
陕北黄土区山杏林下草本层植物群落特征研究
2013-09-27 11:23王露露朱清科赵彦敏郑学良
水土保持通报 2013年1期
王露露,朱清科,赵彦敏,郑学良,李 萍
(北京林业大学 水土保持与荒漠化防治教育部重点实验室,北京100083)
山杏(Armeniaca sibiriea)喜光,耐干旱、耐寒、耐瘠薄。自20世纪50年代以来,半干旱黄土区山杏人工造林面积不断扩大,一是促进生态恢复,二是生产甜杏仁;且大部分无或者有较小人为干扰,群落自然演替。自90年代尤其是退耕还林政策实施以来,山杏研究逐步受到重视,但是大多集中在山杏的丰产技术、栽培技术、抗旱生理特征方面[1-2],而对不同立地类型下山杏人工林的生态系统健康状况和生态恢复效果等未有专题报道。陕北黄土区在植被区划上属森林草原交错带,植被属性复杂,由于原生植被基本被破坏殆尽,在该地区的生态修复工作中主要以人工乔(灌)林和自然恢复草地的生态系统健康状况和生态恢复效果为参考来确定植被恢复的目标和方法[3]。因此,对这一地区分布广泛、种植面积大的山杏人工林的群落结构、生态系统功能、演替过程等方面进行研究,评价其生态恢复效果,对该区的植被恢复和重建具有重要的指导意义。
在森林生态系统中林下草本层是一个重要组成部分,特别是在人工林中,由于建群种种类单一,群落垂直结构简单,群落的物种多样性主要体现在林下草本层,另外林下植被在促进系统养分循环、减少水土流失和维护林地土壤质量中也起着不可忽视的作用[4];林下草本层与生态系统各组分关系密切,其物种种类和生态型组成、多样性、生物量等群落特征随上层林木发育过程不断发生改变,也会随着生境条件的变化而变化[5],因此人工林下草本层植物群落特征和演替方向是评价生态系统健康及生态恢复效果的重要依据。
以往对该区人工林的研究多集中在林分的径级和高度分布、生产力[6],林内土壤水分、养分状况上[7],对林下植物群落的研究则不多,主要为同龄不同密度条件下或同龄不同树种林下物种组成、多样性的差异[8-10],对不同立地类型人工林林下植物演替进程和群落特征鲜有研究。本文以位于半干旱黄土区的陕西省延安市吴起县为研究区,吴起县的造林历史可以追溯到新中国成立之初,但是到1997年之前林木保存率很低,1998年吴起县先于全国开始了大规模“退耕还林(草)”,是退耕还林(草)工程建设试验示范县之一,截至2005年年底,累计退耕还林1 132.47km2。本文选择该县不同立地类型保存下来的70—80年代营造的山杏人工纯林和山杏沙棘混交林的老龄林为研究对象,通过对林下草本层物种组成、多样性、水分生态型、盖度和生物量分布等群落特征动态的研究,揭示陕北半干旱黄土区各立地类型下山杏人工林的林下草本层植物演替过程和方向及不同植被配置模式对林下草本层发育的长期影响,从而从一个新的视角探讨各立地类型下营造乔木林的合理性,为该地区生态恢复中植被配置模式的选择提供依据。
1 研究区概况
