
饲粮策略对不同遗传品系二郎山山地鸡生产性能和屠宰性能的影响
2013-09-20 03:04丁雪梅白世平曾秋凤罗玉衡张克英
动物营养学报 2013年9期
张 凯 丁雪梅 白世平 曾秋凤罗玉衡 朱 庆 张克英*
(1.四川农业大学动物营养研究所,动物抗病营养教育部重点实验室,雅安 625014;2.四川农业大学动物遗传育种研究所,雅安 625014)
二郎山山地鸡是以我国优良地方鸡种为素材,由四川农业大学和雅安隆生农牧有限公司进行选育的,但目前关于二郎山山地鸡营养需要的研究很少,本课题组已开展了二郎山山地鸡饲粮适宜能量和蛋白质水平[1-2]、维生素预混料[3]的相关研究,但对各阶段适宜的饲粮赖氨酸水平尚未开展研究。为促进其规模化养殖生产,有必要针对二郎山山地鸡饲粮赖氨酸水平开展系统研究。赖氨酸作为肉鸡第二限制性氨基酸,具有多种重要生理功能:参与体蛋白,如骨骼肌、酶和多肽激素的合成;是生酮氨基酸之一,当缺乏可利用的碳水化合物时,它参与生成酮体和葡萄糖的代谢;维持体内酸碱平衡;作为合成肉毒碱的前体物,参与脂肪代谢;还可以提高机体抵抗应激的能力[4]。关于肉鸡赖氨酸的需要量,虽然不同研究者给出了不同的推荐值,但都证实NRC(1994)对于商品肉鸡的赖氨酸推荐值已经偏低[5-7]。研究表明,饲粮添加赖氨酸能提高肉鸡生产性能,且全期提高饲粮赖氨酸水平效果更好[8]。为此,本试验旨在研究2种饲粮策略对不同遗传品系和性别的二郎山山地鸡生产性能和屠宰性能的影响,为进一步优选二郎山山地鸡遗传品系和合理饲养提供试验依据。
1 材料与方法
1.1 试验设计
试验采用随机化4×2×2因子设计,4个二郎山山地鸡遗传品系[SD02、SD03、SD03×SD02(SD0302)、SD02×SD03(SD0203)]、2个饲粮策略[1~28 d、29~49 d、50~67 d的赖氨酸水平分别为1.15% ~1.00% ~0.85%(高赖氨酸,HL)、1.00% ~0.85% ~0.70%(低赖氨酸,LL),同阶段内主要饲料原料含量保持一致]、公母2个性别。
1.2 试验动物与饲粮
试验选用1日龄健康二郎山山地鸡1 920只,每个品系公母各240只,按初始体重无差异原则分别随机分配到各处理,每个处理公母各4个重复,每个重复30只鸡。
基础饲粮的能量和蛋白质水平参考本课题组吕铭翰等[1]、张效先等[2]关于二郎山山地鸡 1 ~28 d、29~49 d、50~67 d 3 个阶段饲粮能量、蛋白质需要量的研究结果确定,饲粮组成及营养水平见表1。各阶段饲粮保证原料水平一致,L-赖氨酸盐酸盐含量差异部分由膨润土调节。饲粮为颗粒料,前期制粒粒径1.5 mm(需破碎),中期2.5 mm,后期3.0 mm。
1.3 饲养管理
试验在四川农业大学动物营养研究所教学科研基地进行。采用地面垫料平养,自由采食和饮水。舍内红外灯供热,第1周舍内温度为32~35℃,之后每周降温2~3℃,4周后常温饲养。鸡舍湿度维持在60% ~65%;光照维持24 h/d。舍内自然通风,定期消毒。饲养管理按照常规程序进行。按照常规免疫程序对鸡群进行免疫接种。
1.4 测定指标
1.4.1 生产性能
体重(body weight,BW)和体增重(body weight gain,BWG):于试验第 1、29、50 和 68 天早晨以重复为单位进行称重。计算每个重复的平均阶段增重。称重前12 h断料。
采食量(feed intake,FI):按阶段称量每个重复的剩余料和损耗料,计算每个重复的采食量。
料重比(feed to gain ratio,F/G):按阶段计算各重复阶段料重比。料重比=阶段采食量/阶段增重。
死淘率(mortality):记录各阶段鸡只死亡、淘汰情况(体重、时间),计算阶段死淘率。
1.4.2 屠宰性能
