
槟榔江水牛STAT 5A基因多态性及其与产奶性状的关联性研究
2013-09-22 08:58刘学洪余长林余选富刘定江史宪伟
中国牛业科学 2013年4期
季 敏,刘学洪*,余长林,余选富,刘定江,史宪伟
(1.云南农业大学动物科学技术学院,云南 昆明 650201;2.云南省腾冲县畜牧工作站,云南 腾冲 679100)
1 引言
槟榔江水牛是我国迄今为止发现的唯一的河流型水牛遗传资源,分布在云南省腾冲县及周边。槟榔江水牛有着独特的遗传特性,尤其是在产奶性能比较优秀。有研究结果显示槟榔江水牛平均产奶量是德宏水牛的2倍多,比摩拉水牛高15%,但个体间生产性能差异较大[1]。章纯熙等[2]对101头槟榔江水牛的105个泌乳期泌乳量结果进行测定,结果发现槟榔江水牛平均泌乳天数为270d,比其他河流型和沼泽型水牛平均泌乳天数短,而平均泌乳量为2 450kg,比摩拉和尼里拉菲等河流型水牛高8.4%~15.0%,比广东水牛和温州水牛等沼泽型水牛高124.4%~140.3%。另外通过分析腾冲水牛场提供的31头母牛产奶数据资料,一个产奶周期平均产奶量为2 531.32±641.45kg,产奶量最高可达3 615.7 kg,最低的为1 543.4kg[3]。阐明槟榔江水牛产奶性状的分子基础,为今后开展产奶性能标记辅助选择,提高槟榔江水牛的产奶性能具有重要的意义。
信号转导及转录激活因子(STAT5A)主要参与哺乳动物乳腺组织中细胞因子的信号转导,对泌乳活动具有重要的调控作用。STAT5A也被称作乳腺因子,因为其主要调控靶基因中催乳素和生长激素的活动[4,5],并且已经有研究证明乳蛋白基因的表达受催乳素和肾上腺皮质激素的协同调控[6,7],对奶牛注射生长激素对奶牛的产奶量有提高作用[8]。许多研究也证明了STAT5A蛋白与奶牛的产奶性能有着密切的联系。Maria Selvaggi在意大利布朗牛STAT5A基因中发现多态性,并且CC和CT基因型与产奶量和乳蛋白含量有非常明显的相关性。CC基因型比CT基因型产奶量高,CC基因型乳蛋白质含量也高于CT基因型,但乳脂含量没有明显的差异[9]。Pawe Brym在荷斯坦和新泽西州牛的内含子9发现一个SNP(A/G)。对荷斯坦牛来说,不同的基因型与泌乳性状之间没有明显的相关性,但在新泽西牛中,不同基因型与头胎次和二胎次泌乳量,乳蛋白率和乳脂率之间存在着明显的相关性,GG型牛有高产奶量,而AA和AG型牛蛋白质产量高[10]。我们前期的研究发现STAT5A基因在槟榔江水牛上有高度多态性,为本研究继续开展多态性与产奶性能的关联研究奠定了基础。