吴起县地处107°38′57″—108°32′49″E,36°33′33″—37°24′27″N,海拔1 233~1 809m,面积3 791.5km2,属黄土丘陵沟壑区。多年平均降水量478.3mm,年际变化大、季节分配不均,50%~80%降雨量集中在7—9月,年平均气温7~8℃,极端最高气温37.1℃,极端最低气温-25.1℃,无霜期96~146d,为暖温带大陆性干旱季风气候。多年平均陆地蒸发量400~450mm,属干旱、半干旱地区。土壤类型为黄绵土,质地为轻壤。
该县植物组成以华北区系植物占主导地位,植被为森林草原向草原过渡类型。由于长期的人类经济活动,原有植被已不复存在,在现状植被中以次生植被为主,落叶阔叶林及灌木草丛占主导地位。主要树种有山杏、刺槐(Robinia pseudoacacia)、小叶杨(Populus simonii)、油松(Pinus tabulaeformis)、侧柏(Platycladus orientalis)、山桃(Amygdalus daridiana)等,灌木主要是柠条(Caragana korshinskii)、沙棘(Hippophae rhamnoides)。
2 研究方法
2.1 样地选择与群落调查
对吴起县城周边的杨青川流域、金佛坪流域、袁沟流域、柴沟流域进行全面踏查,了解山杏老龄林的树种组成、龄级、立地类型等。立地类型的划分主要根据地形因子中的坡度和坡向,不考虑海拔、土壤等因素的影响,因此所选样地土壤类型相同,均为黄绵土,样地间高差在100m以内。在黄土丘陵沟壑区,以10°~35°的坡面分布面积最大[11],这类坡面也是营造人工林的重点,因此本研究选择在这一坡度区间内划分立地类型,10°~25°划为缓坡,25°~35°划为陡坡;坡向划分为阳坡和阴坡。在此基础上采用空间代替时间的方法,在以上4个小流域选择不同立地类型的25龄、40龄山杏人工纯林和40龄山杏沙棘人工混交林典型样地。共有样地类型11种(阳向陡坡立地下没有山杏—沙棘混交林),每种类型取3个样地,共有样地数33个。各样地概况如表1所示。

表1 山杏人工林样地基本情况
每个样地面积10m×10m,记录样地的位置、海拔、坡度、坡向,对样地内山杏打生长锥测定林分年龄。在每个样地的四角及中央布设1m×1m的草本层调查样方,记录草本样方中植物种类及每个种的个体数、高度和盖度,对样方内植物地上部分全部收获称量鲜重,并在每个山杏沙棘混交林样地的四角及中央布设2m×2m的灌木样方调查沙棘密度。
2.2 物种多样性测度
采用重要值测度群落种群组成,选取丰富度指数(S),Shannon—Wiener指数(ISW)衡量植物群落物种多样性特征,Pielou均匀度指数(J)衡量植物群落物种的分布均匀程度[12]:
丰富度指数S:
S=群落中出现的物种数目
Shannon—Wiener指数ISW:
ISW=-∑(Pi×lnPi);
Pielou均匀度指数J:
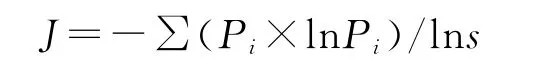
式中:S——物种种数;Pi=Ni/N,i=1,2…S;Ni——样地中第i种物种的重要值,并且N=∑Ni。
重要值的计算公式:

2.3 植物水分生态型