参考杨宁等[9]《家禽生产学》鸡屠宰实验指导,于试验第68天09:00从每个重复中选取与平均体重相近的4只鸡颈部放血致死,在70℃左右热水中去毛,用纸巾擦干水分后进行屠宰测定,测定指标为活体重、屠体重、半净膛重、全净膛重、左侧胸肌重、左侧腿肌重、腹脂重、皮下脂肪厚,计算屠宰率、半净膛率、全净膛率、胸肌率、腿肌率、腹脂率。
全净膛重为屠体(胸、腹部内只留下肺脏和肾脏,其余器官全部去掉)去头和脚(跗关节以下)的重量。

1.4.3 肉品质
选取屠体的左侧胸肌进行肉品质测定,测定指标为肉色(红度、黄度、亮度值分别为a*、b*和L*值)和屠宰后45 min胸肉pH。
1.4.4 血清生化指标
试验第28天,每个重复挑选2只接近平均体重的鸡,空腹12 h后在次日08:00采血,每只鸡采血5 mL,血样室温放置3 h后再离心2 min(4℃、10 000 r/min),制备血清,-20℃保存待用。
采用全自动生化分析仪(TMS-1024i)测定血清中的总蛋白(total protein,TP)、尿酸(uric acid,UA)、尿素氮(urea nitrogen,UN)、总胆固醇(total cholesterol,TC)、甘油三酯(triacylglycerol,TG)含量。
1.5 数据处理与统计分析
试验采用SPSS 17.0一般线性模型(general linear model,GLM)进行三因子方差分析,主效应包括赖氨酸、遗传品系、性别效应及两两之间和三因素间的交互效应,P<0.05时为差异显著;用Duncan氏法进行多重比较;试验结果用平均值和SEM表示。

表1 饲粮组成及营养水平(风干基础)Table 1 Composition and nutrient levels of diets(air-dry basis) %
2 结果
2.1 饲粮策略对不同遗传品系二郎山山地鸡死淘率的影响
由表2可知,各处理的死淘率差异均不显著(P>0.05);4个遗传品系间死淘率差异不显著(P>0.05);性别对山地鸡死淘率无显著影响(P>0.05);但高赖氨酸饲粮组显著提高了1~67 d死淘率(P<0.05);在各阶段死淘率上,饲粮策略与性别或遗传品系及三者间无显著的交互效应(P>0.05)。
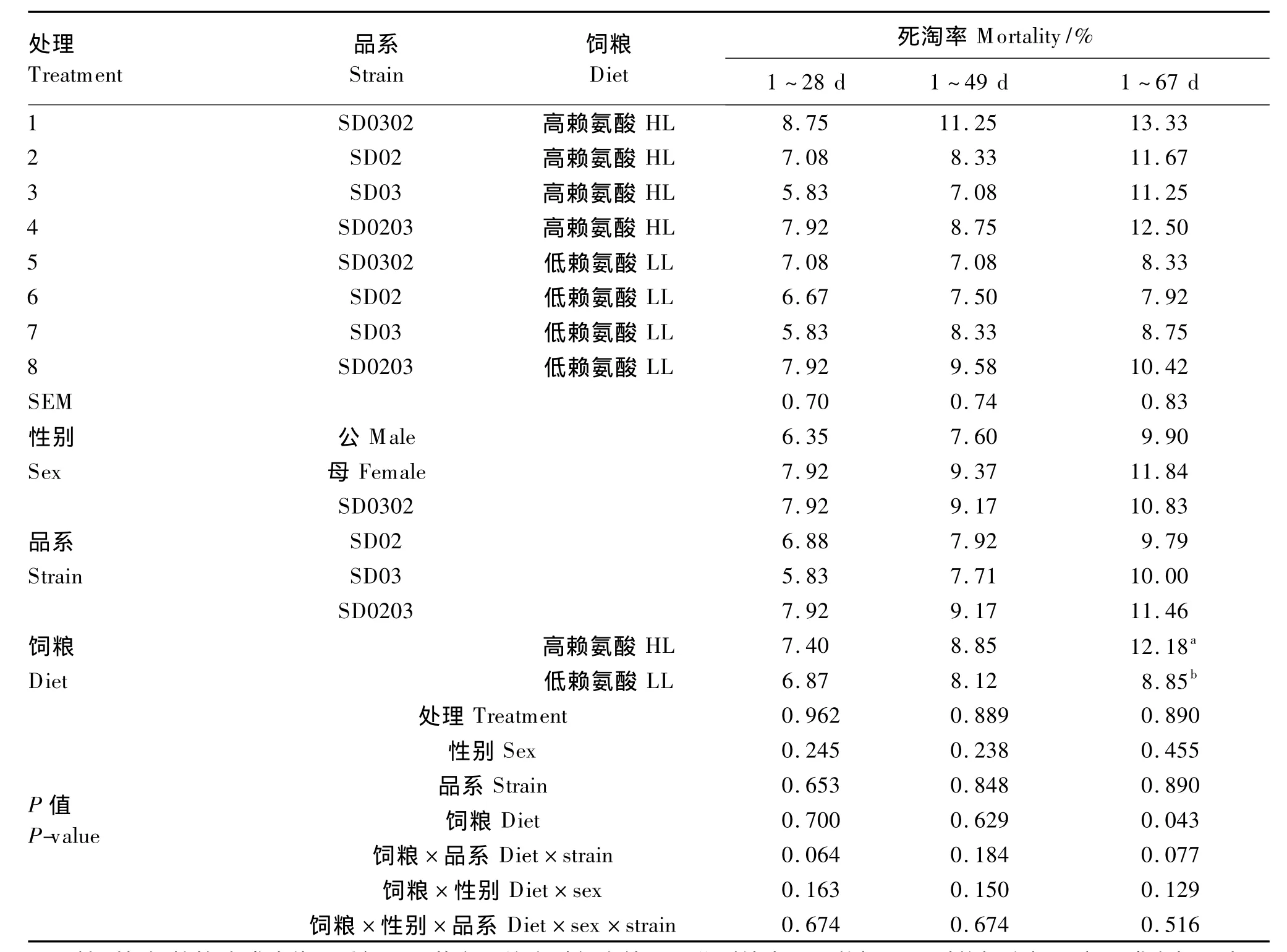
表2 饲粮策略对不同遗传品系二郎山山地鸡死淘率的影响Table 2 Effects of dietary strategy on mortality of Erlang Mountain chickens from different genetic strains
2.2 饲粮策略对不同遗传品系二郎山山地鸡生产性能的影响
由表3可知,体重:各处理间在28、67 d时差异极显著(P<0.01),在49 d时差异显著(P<0.05),其中28 d时品系SD02低赖氨酸饲粮组最高,品系SD0203高赖氨酸饲粮组最低;49 d时品系SD02高赖氨酸饲粮组最高,品系SD0203低赖氨酸饲粮组最低;67 d时品系SD02高赖氨酸饲粮组最高,品系SD03低赖氨酸饲粮组最低。饲粮策略对28和49 d体重无显著影响(P>0.05),但67 d时高赖氨酸饲粮组体重极显著高于低赖氨酸饲粮组(P<0.01)。不同遗传品系间在28、49和67 d时差异均极显著(P<0.01),其中28和67 d时以品系SD02最高,49 d时以品系 SD0302最高,品系 SD02次之,二者间无显著差异(P>0.05)。性别因素极显著影响各阶段体重(P<0.01),均是公鸡高于母鸡。饲粮策略与遗传品系或性别及三者之间在28、49 d体重上未表现出显著的交互效应(P>0.05),但三者之间在67 d体重上表现出极显著的交互效应(P<0.01),由表4可知,同种赖氨酸水平饲粮下,各品系公鸡的67 d体重均显著高于母鸡(P<0.05);同种性别下,不同饲粮间品系SD0302公母鸡67 d体重差异均不显著(P>0.05),品系SD02和SD03母鸡的67 d体重均差异显著(P<0.05),品系SD0203母鸡67 d体重无显著差异(P>0.05),而公鸡67 d体重差异显著(P<0.05)。
