚芽将种壳顶出土面,子叶出土后连同种壳脱落,种子萌发完成。
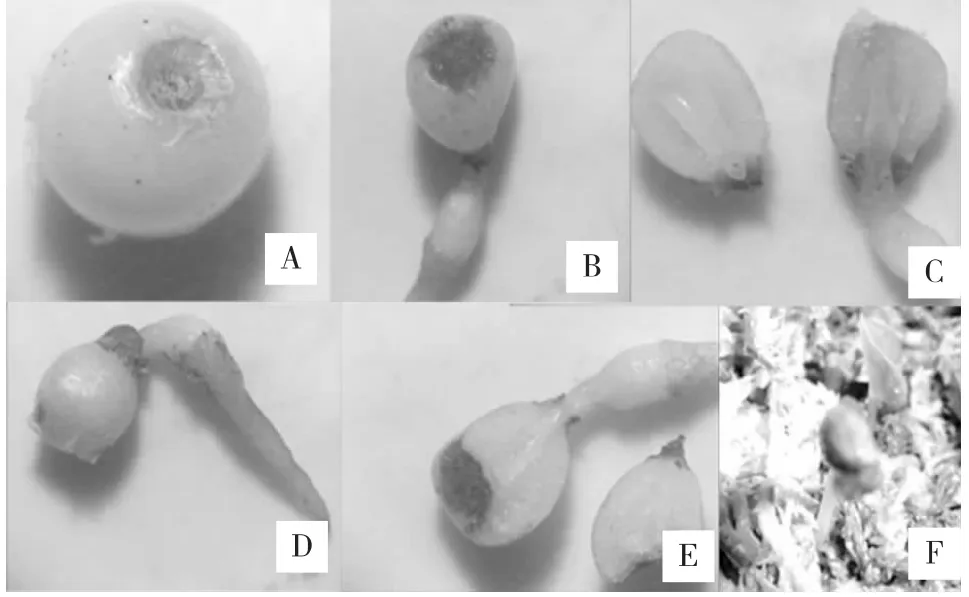
图2 七叶一枝花种子萌发过程
2.3 变温层积处理对种子萌发的影响
不同变温层积处理方式对种子的萌发有不同的影响,其最佳处理方式为“60 d(5℃)→30 d(20℃)→60 d(5℃)”,即T1。T1处理在57 d种子就开始萌发,萌发进程仅需28 d,萌发率高达73.3%,种子萌发较整齐,长势良好。从表1结果看,随着低温处理时间的延长,发芽率有所提高,但并没有达到显著性差异。但是随着低温处理时间的延长,七叶一枝花种子萌发时间提前,萌发过程明显缩短,T1处理萌发过程仅需28 d,T2为35 d,T3为46 d,差异达显著水平(p<0.05)。试验还发现无论冷处理时间长短,种胚都要经过近两个月以上的生理后熟和形态后熟的过程才能萌发。
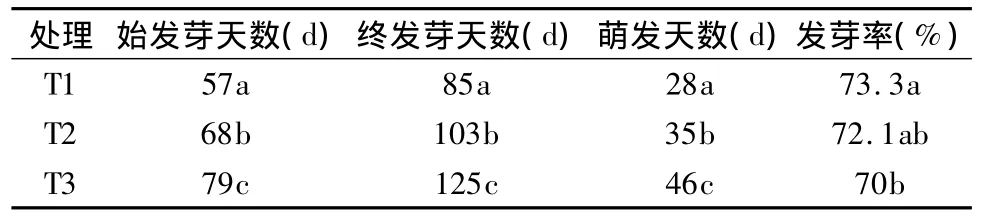
表1 不同层积处理对种子萌发的影响
2.4 不同浓度GA3处理对种子萌发的影响
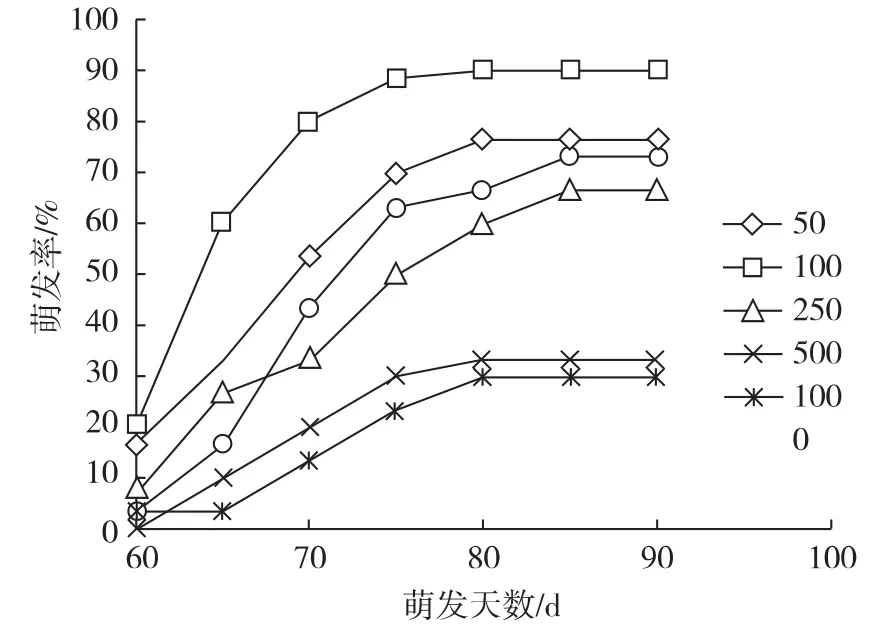
图3 不同浓度的GA3处理下七叶一枝花种子逐日萌发曲线
将经过变温层积T1处理的种子采用不同浓度的GA3浸泡处理24 h,用清水作对照,种子萌发率见图2和表2。由图3可知,七叶一枝花种子在不同浓度的赤霉素下均能萌发,萌发率在100 mg·L-1以下时随着浓度的增加而增加,100 mg·L-1以上时随浓度的增加而降低。说明低浓度的GA3对七叶一枝花种子萌发有促进作用,当浓度过高时则对其萌发产生抑制作用。
由表2可知,经50~100 mg·L-1GA3处理的种子,其萌发率高于对照,其中100 mg·L-1GA3处理的种子萌发率高达90%,高出对照16.7%,差异显著(P <0.05);经250 ~1 000 mg·L-1GA3处理的种子,其萌发率则显著(p<0.05)低于对照,其中500 mg·L-1以上的GA3处理的种子萌发率仅为30%左右。不同处理的七叶一枝花种子开始萌发时间也不同,100 mg·L-1GA3处理组在第52 d开始萌发,比对照组提前 5d,差异显著;50 mg·L-1、250 mg·L-1、1 000 mg·L-1处理组分别在第 54 d、56 d、60 d 开始萌发,与对照组差异不显著;500 mg·L-1处理组在第63 d开始萌发,比对照推迟了5 d,差异显著。