依照植物与水分的关系,可以将植物分为旱生植物、中生植物和水生植物等水分生态类型。黄土高原干旱缺水,其显域生境中生活着从旱生到中生各种水分生态类型的植物。本文将山杏林下植物分为4种水分生态型:旱生型,如冰草(Agropyron cristatum)、星毛委陵菜(Potentilla acaulis)、长芒草(Stipa bungeana)、甘草(Glycyrrhiza uralensis)、糙隐子草(Cleistogenes squarrosa)、硬质早熟禾(Poa sphondylodes)、火绒草(Leontopodium leontopodioides)、二裂 委 陵 菜 (Potentilla bifurca)、冷 蒿 (Artemisia frigida)、糙叶黄耆(Astragalus scaberrimus)等;旱中生(中旱生)型,如铁杆蒿(Artemisia sacrorum)、茭蒿(Artemisia giraldii)、达乌里胡枝子(Lespedeza davurica)、大披针苔草(Carex lanceolata)、阿尔泰狗娃花(Heteropappus altaicus)、中华隐子草(Cleistogenes chinensis)、白羊草(Bothriochloa ischaemum)、菊叶委陵菜(Potentilla tanacetifolia)、茵陈蒿(Artemisia capillaris)、野菊花(Dendranthema lavandulifolium)、狭叶柴胡(Bupleurum scorzonerifolium)、披针叶黄华(Thermopsis lanceolala);中生型,如龙须菜(Asparagus schoberioides)、风毛菊(Saussurea japonica)、蓬子菜(Galium verum)、茜草(Rubia cordifolia);湿生型,如芦苇(Phragmites australis)[13-14]。
3 结果与分析
3.1 林下草本层物种组成
山杏林下草本层共有维管束植物58种,分属19科,41属。其中含有5个种以上的科有菊科(Compostae,14种),其次是禾本科(Gramineae,11种)和豆科(Leguminosae,8种),占总科数的15.8%,这3科包含的种数有33个,占总种数的56.9%。其余的16科中有9科为单属单种占总科数的47.4%。含有1个种以上的属11个占总属数的26.8%,11属共包含28种占总种数的48.3%,其中包含种数较多的有菊科蒿属(Artemisia,4种),豆科黄耆属(Astragalus,3种),菊科菊属(Chrysanthemum,3种),蔷薇科陵菜属(Potentilla,3种),禾本科隐子草属(Cleistogenes,3种)。其余30个属均为单种属,占总属数的73.2%。物种组成分析结果表明,菊科、禾本科、豆科3科在半干旱黄土区人工山杏林林下草本层中占有重要地位,物种区系构成表现为多数种属于少数科,少数种属于多数科,且很多物种为单属种,符合西北干旱区的植物区系特征[15]。