2 材料与方法
2.1 试验材料
83头槟榔江水牛耳组织样本采自云南省腾冲县巴福乐槟榔江水牛良种繁育公司。浸泡于无水酒精中于-20℃保存备用。鉴于槟榔江水牛第三胎产奶量最高,本研究采用槟榔江水牛第三胎次产奶量、乳蛋白率和乳脂率数据为产奶性状资料。
2.2 试验方法
2.2.1 基因组DNA的提取 采用传统的苯酚/氯仿抽提法。
2.2.2 引物合成 根据NCBI上提供的牛(Bostaurus)的STAT5A全基因组序列(Gene bank登录号:NW_003104496.1)利用primer3在线设计3对引物,引物由上海生工生物工程公司合成。引物序列、PCR产物大小、PCR体系与条件见文献[11]。
2.2.3 序列分析及数据分析 采用DNAStar软件包对测序结果进行分析,采用人工校正。选用适当的限制性内切酶建立PCR-RFLP检测方法。等位基因频率、基因型频率及Hardy-Weinberg遗传平衡分析采用SAS9.0进行。
2.2.4 PCR-RFLP 通过对引物 P3(5'-GGCTGGAACTACACCTTCTGG-3',5'-GGGTGTTCTCGTTCTTGAGC-3')扩增产物片段测序结果进行分析,结果发现在内含子15的一个C/T碱基替换产生了一个MspI限制性内切酶位点,因此本研究建立了PCR-RFLP分子标记技术。酶切反应体系为20 μL:10×酶解缓冲液2μL,MspI(20I/μL)0.5μL,4 μL PCR产物,加水至20μL,37℃水浴4h以上。MspI酶切位点由C和G两个等位基因控制,经酶切得到三种基因型,分别为CC,CG,GG。基因型为CC的片段被切成452bp和330bp两条带;杂合子CG为782bp,452bp和330bp三条带;基因型为GG的片段只有一条782bp的条带,未被切开。
2.2.5 产奶性状分析 本研究中的产奶量、乳脂率、乳蛋白率均为牛场DHI生产记录数据。根据最小二乘法建立下列模型:
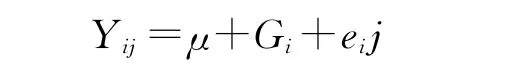
式中Yij为表型值;μ为群体平均值;Gi为第i种基因型效应;eij为随机误差。根据以上线性模型,利用SAS9.0对产奶数据进行分析。
3 结果与分析