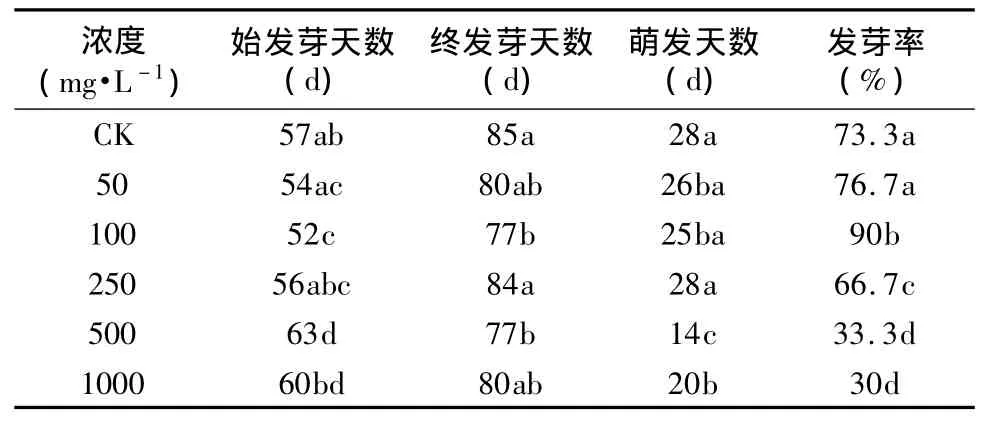
表2 不同浓度GA3对七叶一枝花种子萌发的影响
2.5 不同温度对种子萌发的影响
将经过 T1层积和100 mg·L-1GA3处理的种子,分别放在10℃、20℃、30℃条件下进行催芽试验,结果如表3所示。七叶一枝花种子在10℃无萌发现象,在20℃时发芽率高达90%,30℃发芽率只有13.3%,各温度处理间萌发率的差异均达到了显著水平(p<0.05),说明七叶一枝花种子萌发对温度要求严格。从以上结果看,七叶一枝花种子萌发最适宜温度为20℃,萌发温度范围较窄,过高或过低对其萌发均不利。
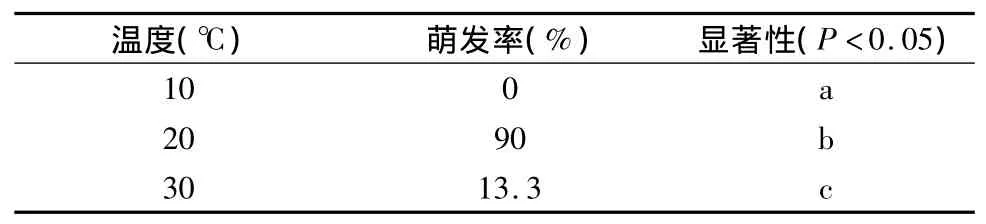
表3 不同温度对七叶一枝花种子萌发的影响
2.6 不同基质对种子萌发的影响
不同基质发芽试验结果表明:细沙与其他3种基质的发芽率存在显著差异(p<0.05),其萌发率最高为96%;珍珠岩、细黄土和滤纸3种基质萌发率分别为85%、86%和90%,差异不显著。试验表明,七叶一枝花种子萌发以细沙基质为佳。
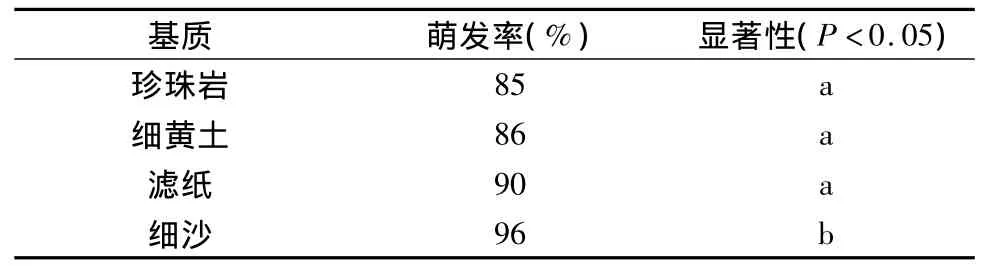
表4 不同基质对七叶一枝花种子萌发的影响
2.7 光照对种子萌发的影响
采用1 500 lx每天12 h光照和黑暗对比试验,结果如表5所示。光照条件与黑暗条件下七叶一枝花种子萌发率分别为96%和93%,无显著性差异,说明七叶一枝花种子萌发不受光照的影响。
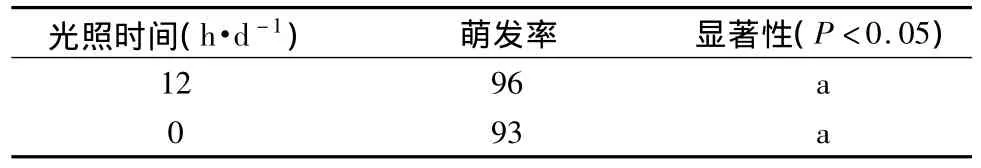
表5 光照对七叶一枝花种子萌发的影响
3 讨论
3.1 变温层积处理与激素对种子萌发的影响
休眠是植物经过长期演化过程对环境与季节变化的适应,就本身而言,它既是一个重要的生命过程,又是一个重要的生物学特性。由于七叶一枝花花期较短、果期较长,大量种子不能完全成熟[8],有些果实虽然成熟了,但由于其生长在海拔相对较高的山区,为了适应较为寒冷季节低温的到来,种胚停止了发育,以度过不良的生长环境。本研究也发现七叶一枝花种子具有形态与生理深度休眠的特性,种子播种前采用变温处理与激素处理相结合,可以在较短的时间内打破休眠,促使种子萌发,比自然条件下提前10个月至1年出苗。