体增重:各处理间1~28 d、1~67 d差异极显著(P<0.01),其中1~28 d以品系SD02低赖氨酸饲粮组最高,品系SD0203高赖氨酸饲粮组最低,1~67 d以品系SD02高赖氨酸饲粮组最高,品系SD03低赖氨酸饲粮组最低;饲粮策略对1~28 d和1~49 d体增重无显著影响(P>0.05),但可极显著影响1~67 d体增重(P<0.01),高赖氨酸饲粮组高于低赖氨酸饲粮组;不同遗传品系间1~28 d、1~49 d和1~67 d体增重差异极显著(P<0.01),均以品系SD02最高,品系SD0203最低;性别因素极显著影响各阶段体增重(P<0.01);饲粮策略与遗传品系或性别及三者之间在1~28 d、1~49 d体增重上未表现出显著的交互效应(P>0.05),但三者之间在1~67 d体增重上表现了极显著的交互效应(P<0.01),由表4可知,同种赖氨酸水平饲粮下,各品系公鸡的1~67 d体增重均显著高于母鸡(P<0.05);同种性别下,不同饲粮间1~67 d体增重差异不显著(P>0.05),公鸡以品系SD0203低赖氨酸饲粮组最低,母鸡以品系SD03低赖氨酸饲粮组最低;品系SD02和SD03母鸡饲喂高赖氨酸饲粮时1~67 d体增重显著高于同品系母鸡饲喂低赖氨酸饲粮(P<0.05)。
采食量:各处理间1~28 d差异极显著(P<0.01),其中品系SD0302和SD02低赖氨酸饲粮组最高,品系SD03高赖氨酸饲粮组最低,而其余2个阶段各处理间采食量差异不显著(P>0.05);饲粮策略对各阶段采食量影响均不显著(P>0.05);不同遗传品系间1~28 d采食量差异极显著(P<0.01),其中品系SD0302和SD02极显著高于品系SD03和SD0203(P<0.01),不同遗传品系间1~49 d体增重差异接近显著水平(P=0.052),1~67 d体增重差异显著(P<0.05),3个阶段均以品系SD0302最高;性别因素极显著影响各阶段采食量(P<0.01),均为公鸡高于母鸡;饲粮策略与遗传品系或性别及三者之间在各阶段采食量上均无显著的交互效应(P>0.05)。
料重比:各处理间1~67 d差异显著(P<0.05),其中以品系SD03高赖氨酸饲粮组最低,品系SD03低赖氨酸饲粮组最高,而其他3个阶段料重比各处理间差异不显著(P>0.05);饲粮策略对1~28 d料重比影响不显著(P>0.05),但高赖氨酸饲粮组1~49 d料重比显著低于低赖氨酸饲粮组(P<0.05),1~67 d料重比极显著低于低赖氨酸饲粮组(P<0.01);不同遗传品系间各阶段料重比差异均不显著(P>0.05);性别因素极显著影响各阶段料重比(P<0.01);饲粮策略与遗传品系或性别及三者之间在各阶段料重比上均无显著的交互效应(P>0.05)。
2.3 饲粮策略对不同遗传品系二郎山山地鸡屠宰性能的影响
由表5可知,各处理间山地鸡腹脂率差异显著(P<0.05),其中以品系SD02高、低赖氨酸饲粮组显著低于其他处理,而其他屠宰性能指标各处理间差异均不显著(P>0.05);饲粮策略对各项屠宰性能指标均无显著影响(P>0.05),但高赖氨酸饲粮组屠宰率、全净膛率和皮脂厚较低,左胸肌率、左腿肌率和腹脂率则较高;不同遗传品系间屠宰率差异显著(P<0.05),腹脂率差异极显著(P<0.01),其中品系SD02的屠宰率显著低于另外3个品系(P<0.05),腹脂率极显著低于其他3个品系(P<0.01);性别因素极显著影响了左胸肌率、左腿肌率和腹脂率(P<0.01),而对其他屠宰性能指标无显著影响(P>0.05);饲粮策略与遗传品系或性别及三者之间在各项屠宰性能指标上均无显著的交互效应(P>0.05)。

?