槟榔江水牛耳组织基因组DNA的提取及STAT5A基因PCR扩增结果良好,可以用于测序和酶切反应。为了检测槟榔江水牛STAT5A基因多态性,选择8个具有特征特性的水牛个体的DNA用三对引物进行扩增,然后测序。通过对所有测序结果进行分析,共发现38个变异位点。其中引物P1扩增产物中发现24个变异位点;引物P2扩增产物中发现11个变异位点;引物P3扩增产物中发现3个变异位点。所检测到的所有变异位点均为单核苷酸变异(SNP),所有变异位点主要以转换、颠换为主,未发现碱基插入或缺失。其中外显子8产生2处C924T,C975T突变,氨基酸分析表明,没有导致308位氨基酸(脯氨酸)和325位氨基酸(丝氨酸)的变化;外显子13产生2处G1 482A,C1 548T突变,没有导致494位氨基酸(脯氨酸)和516位氨基酸(丙氨酸)的变化。具体结果见文献[11]。
3.1 槟榔江水牛STAT 5A基因序列测定及基因分型
本研究对所有个体进行了基因分型,选取了STAT5A基因变异中4个外显子变异位点(924/E8,975/E8,1482/E13,1548/E13)进行分析。各位点测序结果见图1~4。
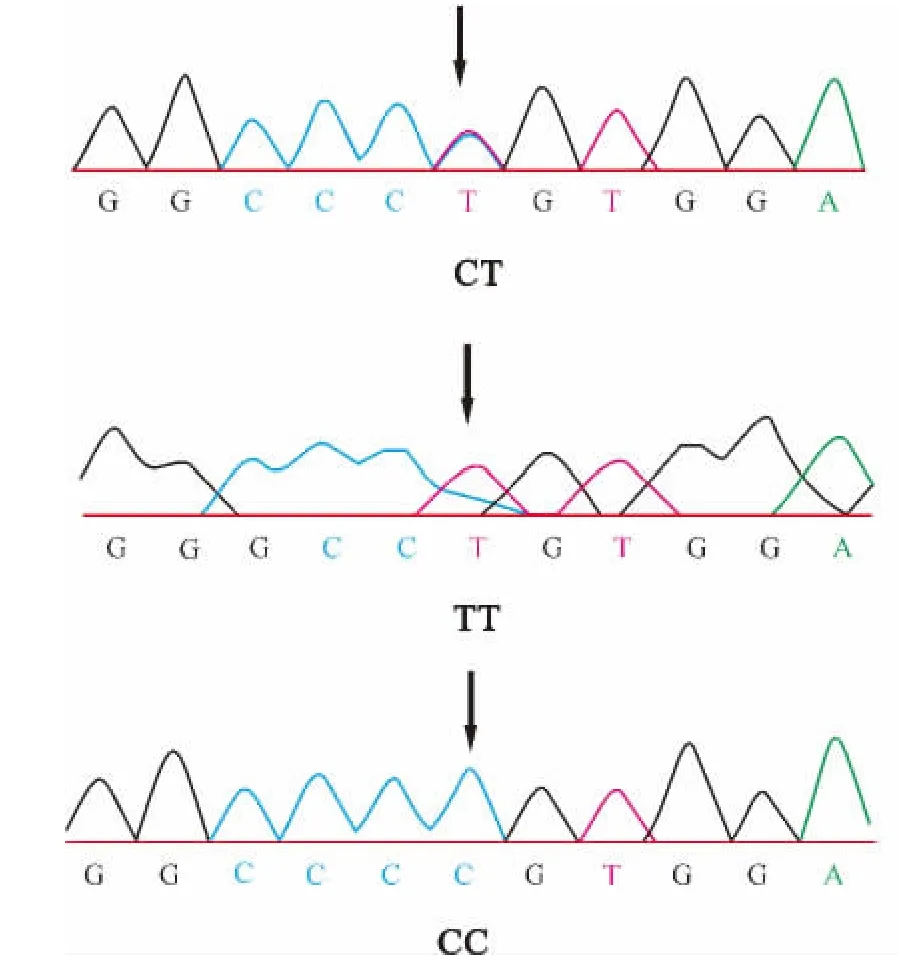
图1 924/E8变异位点序列
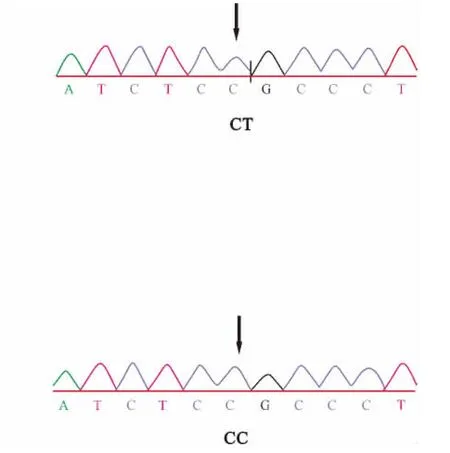
图2 975/E8变异位点序列
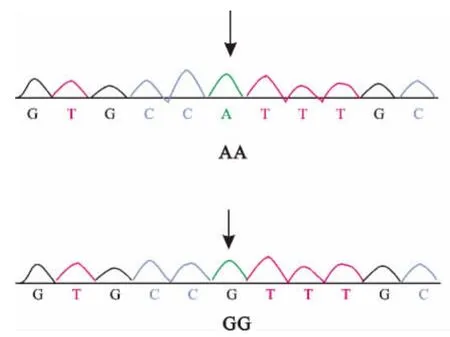
图3 1482/E13变异位点序列

图4 1548/E13变异位点序列
3.2 槟榔江水牛STAT5A基因 MspI标记建立
用MspI限制性内切酶对全部83个样品进行酶切反应,得到的结果如下。图5为扩增产物酶切电泳结果,表1为对MspI酶切位点统计的基因型频率和基因频率。
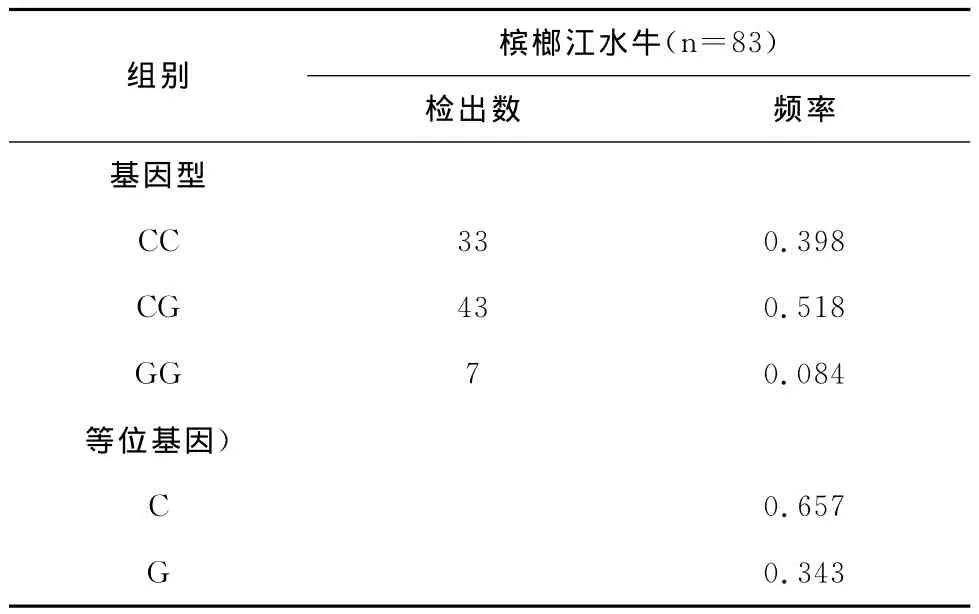
表1 引物P3扩增产物PCR-RFLP检测到的基因型频率和等位基因频率
通过计算发现,CG为优势基因型,C为优势基因,等位基因C的频率明显高于等位基因G的频率。
3.3 槟榔江水牛STAT5A基因多态信息分析
对4个外显子变异及MspI酶切位点进行多态信息分析,利用遗传学理论对每个位点的多态信息含量、杂合度和有效等位基因数进行分析并进行χ2适合性检验,结果见表2。