变温层积处理可以打破植物休眠、促进胚的发育和种子萌发已经在滇重楼等其他重楼属植物种子上取得了较好的效果,陈疏影认为种子休眠的真正原因,可能是种子内源激素含量较高而抑制种胚的发育,通过一段时间的低温处理,抑制物被解除,促使细胞分裂和伸长的激素合成,胚才能生长[9]。本研究也发现,变温层积可提前结束七叶一枝花种子的休眠期,促进其萌发,且变温层积的关键在低温处理时间长短与温度的高低。陈疏影等认为较长的低温环境是滇重楼种子休眠的重要原因,而在20℃环境下滇重楼种子萌发状况良好,他建议提高变温层积的温度以打破休眠,缩短种子后熟期[9],而本研究则发现随低温(5℃)处理时间的相对延长,七叶一枝花种子发芽率提高,本研究从侧面证实了徐文娟关于滇重楼种子的研究结论,她认为低温处理可促进种子内GA3的合成[10]。另外本研究还发现,无论何种处理条件下,种胚必须经过2个月左右的生理后熟期才能萌发,其原因有待进一步深入研究。
GA3可软化种胚周围的组织,促进胚的生长,是解除种子休眠的关键内源信号分子[11]。孟繁蕴等研究了滇重楼种子发育过程中内源激素的变化,发现GA3可以将胚乳的贮藏物质转化为可供胚细胞分裂与伸长的物质,其含量的增加与其种胚发育的速度是一致,他认为GA3可诱导或抑制某些mRNA和蛋白质的合成,从而促进胚乳中酶的合成,提高酶活性,加速物质的合成与分解,打破种子休眠,促进胚的发育[12]。本研究表明GA3可促进七叶一枝花种子萌发,最佳浓度为100 mg·L-1,但并不是所有浓度对其萌发都有促进作用,当浓度过高(超过250 mg·L-1)则对其萌发有不同程度的抑制作用,这与马红媛等关于GA3对羊草种子萌发影响的研究结果类似[13]。
3.2 温度、基质与光照对种子萌发的影响
温度是七叶一枝花种子萌发的重要因素,对其萌发影响达极显著水平(p<0.01)。七叶一枝花种子萌发在20℃条件下其萌发率可达90%,在30℃条件下其萌发率显著降低,仅为13%,10℃条件下种子不萌发,由此可见七叶一枝花种子萌发对温度适应范围狭窄。七叶一枝花种子在10月成熟[8],此时已是深秋,温度较低,不适合种子萌发,另外低温可促进种子内GA3的积累[10],使种子完成形态与生理的后熟,等来年春天气候适宜时萌发,避开不良的生长环境,这是植物对环境适应性的表现。