表2表明,阴向缓坡、阴向陡坡、阳向缓坡25龄山杏纯林林下均为旱中生小半灌木铁杆蒿、达乌里胡枝子组成的共优群落。主要伴生种一般有阿尔泰狗娃花、大披针苔草、中华隐子草、火绒草、长芒草等,仅在不同立地类型下重要值大小不同。随着自然演替的进行,林下草本层中大披针苔草的优势度不断增加,在40龄的山杏纯林下成为优势种,同时铁杆蒿仍在群落中占有重要地位。
以上3种立地类型下的40龄山杏沙棘混交林林下草本层群落类型均与同龄山杏纯林相似,为铁杆蒿与大披针苔草的共优群落。该地区自然恢复草地在阴坡和阳坡建群种一般不同,阳坡物种更趋向于旱生[16];但各龄级山杏林下草本层优势种却基本相同,可能由于山杏林的生长一定程度上减小了生境间尤其是不同坡向立地间的差异。

表2 不同立地类型不同植被配置模式山杏林林下草本层物种组成
阳坡陡坡的25龄及40龄山杏纯林林下则分别为星毛委陵菜+铁杆蒿群落和甘草+大披针苔草群落。星毛委陵菜、甘草在半干旱黄土区多出现在山地草原或森林草原干旱生境中,均为典型旱生植被[14]。因此,阳坡陡坡山杏纯林下出现这类次生植被,可能是由于生境逐渐在旱化。
3.2 林下草本层物种多样性与生物量
由表3可见,相同立地类型下40龄山杏纯林林下草本层的香农指数、丰富度指数和均匀度指数均低于25龄山杏纯林。林木进入成熟期以后,林下环境因子趋于稳定,因此优势种的优势度持续增加,导致多样性降低。另外,多样性各指数的方差分析结果表明,除阳坡陡坡立地40龄山杏纯林林下物种多样性显著低于25龄林(p<0.01),其他立地类型下这2个龄级山杏林林下物种多样性水平的差异并不显著,说明在这一阶段林下物种多样性变化缓慢。40龄山杏沙棘混交林林下物种多样性高于同龄山杏纯林,但不显著。
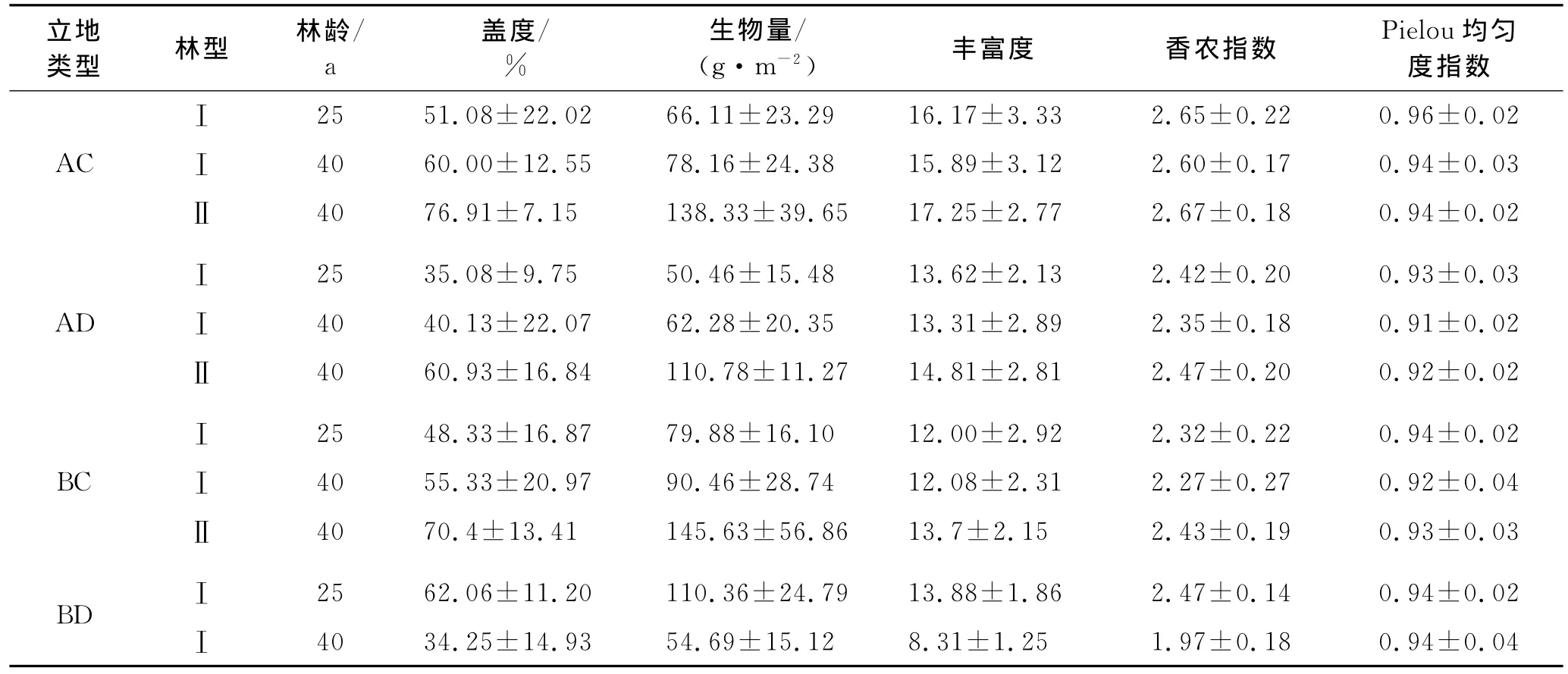
表3 不同立地类型不同植被配置模式山杏林林下草本层物种多样性、盖度和生物量
山杏纯林林下草本层盖度与生物量的变化规律基本一致,在阴坡缓坡、阴坡陡坡、阳坡缓坡3种立地类型下均为40龄山杏纯林高于25龄山杏纯林,但不显著;而在阳坡陡坡40龄山杏纯林则显著低于25龄山杏纯林(p<0.01)。40龄山杏沙棘混交林林下草本层盖度与生物量均高于同龄山杏纯林,且生物量的差异极显著(p<0.01),盖度的差异也达到了显著水平(p<0.05)。
同林型山杏林在不同立地类型下林下物种多样性高低排序为阴向缓坡>阴向陡坡>阳向缓坡>阳向陡坡,其中阴向缓坡林下物种多样性与其他3种立地的差异显著(p<0.05)。40龄山杏纯林及混交林在各立地类型下林下草本层生物量大小顺序为阳向缓坡>阴向缓坡>阴向陡坡>阳向陡坡,25龄山杏纯林则是在阳向陡坡立地下生物量最大。林下草本层盖度大小顺序与生物量基本一致,也为缓坡大于陡坡。林下草本层盖度及生物量的变化与乔木层郁闭度有密切的关系[17],郁闭度较高的林分其林下光照条件较差,并且种内种间竞争较大,不利于林下植被的生长发育。在缓坡条件下阴坡林分的郁闭度相对于同龄阳坡林分要大,因此林下植被生物量相对减小一些;陡坡不利于植物种子的定居,水、养分条件相对缓坡也较差,因此盖度和生物量也相对较小。阳坡陡坡的山杏纯林林下草本层生物量25龄时在4种立地类型中最大而40龄时最小,可能的原因是在该立地下山杏林乔木层生长差,郁闭度低,在25龄林林下发育了大量的旱生、喜阳植物,但乔木的生长对地力的消耗超出了其承载能力,使生境无法得到改善甚至恶化,表现在林下植被发育上为随着林龄进一步增大,植被盖度、生物量、多样性均急剧下降。