由统计结果可以看出,在924/E8变异位点,群体多态信息含量(PIC)为0.336,达到中度多态性(0.25<PIC<0.5)。有效等位基因数接近观察到的实际等位基因数。在975/E8变异位点中,PIC为0.100,属于低度多态(PIC<0.25),杂合度和有效基因数的数值也反映该位点遗传变异较小。在1482/E13变异位点,PIC为0.346,达到中度多态,杂合度和有效等位基因数的数值也反映相同的趋势。在外显子1548/E13变异位点,PIC为0.240,属于低度多态,遗传变异较小,杂合度和有效等位基因数也反映了这一趋势。MspI酶切位点的PIC为0.349,达到中度多态性。χ2适合性检验结果显示,5个变异位点差异水平均不显著(P>0.05),处于Hardy-Weinberg平衡状态。
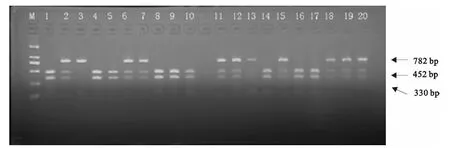
图5 引物P3扩增产物MspI酶切结果M:DGL 2000DNA Marker
3.4 槟榔江水牛STAT5A基因多态性与槟榔江水牛产奶性状的关联性分析
选取STAT5A基因变异中4个外显子变异位点(924/E8,975/E8,1482/E13,1548/E13)和 MspI酶切位点分别与槟榔江水牛第三胎次的产奶量、乳蛋白率和乳脂率进行关联性分析,结果如下:
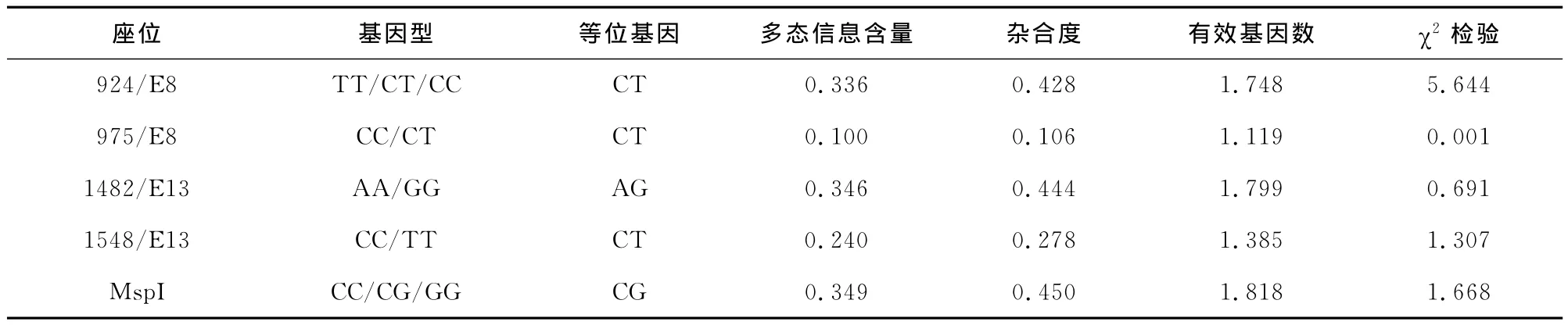
表2 槟榔江水牛STAT5A基因变异位点的遗传特性

表3 924/E8变异位点多态性与槟榔江水牛产奶性状的关联性分析
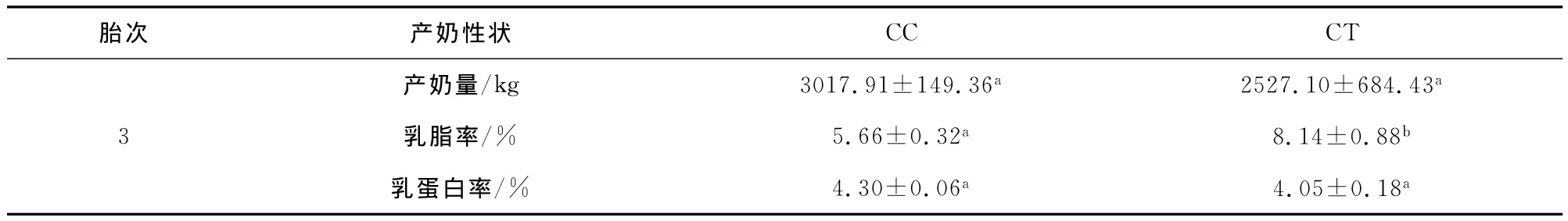
表4 975/E8变异位点多态性与槟榔江水牛产奶性状的关联性分析
分别将STAT5A基因外显子8的两个变异位点的不同基因型与槟榔江水牛第三胎产奶性状进行关联分析,结果表明:在924/E8位点,CT基因型个体的产奶量极显著高于TT基因型个体(P<0.01),三种基因型对乳蛋白率及乳脂率均没有显著影响(P>0.05);在975/E8位点,CT基因型个体的乳脂率显著高于CC基因型个体(P<0.05),产奶量和乳蛋白率在两种基因型之间没有显著差异(P>0.05)。分析结果见表3和4。
分别将STAT5A基因外显子13的两个变异位点的不同基因型与槟榔江水牛第三胎产奶性状进行关联分析,结果表明:在1482/E13位点,产奶量,乳蛋白率和乳脂率在两种基因型之间没有显著差异;在1548/E13位点,产奶量,乳蛋白率和乳脂率在两种基因型之间也没有显著差异。分析结果见表5和6。