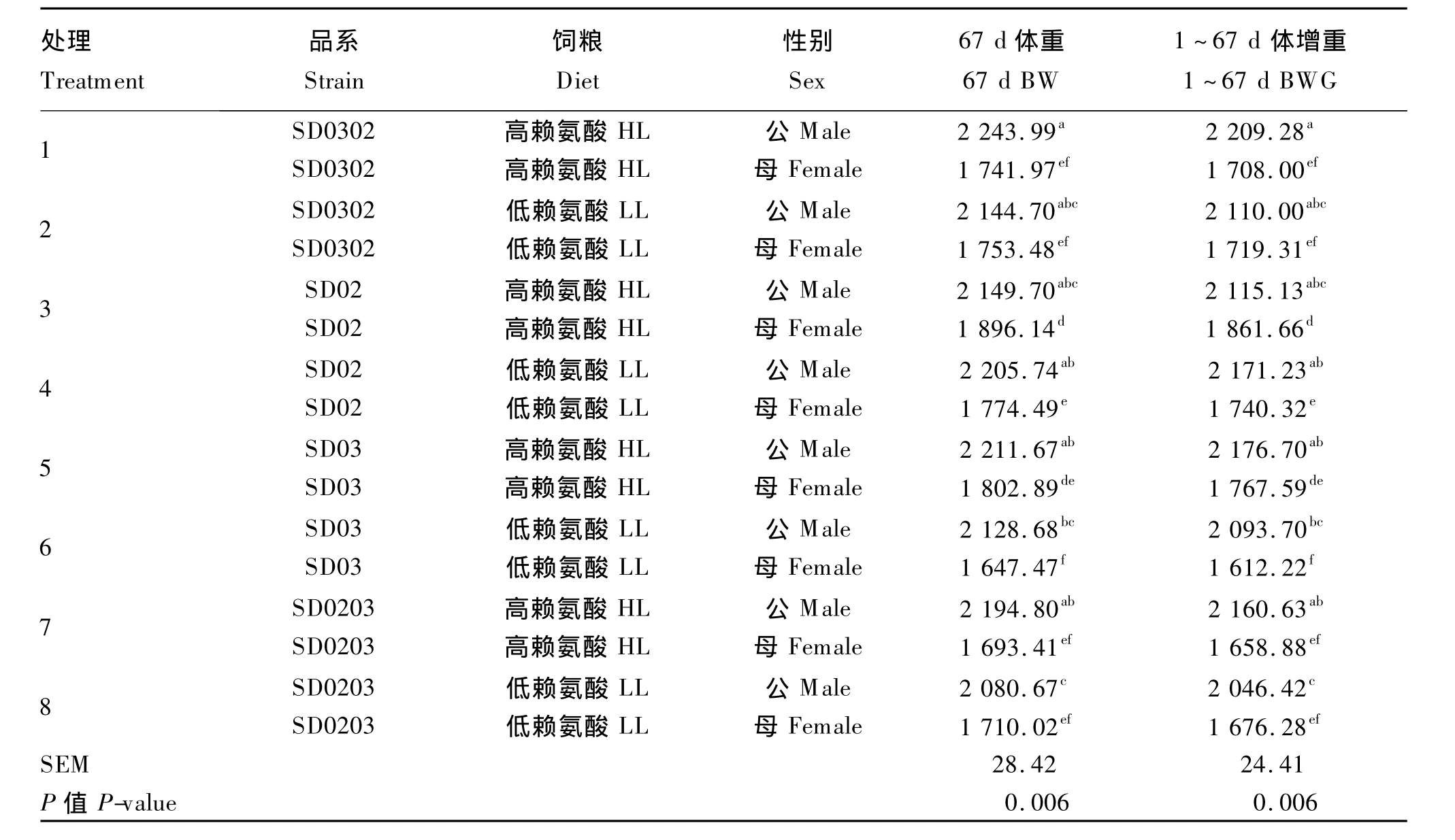
表4 饲粮策略、性别及遗传品系在二郎山山地鸡体重和体增重的交互效应Table 4 The interaction of dietary strategy,sex and genetic strain in body weight and body weight gain of Erlang Mountain chickens g

表5 饲粮策略对不同遗传品系二郎山山地鸡屠宰性能的影响Table 5 Effects of dietary strategy on carcass traits of Erlang Mountain chickens from different genetic strains
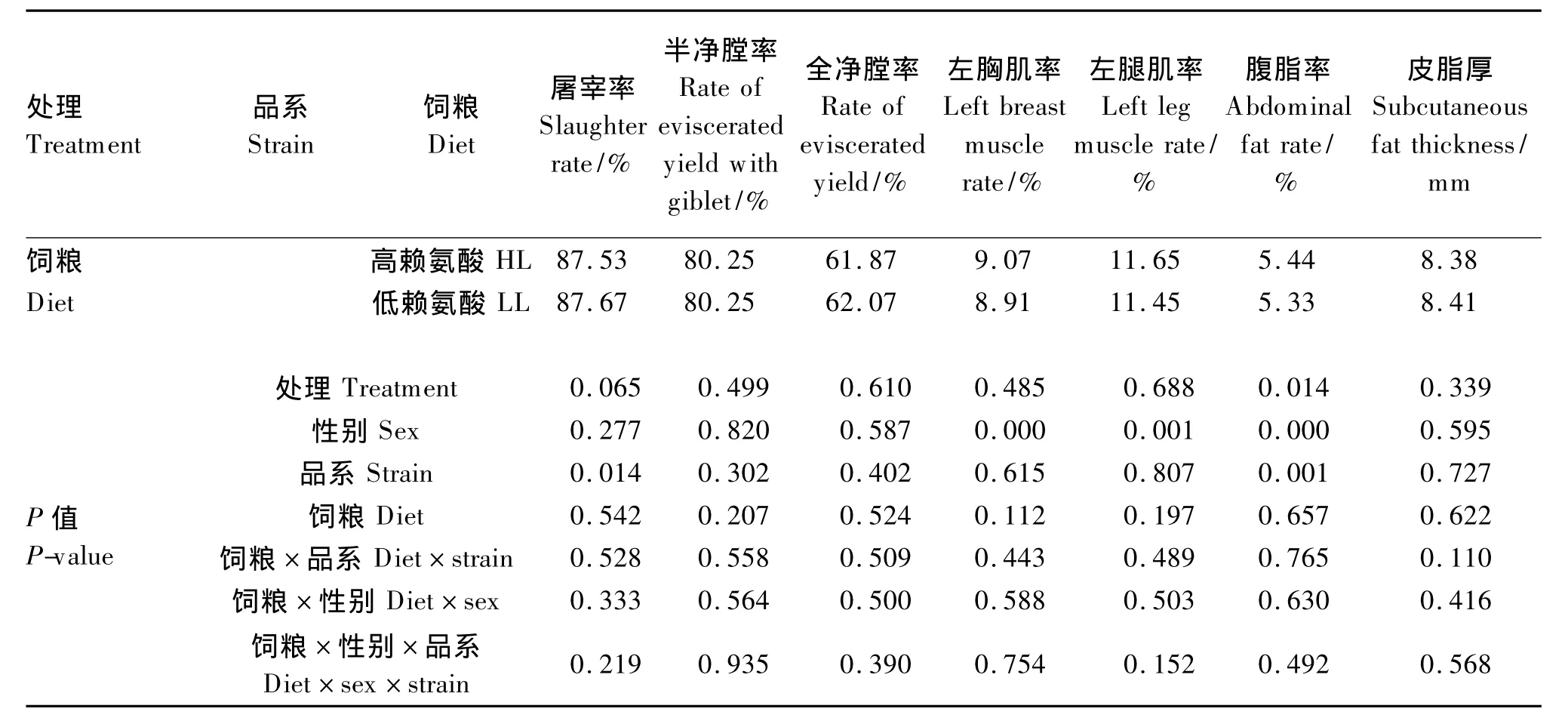
续表5
2.4 饲粮策略对不同遗传品系二郎山山地鸡肉品质的影响
由表6可知,各处理间山地鸡肉色a*、b*和L*值以及屠宰后45 min pH均无显著差异(P>0.05);饲粮策略和不同遗传品系对山地鸡肉色a*、b*和L*值以及屠宰后45 min pH均无显著影响(P>0.05);品系性别因素对山地鸡肉色a*、L*值以及屠宰后45 min pH也无显著影响(P>0.05),但极显著影响肉色b*值(P<0.01),其中公鸡低于母鸡;饲粮策略与遗传品系或性别及三者之间在各项肉品质指标上均无显著的交互效应(P>0.05)。
2.5 饲粮策略对不同遗传品系二郎山山地鸡28 d血清生化指标的影响
由表7可知,各处理间山地鸡28 d各项血清生化指标差异均不显著(P>0.05);饲粮策略极显著影响了28 d山地鸡血清UA和TC含量(P<0.01),其中高赖氨酸饲粮组山地鸡28 d血清UA含量高于低赖氨酸饲粮组,28 d TC含量低于低赖氨酸饲粮组;不同遗传品系间山地鸡28 d各项血清生化指标差异均不显著(P>0.05);性别因素对28 d血清 TP(P<0.05)、TG(P<0.05)和 TC含量(P<0.01)的影响显著,均是公鸡高于母鸡;饲粮策略与遗传品系或性别及三者之间在28 d各项血清生化指标上均无显著的交互效应(P>0.05)。
3 讨论
3.1 饲粮策略对山地鸡生产性能、屠宰性能、肉品质及血清生化指标的影响