3.3 林下草本层植物水分生态型
植物由于外界生态因素的影响,逐渐演化出各种各样的形态和结构以适应生长的环境,其中影响最大的是植物生长环境中水分的供应状况。黄土高原干旱缺水,其显域生境中生活着从旱生到中生各种水分生态类型的植物。
图1表明,所有4种立地类型下的山杏25龄、40龄人工纯林及混交林林下草本层均为以铁杆蒿、大披针苔草、达乌里胡枝子、茭蒿、阿尔泰狗娃花等为代表的旱中生(中旱生)植物重要值所占比例最大,取值均在53%~61%,其次是旱生植物,如冰草、长芒草、糙隐子草等,重要值百分比为19%~34%,中生和湿生植物比例最低,分别为5%~20%和0%~5%。分别对比相同立地类型下25龄与40龄山杏纯林林下草本层植物水分生态型分布比例可以看出,在阴向缓坡、阴向陡坡、阳向缓坡均为随着林龄的增加旱生植物比例减小、旱中生和中生植物比例增加,说明林下草本层逐渐由旱生向中生转变;在阳向陡坡则是随着林龄增加旱生植物如甘草、星毛委陵菜的重要值百分比增加,而中生植物优势度减小,林下草本层逐渐趋向于旱生。另外,40龄山杏沙棘混交林林下旱生植物所占比例均小于同龄山杏纯林,中生加湿生植物比例也均高于山杏纯林。山杏纯林林下草本层植物各种水分生态型在不同立地类型下重要值所占比例按大小排序为:旱生植物为阴向缓坡<阴向陡坡<阳向缓坡<阳向陡坡,中生加湿生植物则为阴向缓坡>阴向陡坡>阳向缓坡>阳向陡坡。
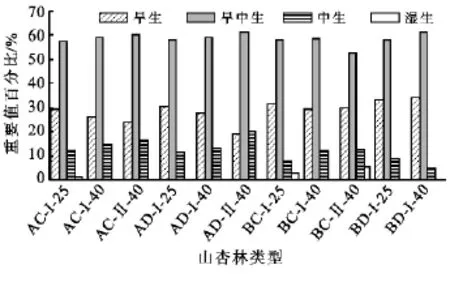
图1 不同立地类型与不同植被配置模式山杏林林下草本层植物水分生态型
4 结论与讨论
陕北黄土区山杏林下草本层共出现维管束植物19科41属58种,以菊科、豆科、禾本科为主,有典型西北干旱区的植物区系特征。山杏纯林进入成熟期后,草本层演替趋势在阴向缓坡、阴向陡坡、阳向缓坡3种立地类型下基本相同,而在阳向陡坡则相反。阴向缓坡、阴向陡坡、阳向缓坡草本层由铁杆蒿、达乌里胡枝子组成的多年生蒿类群落演替到大披针苔草或铁杆蒿与大披针苔草组成的森林苔草群落。对比该地区撂荒地(退耕地)由以铁杆蒿、茭蒿等为优势种的多年生蒿类群落到较稳定的地带性白羊草草原群落[18]的自然演替过程,说明营造山杏乔木林后产生了林下小环境,草本层没有向地带性的草原植被群落发展而是具有更明显的森林群落特征。并且在造林25~30龄后,随着林龄的增加,草本层群落整体水分生态型由旱生型向中生型转变、盖度和生物量小幅升高、多样性略有降低也均符合群落正向演替并趋于稳定的一般规律,说明在以上几种立地类型下,山杏人工林发挥了森林生态系统的功能,对生态环境的改善有促进作用。而在阳坡陡坡,山杏纯林林下草本层物种组成由星毛委陵菜—铁杆蒿群落演替到甘草—大披针苔草群落,典型旱生物种始终占据优势,而且多样性、盖度、生物量均随林龄增加显著降低等现象均说明群落正在逆行演替,生境不断恶化。