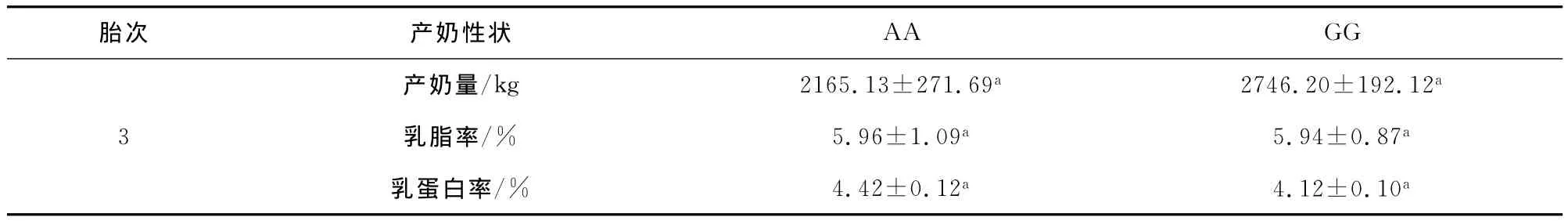
表5 1482/E13变异位点多态性与槟榔江水牛产奶性状的关联性分析
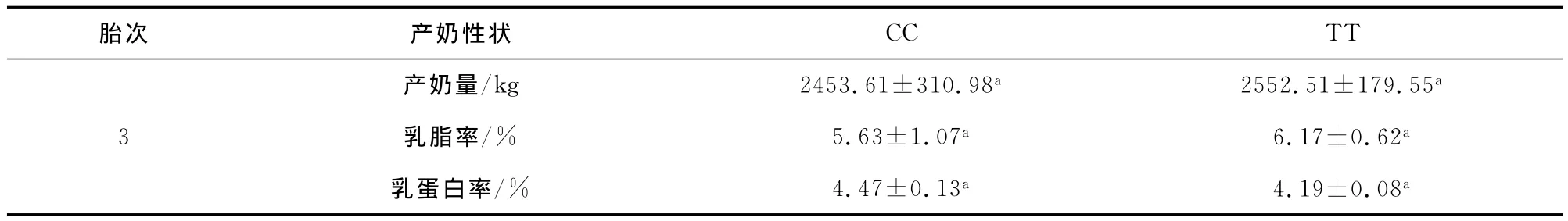
表6 1548/E13变异位点多态性与槟榔江水牛产奶性状的关联性分析

表7 STAT5A基因MspI酶切位点基因多态性与槟榔江水牛产奶性状的关联性分析
将STAT5A基因的MspI位点与槟榔江水牛的产奶性状进行关联分析,得到的结果如下:CG基因型个体产奶量和乳脂率极显著高于CC基因型个体(P<0.01),乳蛋白率在三种基因型个体之间没有显著差异(P>0.05)。结果见表7。
4 讨论
4.1 槟榔江水牛STAT5A基因序列显示高度多态
检测到的38个变异位点中,发生在内含子的变异有34个,外显子变异有4个。本实验是首次以槟榔江水牛为样本对STAT5A作检测,他人未作过相关报道,因而无从比较。但是发生在槟榔江水牛STAT5A基因上的变异与有关报道发生在其他牛STAT5A基因上的变异相比较还是不同的。Maria Selvaggi运用PCR-RFLP(AvaI酶切)技术在意大利布朗牛发现多态性,它是位于外显子7第6 853位置的一个C/T碱基替换[9],Flisikowski等对夏洛莱,红安格斯,利木赞,海福特,西门塔尔及荷斯坦公牛进行了STAT5A外显子7单核苷酸多态性的研究,结果表明在STAT5A基因外显子7存在一个SSCP位点[12];Khatib等人在奶牛STAT5A基因外显子8中发现一个SNP位点是G/C碱基替换[13];McCracken等人[14]在STAT5A 基因内含子12中发现了不同长度的TG重复;Flisikowski等人在牛内含子15中发现CCT缺失,而在外显子16中发现一个 T/C突变[15,16];2005年Jang等报道了在韩国奶牛STAT5A基因内含子15检测到1个SNP变异位点[17]。通过比较可以看出,STAT5A基因在槟榔江水牛上的变异与在其他牛上检测到的变异不同并且远远高于在其他牛上的变异,这也反映了槟榔江水牛STAT5A基因高度的多态性。此外Flisikowski等人还在STAT5A基因的启动子区发现两个SNP位点,分别是-247A/G替换和-226G/A替换[18]。本研究未对此区域作检测,因而无法比较。