赖氨酸作为家禽的第二限制性氨基酸,在肉鸡生长中发挥着重要作用。许多研究报道,饲粮中随着赖氨酸水平的增加,肉鸡生产性能呈现出先升后降的显著趋势,适当添加赖氨酸可提高肉鸡增重,改善料重比;但赖氨酸添加量超过一定水平后又对肉鸡生产性能产生不利影响[10-14]。本试验结果显示,高赖氨酸饲粮组极显著提高了67 d体重和1~67 d体增重,显著降低了1~49 d料重比,极显著降低了1~67 d料重比,但对各阶段采食量无显著影响,这与上述文献报道相符。高赖氨酸饲粮组显著提高了山地鸡1~67 d死淘率,这与 Kidd 等[15]和 Latshaw[16]的研究报道一致,分析原因可能是高赖氨酸饲粮提高了山地鸡的增长速度,但同时也会降低其对环境的耐受力,增加代谢性疾病的发生几率,从而导致死淘率升高。
本试验中饲粮策略对试验前期(1~21 d)、中前期(1~49 d)的体重和体增重均无显著影响,但高赖氨酸饲粮组极显著提高了全期(1~67 d)的体重和体增重,这与Corzo等[17]报道一致,可能是试验前期、中前期低赖氨酸饲粮已基本能满足山地鸡的生长需要,高赖氨酸饲粮的效应不明显,但高赖氨酸饲粮具有累积效应。饲粮策略并未影响山地鸡各阶段的采食量,而山地鸡后期体重显著改善。

表6 饲粮策略对不同遗传品系二郎山山地鸡肉品质的影响Table 6 Effects of dietary strategy on meat quality of Erlang Mountain chickens from different genetic strains
屠宰率和全净膛率是衡量畜禽产肉性能的主要指标。一般认为,屠宰率在80%以上、全净膛率在60%以上即表示肉用性能良好[10]。本试验结果表明,山地鸡高、低赖氨酸饲粮组屠宰率都在87%以上,全净膛率也均超过61%,说明山地鸡具有较好的产肉性能。赵丽红等[10]、刘升军等[12]、Cozro等[17]认为饲喂高水平赖氨酸饲粮的肉鸡能获得较高的屠宰率和胸肉产量;本试验结果显示,饲粮策略对各项屠宰性能无显著差异,并未表现出上述趋势,分析原因可能与2种饲粮策略的赖氨酸水平差值较小有关。赵丽红等[10]研究显示提高饲粮赖氨酸水平能够显著降低公鸡腹脂沉积,Kidd等[15]也指出高水平赖氨酸饲粮能够显著降低母鸡腹脂率,而本试验结果显示饲粮策略并未对肉鸡腹脂率产生显著影响,高赖氨酸饲粮组公母鸡腹脂率均较高,与上述研究不一致;而刘升军等[12]认为当饲粮赖氨酸水平达到1.0%后再继续提高,腹脂率下降不显著。这些研究结果的不同可能与鸡的品种、饲粮组成、环境控制等差异有关。