另外,在阴向缓坡、阴向陡坡及阳向缓坡,与纯林相比山杏沙棘混交林下草本层优势种虽然相同,但物种多样性、植被盖度和生物量均增加,其中盖度和生物量显著增大,并且旱生物种优势度减少,中生成分增加。在半干旱的黄土区,能够维持较高生态系统稳定性的能力和良好的水土保持功能是选择人工林植被配置模式的重要依据。稳定性与物种多样性关系密切,虽然多年来对于有关“多样性与稳定性”的生态学基本问题争论不断,但大多数试验研究结论和多种假说认为多样性导致群落和生态系统水平上的稳定性[19];高的物种多样性可能意味着更多种功能群或者冗余作用的存在,能够对于干扰作用下生态系统功能的发挥起到补偿作用,从而提高群落和生态系统的稳定性[20]。人工林物种多样性主要体现在林下植被层,因此增加林下植被层物种多样性对提高生态系统稳定性有着一定的意义。另外,人工林群落中地被植物层的物种多样性、盖度、生物量等的提高,能够直接减弱降雨动能,同时通过改善表层土壤结构,增加入渗和土壤抗蚀性,从而减少地表径流和土壤侵蚀量[21],具有更好的水土保持效果。因此从林下植被发育情况的角度来看,山杏沙棘混交林比山杏纯林具有更高的稳定性和水土保持功能。
近年来,半干旱黄土高原森林草原区人工林下普遍出现不同程度的土壤干层,林木出现生长发育不良现象,部分形成“小老树”,甚至成片死亡,引发了对在这一地区配置乔木林的质疑[22]。然而研究表明,这一地区能够自然更新的天然针叶疏林同样生长缓慢并且林下存在土壤干层的发育[23]。说明森林在半干旱地区生长缓慢和产生一定程度的土壤干化也是自然现象[24],抗旱性强的树种在受到干旱胁迫后会通过减慢生长速度来减少蒸腾耗水同时提高水分利用效率,从而保持与环境供水能力的平衡状态,维持群落稳定性;只有当树种选择失当、密度过大、群落生产力过高导致林下土壤处于持续而严重的“赤字”状态,才会使植物生长衰退,群落发生逆向演替。因此本研究认为在半干旱陕北黄土区恢复植被应根据立地条件选择树种和控制密度。在阴向缓坡、阴向陡坡、阳向缓坡立地应配置山杏这类低耗水、生长慢乔木树种,采用与沙棘等乡土灌木树种混交的模式更有利于提高群落稳定性和加快生态恢复的进程,但应严格控制密度,山杏林林分稳定密度约为400~800株/hm2,且山杏沙棘混交林密度比纯林更小。不同立地类型下准确的林分合理初植密度和稳定密度还需要更进一步研究确定。在阳向陡坡立地应以营造生长速度适中的灌木林或自然恢复草地植被群落为宜。
[1]刘明国,王威,贺江,等.山杏混交林花果期小气候特点及其对坐果率的影响[J].东北林业大学学报,2010,38(6):28-30.
[2]黄世臣,李熙英.水分胁迫条件下接种菌根菌对山杏实生苗抗旱性的影响[J].东北林业大学学报,2007,35(1):31-32.
[3]刘中奇,朱清科,秦伟,等.半干旱黄土区自然恢复与人工造林恢复植被群落对比研究[J].生态环境学报,2010,19(4):857-863.