出现这种情况可能与槟榔江水牛所处的地理环境有关。腾冲县位于云南省保山市西南部,紧邻孟加拉湾和东南亚地区,西部与缅甸毗邻,历史上曾是古西南丝绸之路的要冲,腾冲县的地理位置决定了槟榔江水牛与亚洲其他水牛种群之间的交流十分频繁,而LEI等的研究也揭示该地区属于沼泽型水牛和河流型水牛的混杂之地[19],这可能是导致槟榔江水牛的遗传变异十分丰富的原因。
本研究对5个变异位点作了多态信息含量、有效等位基因数和遗传杂合度的分析,其中有3个位点达到了中度多态,一个位点非常接近中度多态,只有一个位点属于低度多态,有效等位基因数和杂合度也反映了相同的趋势,这说明槟榔江水牛群体遗传变异程度较高,遗传多样性较丰富。χ2检验结果显示所有位点都达到了 Hardy-Weinberg平衡,这表明在此位点的基因型频率和基因频率将稳定遗传,说明了槟榔江水牛STAT5A基因在有丰富的遗传变异的同时也具有一定的遗传稳定性。
4.2 STAT5A基因多态性与槟榔江水牛产奶性状间有显著关联性
STAT5A基因作为产奶性能的候选基因之一,一直是遗传学研究的热点内容。本研究选择了STAT5A基因的5个变异位点与槟榔江水牛第三胎产奶量、乳蛋白率、乳脂率等产奶性状进行了关联性分析,以期寻找与数量性状位点相关的遗传标记。分析发现,5个变异位点中有3个位点与产奶性能有显著性相关。在975/E8位点,虽然产奶量和乳蛋白率在两种基因型之间没有显著差异(P>0.05),但呈现CC>CT的趋势。在MspI酶切位点,CG基因型个体产奶量和乳脂率显著高于CC基因型个体(P<0.01),乳蛋白率在三种基因型个体之间没有显著差异(P>0.05),但也呈现CG>CC>GG的趋势,由此可以看出CG基因型对槟榔江水牛的产奶性能起到了正面的效应。
本研究结果与其他关于STAT5A基因与产奶性能关联的研究结果一致。Maria Selvaggi运用PCR-RFLP(AvaI酶切)技术在意大利布朗牛上作的STAT5A基因与产奶性能关联性分析结果发现,外显子7第6 853位置的一个C/T碱基替换,并且CC和CT基因型与产奶量,乳蛋白含量有非常明显的关联性。CC基因型比CT基因型产奶量高,CC基因型乳蛋白质含量也高于CT基因型,但乳脂含量没有明显的差异[9]。Brym等在2004年报道了STAT5A基因内含子9的一个SNP(A/G),在186头荷斯坦牛中,不同的基因型与泌乳性状之间没有明显的相关性,而在138头新泽西牛中,不同基因型与头胎次和二胎次泌乳量,乳蛋白率和乳脂率之间存在着明显的相关性,GG型牛有高产奶量,而AA和AG型牛蛋白质产量高[10]。