肉色是肌肉本身生理学、生物化学和微生物复杂变化的结果,主要取决于肌肉中肌红蛋白和血红蛋白的含量。肌红蛋白中的亚铁血红素随着屠宰后时间延长,其中的二价铁转化为三价铁,肌红蛋白被氧化成高铁肌红蛋白,颜色加深。pH不仅反映了肌肉的酸度,同时对肌肉品质具有重要影响,是肉品质测定的重要指标之一。畜禽屠宰后,肌肉组织糖酵解产生的乳酸导致其pH降低,但随着糖酵解的进行,ATP分解生成胺,pH降低,又使糖酵解相关酶活性降低或失活,从而使肌肉的pH达到稳定状态。本试验中饲粮策略对山地鸡胸肉肉色a*、b*和L*值以及屠宰后45 min pH影响均不显著,但高赖氨酸饲粮组的屠宰后45 min pH较高,这与 Berri等[18]结果相符;全期提高饲粮赖氨酸水平可使鸡产肉增加,肌肉中脂肪含量降低,肌糖原含量下降,导致屠宰后肌肉pH较高,颜色加深[19]。
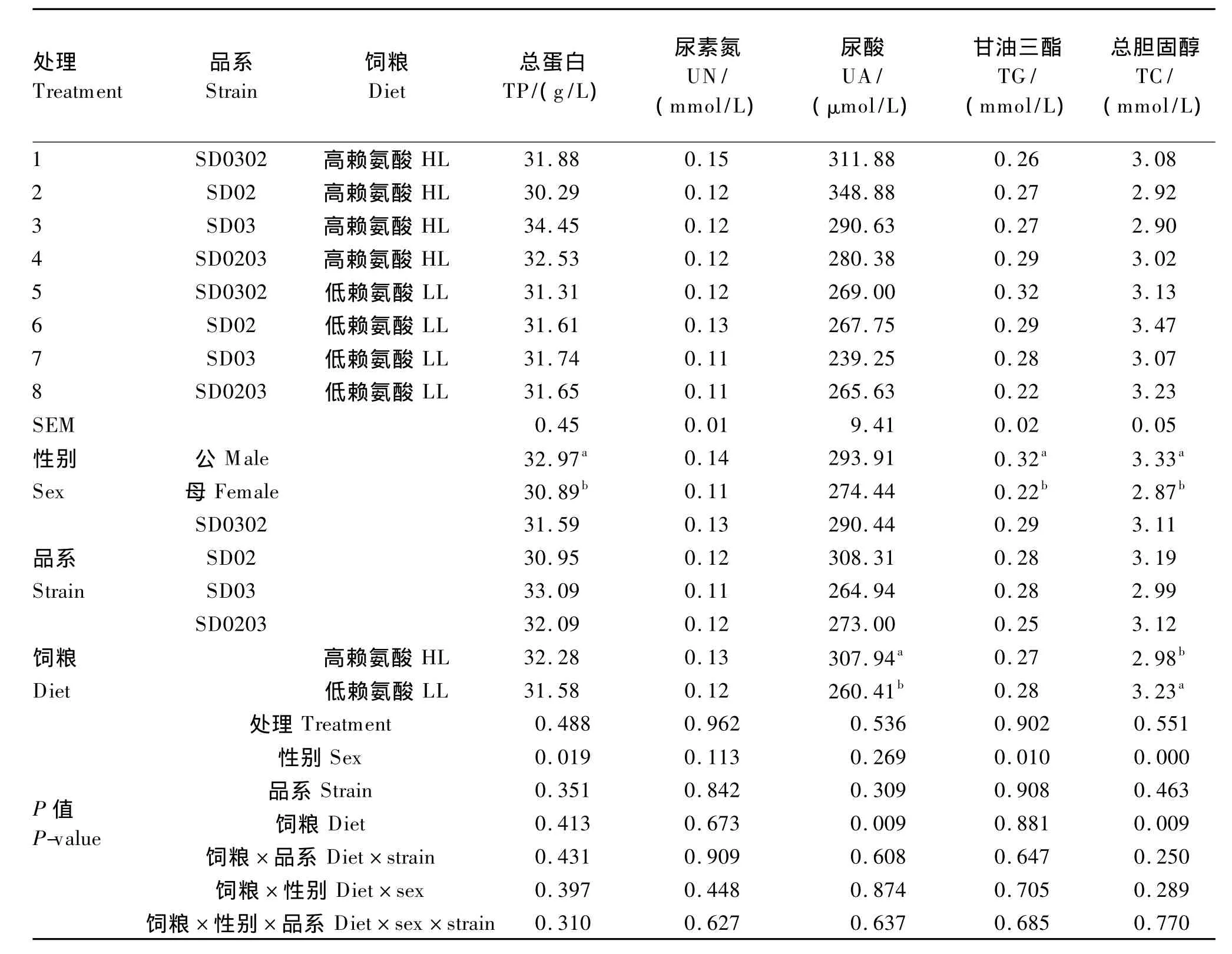
表7 饲粮策略对不同遗传品系二郎山山地鸡28 d血清生化指标的影响Table 7 Effects of dietary strategy on serum biochemical parameters of Erlang Mountain chickens from different genetic strains
血清UA是衡量体内蛋白质营养状况和代谢水平的一个重要指标,家禽体内未被利用的氨基酸经分解代谢后以UA的形式稀释入血,并经肾随尿排出体外。个别氨基酸缺乏或过量将影响自身及其他氨基酸的利用率,使血中UA含量升高。陈志敏等[20]研究表明,饲粮赖氨酸水平0.65% ~1.10%时肉鸡血清的UA含量逐渐降低,饲粮赖氨酸水平1.10%~1.40%时血清UA含量逐渐升高。本试验中高赖氨酸饲粮极显著升高了山地鸡28 d血清UA含量,表明1~28 d高赖氨酸饲粮的赖氨酸含量已经超过了山地鸡所需要的适宜值,这也与1~28 d生产性能低赖氨酸饲粮组较好相符合。
3.2 不同遗传品系对山地鸡生产性能、屠宰性能、肉品质及血清生化指标的影响
动物的遗传基因决定着动物的生长发育,不同品种和品系家禽的遗传特性不同,其生长速度、体格大小、胴体组成以及消化生理都存在差异。Ojedapo等[21]指出垫料平养下3个品系商品肉鸡的体重差异显著;Shelton等[22]考察了不同能量、蛋白质水平下2种肉鸡的生产性能,结果显示2种肉鸡生产性能差异显著。本试验不同遗传品系间28、49和67 d体重均差异极显著,其中28和67 d体重以品系SD02最高,49 d体重以品系SD0302最高,品系 SD02次之;1~28 d、1~49 d和 1~67 d体增重差异均极显著,且均以品系SD02最高;1~28 d采食量差异极显著,其中品系SD0302和SD02极显著高于品系SD03和SD0203,而1~49 d采食量差异接近显著水平(P=0.052),1~67 d采食量差异显著,均以品系SD0302最高;表明4个品系间生产性能存在差异,以品系SD02和SD0302较好。