[4]褚建民,卢琦,崔向慧,等.人工林林下植被多样性研究进展[J].世界林业研究,2007,20(3):9-13.
[5]Kume A,Satomura T,Tsubei N,et al.Effects of understory vegetation on the ecophysiological characteristics of an overstory pine,Pinus densiflora [J].Forest Ecology and Management,2003,176:195-203.
[6]张光灿,刘霞,周泽福,等.黄土丘陵区油松水土保持林生长过程与直径结构[J].应用生态学报,2007,18(4):728-734.
[7]张建军,李慧敏,徐佳佳.黄土高原水土保持林对土壤水分的影响[J].生态学报,2011,31(23):7056-7066.
[8]赵娜,查同刚,周志勇.晋西黄土区不同树种配置对林下植被物种多样性的影响[J].东北林业大学学报,2011,39(3):44-45.
[9]杨晓毅,李凯荣,李苗,等.陕西省淳化县人工刺槐林林分结构及林下植物多样性研究[J].水土保持通报,2011,31(3):194-201.
[10]高艳鹏,赵廷宁,骆汉.晋西黄土丘陵沟壑区人工林下草本植物生物多样性研究[J].水土保持通报,2011,31(1):103-108.
[11]赵荟,朱清科,秦伟,等.黄土高原沟壑区干旱阳坡的地域分异特征[J].地理科学进展,2010,29(3):327-334.
[12]马克平,黄建辉,于顺利,等.北京东灵山地区植物群落多样性的研究:Ⅱ.丰富度、均匀度和物种多样性指数[J].生态学报,1995,15(3):268-277.
[13]朱志诚,黄可.陕北黄土高原森林草原地带植被恢复演替初步研究[J].山西大学学报:自然科学版,1993,16(1):94-100.
[14]雷明德.陕西植被[M].北京:科学出版社,1999:233-282.
[15]党荣理,潘晓玲.西北干旱荒漠区种子植物科的区系分析[J].西北植物学报,2002,22(1):24-32.
[16]秦伟,朱清科,张宇清,等.陕北黄土区生态修复过程中植物群落物种多样性变化[J].应用生态学报,2009,20(2):403-409.
[17]Rankin W J,Tramer E J.Understory succession and the gap regeneration cycle in a Tsuga canadensis forest[J].Canadian Journal of Forest Research,2002,32(1):16-23.
[18]秦伟,朱清科,刘中奇,等.黄土丘陵沟壑区退耕地植被自然演替系列及其植物物种多样性特征[J].干旱区研究,2008,25(4):507-512.
[19]Tilman D,Wedin D,Knops J.Productivity and sustainability influenced by biodiversity in grassland ecosystems[J].Nature,1996,379:718-720.
[20]党承林.植物群落的冗余结构:对生态系统稳定性的一种解释[J].生态学报,1998,18(6):665-672.
[21]王震洪,段昌群,侯永平,等.植物多样性与生态系统土壤保持功能关系及其生态学意义[J].植物生态学报,2006,30(3):392-403.
[22]李军,陈兵,李小芳,等.黄土高原不同植被类型区人工林地深层土壤干燥化效应[J].生态学报,2008,28(4):1429-1445.
[23]朱志诚.陕北黄土高原杜松疏林草原初步研究[J].林业科学,1991,27(4):447-451.
[24]赵景波,周旗,侯甬坚.黄土高原土壤干层对生态环境建设的影响[J].陕西师范大学学报:自然科学版,2003,31(4):93-97.
猜你喜欢
现代农村科技(2022年11期)2022-12-16
汽车实用技术(2022年7期)2022-04-20
现代农村科技(2021年3期)2021-04-11
山东交通科技(2020年1期)2020-07-24
森林工程(2018年4期)2018-08-04
方圆(2018年23期)2018-01-07
辽宁林业科技(2017年4期)2017-06-22
甘肃林业(2016年4期)2016-11-07
广西林业科学(2016年4期)2016-03-16
湖南安全与防灾(2015年10期)2015-03-03