何峰等在2007年对758头中国荷斯坦奶牛进行了STAT5A基因多态性检测,并将其与5个产奶性状进行了关联分析。结果发现,在内含子9的一个SNP(A/G)对乳蛋白率有显著影响(P<0.05),12 440位T→C和12 550位的CCT插入/缺失,此位点对产奶量有极显著影响(P<0.01),对乳脂量、乳蛋白量有显著影响(P<0.05)[20]。
通过本研究和国内外其他的研究可以看出,STAT5A基因多态性在多种牛上检测到了其与产奶性能的显著关联性,因此本研究认为STAT5A基因可以作为与产奶性能有关的候选基因。前人的研究发现槟榔江水牛产奶性比云南当地饲养的其他河流型水牛和沼泽性水牛产奶天数短,产奶量却高,但个体间产奶性能差异很大[1]。本研究检测了5个变异位点与产奶性能的关系,这为提高槟榔江水牛的产奶性能作了很好的铺垫,特别是与产奶性能有相关性的变异位点可以作为分子标记而应用,如逐步固定对产奶性能有正面效用的基因型,以提高生产性能。同时这些变异位点可以整合到槟榔江水牛选育方案中,建立产奶性能标记辅助选择。
4.3 槟榔江水牛STAT5A基因MspI酶切位点可作为一个有效分子标记
在MspI酶切位点χ2检验达到了Hardy-Weinberg平衡,这说明在此位点的基因型频率和基因频率将稳定遗传,处于遗传平衡状态。PIC为0.349,达到中度多态,这说明该遗传标记可以提供比较丰富和合理的遗传信息。酶切结果显示,此酶切位点CG基因型频率高于CC和GG基因型频率,说明在此位点杂合子是优势基因型,在特定的条件下杂合子个体可能比正常纯合子个体更有利于生存和繁殖后代。PCR-RFLP技术操作简便,分型时间短,特异性强,并且MspI限制性内切酶价格便宜,性能稳定,是十分常用的一种内切酶。因此在大规模群体检测中此位点可以作为一个快速简便并且经济的有效分子遗传标记。
[1] 屈在久,沈雪鹰,杨加用,等.腾冲槟榔江水牛的起源及初步研究[J].云南畜牧兽医,2008,(增刊):34-37.
[2] 章纯熙,苗永旺,李大林.我国首例本土河流型水牛-槟榔江水牛的种质特征[J].家畜生态学报,2011,32(6):39-45.
[3] 屈在久,李大林,苗永旺.槟榔江水牛种质资源调查与评价[J].云南农业大学学报,2008,23(2):265-269.