陈志欣等以“勐海大叶茶”种子为材料,研究不同基质对其萌发的影响,在不同的基质中,‘勐海大叶茶’种子萌发率的变化不大[14]。本研究也得出相似的结论,七叶一枝花在珍珠岩、细黄土、滤纸、细沙中种子的萌发率依次升高,但差异并不显著。这可能是因为七叶一枝花属单子叶植物,其种子有胚乳,胚乳中丰富的营养物质完全能够支持种子萌发,所以基质对其影响不大。
光照对不同植物种子萌发的影响不同,郑光华认为光照可促进大部分植物种子的萌发,抑制少部分植物种子萌发,尚有极少植物种子的萌发不受光照的影响[15]。本研究发现,七叶一枝花种子萌发对光照的要求很低,在光照与黑暗条件下,其萌发率均能达到90%以上,但在光照条件下,其萌发率较黑暗整齐且胚轴生长状况也比较好,与沈素贞等关于光照对短梗大参种子萌发的影响结论一致[16],其原因可能是光照可促使胚轴快速合成叶绿素,从而进行光合作用,为其生长发育提供少量的能量。
4 结论
(1)七叶一枝花种子不存在吸水障碍,解剖镜观察其萌发过程发现种子具有二次休眠的特性。
(2)变温层积处理过程中,随着低温处理时间的相对延长,萌发率有所提高。无论冷处理时间长短,种胚都要经过近两个月以上的生理后熟和形态后熟的过程才能萌发。
(3)低浓度的赤霉素可显著促进七叶一枝花种子的萌发,高浓度则抑制其萌发,100 mg·L-1的赤霉素处理种子时萌发率最高。
(4)七叶一枝花萌发温度范围较窄,20℃为其萌发的最佳温度,低于10℃不萌发;基质与光照对其萌发影响不大。
[1] 国家药典委员会.中华人民共和国药典:一部[M].北京:化学工业出版社,2005.
[2] 国家中医药管理局《中华本草》编委会.中华本草精选本:下册[M].上海:上海科技出版社,1998:2062-2063.
[3] 袁宏伟.中药重楼药理作用及临床应用进展[J].中国社区医师,2007,12:21 -23.
[4] 天地.中药重楼和山豆根的后市走向[J].农家致富顾问,2009,4:12 -13.
[5] 陈志欣,郑丽,兰芹英,等.‘勐海大叶茶’种子萌发特性[J].植物分类与资源学报,2012,34(1):63 -68.
[6] 潘瑞炽.植物生理学[M].北京:高等教育出版社,2008.