Ojedapo等[21]报道3种商品肉鸡屠宰率、胸肌重、腿肌重、肌胃重差异均极显著。本试验不同遗传品系间屠宰率差异显著,腹脂率差异极显著,其中屠宰率以品系SD0203最高,品系SD02最低,腹脂率以品系SD0302最高,品系SD02最低。这表明不同品种间差异更明显,同一品种不同遗传品系间的屠宰性能也存在差异。因此可以通过不同的杂交方式筛选最佳屠宰性能的品系,以期发挥山地鸡更大的屠宰性能。
本试验中不同遗传品系间胸肌肉色a*、b*和L*值以及屠宰后45 min pH均无显著差异,表明山地鸡肉品质遗传性能较稳定,杂交后的2个品系也未出现显著差异。不同遗传品系间山地鸡28 d各项血清生化指标差异均不显著,说明本试验中4个品系山地鸡的蛋白质代谢、氨基酸合成和脂质代谢方面无显著差异。
3.3 性别对山地鸡生产性能、屠宰性能、肉品质及血清生化指标的影响
本试验中公母鸡各阶段体重、体增重、采食量、料重比、左胸肌率、左腿肌率和腹脂率差异均极显著,公鸡的各项生产性能指标均优于母鸡,公鸡左腿肌率较高,而母鸡左胸肌率和腹脂率较高,这与前人研究报道[6,8,15,21-23]结果一致。公鸡28 d血清TP、TG和TC含量显著高于母鸡,说明公鸡在1~28 d蛋白质代谢和脂类代谢均好于母鸡。鸡的不同性别间消化生理、躯体结构存在一定差异,因此对生产性能和屠宰性能有显著性影响。在生产实践中,我们应该注意这种差异,对公母鸡进行分群饲养。
3.4 饲粮策略、遗传品系及性别的互作效应
饲粮营养水平、遗传品系、性别间的交互效应均会影响肉鸡生产性能和屠宰性能,这在前人研究(Han 等[6]、Ojedapo 等[21]、Shelton 等[22])中均有报道。饲粮策略、遗传品系和性别在67 d体重上有极显著的交互效应,其中品系SD02公鸡高赖氨酸饲粮组最高,品系SD03母鸡低赖氨酸饲粮组最低;饲粮策略、品系和性别在1~67 d体增重上有极显著的交互效应,以品系SD02公鸡高赖氨酸饲粮组最高,品系SD03母鸡低赖氨酸饲粮组最低,说明4个品系的公母鸡对2种饲粮策略表现出不同的响应,这与Dozier等[23]结果不完全一致。因此我们在生产实践中,要根据不同遗传品系、不同性别的需要,提供适宜的饲粮策略,以期发挥更好的生产性能。而饲粮策略、遗传品系和性别在各项屠宰性能、肉品质、血清生化指标上均无显著的互作效应。