[4] Wakao H,Gouilleux F,Groener B,et al.Mammary gland factor(MGF)is a novel member of the cytokine-regulated transcription factor gene family and confers the prolactin response[J].EMBO 13(9):2182-2191.
[5] Argetsinger L S,Carter-Su C.Growth hormone signalling mechanisms:involvement of the tyrosine kinase JAK2[J].Hormone Research,1996,4522-4524.
[6] Ball R K,Friis R R,Schoenenberger C A,et al.Prolactin regulation ofβ-casein gene expression and of a cytosolic 120KD protein in a cloned mouse mammary epithelial cell line[J].EMBO,1988,7:2089-2095.
[7] Lee C S,Kim K,Yu D Y,et al.Pretreatment with glucocorticoid is essential for lactogenic induction of the bovine beta-casein/CAT expression in HC11cells[J].Endocrine Research,1998,24:65-77.
[8] Bauman D E,Vernon R G.Effects of exogenous bovine somatotropin on lactation[J].Annual review of nutrition,1993,13:437-461.
[9] Maria Selvaggi,Cataldo Dario,Giovanni Normanno,et al.Genetic polymorphism of STAT5Aprotein:relationships with production traits and milk composition in Italian Brown cattle[J].Journal of Dairy Research,2009,76(4):441-445.
[10] Pawe Brym,Stanisaw Kamñski,Anna Ruceae.New SSCP polymorphism within bovine STAT5Agene and its associations with milk performance traits in Black-and-White and Jersey cattle[J].Appl Genet,2004,45(4):445-452.
[11] 季 敏,余长林,余选富,等.槟榔江水牛STAT5A基因多态性及 MspI PCR-RFLP遗传标记的建立[J].中国牛业科学,2012,38(6):12-18.
[12] Flisikowski K,Zwierzchowski L.Single strand conformation polymorphism with exon 7of the bovine STAT5Agene[J].Animal Science Papers and Reports,2002,2(20):133-137.
[13] Khatib H,Monson R L,Schutzkus V,et al.Mutations in the STAT5Agene are associated with embryonic survival and milk composition in cattle[J].Journal of Dairy Science,2008,91:784-793.
[14] McCracken J Y,Molenaar A J,Snell R J,et al.A polymorphic TG repeat present within the bovine STAT5Agene[J].Animal Genetics.1997,28,453-464.
[15] Flisikowski K,Szymanowska M,Zwierzchowski L.The DNA-binding capacity of genetic variants of the bovine STAT5A transcription factor[J].Celluar & Molecular Biology Letters,2003,8:831-840.
[16] Flisikowski K,Zwierzchowski L.Polymerase chain reactionheteroduplex (PCR-HD)polymorphism within the bovine STAT5Agene[J].Appl Genet,2003,44:185-189.
[17] Jang G W,Cho K H,Kim T H,et al.Association of Candidate genes with production traits in Korean dairy Proven and Young bulls[J].Asian-Aust J Anim Sci,2005,18(2):165-169.
[18] Flisikowski K,Hiendleder S,Zwierzchowski L.Nucleotide Sequence Variation in the Transcription Factor STAT5Agene 5'-noncoding Region in Bostaurus and Bos indicus Cattle[J].Biochemical Genetics,2005,43:459-464.
[19] Lei C Z,Zhang W,Chen H,et al.Independent maternal origion of Chinese swamp buffalo(Bubalus bubalis)[J].AnimGenet,2007,38(2):97-102.
[20] 何 峰,孙东晓,俞 英,等.荷斯坦奶牛STAT5A基因的SNPs检测及其与产奶性状的关联分析[J].畜牧兽医学报,2007,38(4):326-331.
猜你喜欢
动物营养学报(2022年9期)2022-10-12
农村农业农民·B版(2021年11期)2021-12-03
阅读与作文(小学低年级版)(2021年2期)2021-05-08
中国畜牧杂志(2021年1期)2021-01-20
中国畜禽种业(2020年9期)2020-12-16
中国乳业(2019年12期)2020-01-17
河南畜牧兽医(2020年1期)2020-01-11
中国奶牛(2019年12期)2020-01-08
分析化学(2018年4期)2018-11-02
现代畜牧科技(2018年3期)2018-05-14