[7] 王英,饶军,王海辉,等.七叶一枝花人工栽培技术研究[J].卫生职业教育,2011(23):116-117.
[8] 田启建,陈功锡,刘冰,等.人工栽培七叶一枝花的生物学特征及物候期研究[J].湖南农业科学,2010(13):18-20.
[9] 陈疏影,尹品训,杨艳琼,等.变温层积对解除滇重楼种子休眠及内源激素变化的研究[J].中草药,2011,(4):793 -795.
[10] 徐文娟,李先恩,孙鹏,等.滇重楼种子层积后脱落酸和赤霉素相关基因表达水平的研究[J].中草药,2013,44(3):338-342.
[11] Yamaguchi S,Kamiya Y.Gibberellins and light stimulated seed germination[J].Journal of Plant Growth Regulation,2001,20(4):369-376.
[12] 孟繁蕴,汪丽娅,张文生,等.滇重楼种胚休眠和发育过程中内源激素变化的研究[J].中医药学报,2006,4:334 -337.
[13] 马红媛,梁正伟,黄立华,等.4种外源激素处理对羊草种子萌发和幼苗生长的影响[J].干旱地区农业研究,2008,26(2):69-73.
[14] 陈志欣,郑丽,兰芹英,等.‘勐海大叶茶’种子萌发特性[J].植物分类与资源学报,2012,34(1):63-68.
[15] 郑光华.种子生理研究[M].北京:科学出版社,2004.
[16] 沈素贞,梁文斌,宋雷.野生观赏植物短梗大参种子萌发特性的研究[J].中南林业科技大学学报,2013,33(8):53-58.
猜你喜欢
中国机械工程(2022年8期)2022-05-09
中国土壤与肥料(2021年5期)2021-12-12
中国土壤与肥料(2021年5期)2021-12-02
儿童时代·幸福宝宝(2021年1期)2021-03-29
红楼梦学刊(2020年2期)2020-02-06
北京园林(2020年4期)2020-01-18
小资CHIC!ELEGANCE(2019年40期)2019-12-10
当代陕西(2019年6期)2019-04-17
现代园艺(2019年3期)2019-03-08
中成药(2017年7期)2017-11-22