4 结论
①高赖氨酸饲粮可显著改善二郎山山地鸡67 d时体重、1~67 d体增重、1~49 d和1~67 d料重比,但对其他生产性能和屠宰性能指标无显著影响。
②4个遗传品系的体重、体增重、采食量、屠宰率和腹脂率存在显著差异,而料重比等无显著差异。总体上优劣顺序为:品系SD0302、SD02>品系SD03>品系SD0203。
③各遗传品系公鸡的生产性能均优于母鸡;饲粮策略、性别和遗传品系在体增重上有显著的互作效应,应区别饲养。
[1]吕铭翰,张克英,丁雪梅,等.二郎山山地鸡0~28 d日龄适宜能量蛋白水平研究[D].硕士学位论文.雅安:四川农业大学,2010.
[2]张效先,张克英,丁雪梅,等.52~75 d日龄二郎山山地鸡饲粮适宜代谢能和粗蛋白质水平的研究[J].动物营养学报,2010,22(5):1257-1264.
[3]伍剑,张克英,丁雪梅,等.饲粮能量水平和维生素预混料对二郎山山地鸡生产性能和肉品质的影响[J].中国畜牧杂志,2011,47(19):48-52,73.
[4]周俊,宋代军.赖氨酸营养研究进展[J].饲料工业,2006,27(8):48-50.
[5]HAN Y M,BAKER D H.Lysine requirements of fastand slow-growing broiler chicks[J].Poultry Science,1991,70:2108-2114.
[6]HAN Y,BAKER D H.Effects of sex,heat stress,body weight,and genetic strain on the dietary lysinerequirement of broiler chicks[J].Poultry Science,1993,72:701-708.
[7]VASQUEZ M,PEST G M.Estimation of the lysine requirement of broiler chicks for maximum body gain and feed efficiency[J].Journal of Applied Poultry Research,1997,6:241-246.
[8]KIDD M T,FANCHER B I.Lysine needs of starting chicks and subsequent effects during the growing Period[J].Journal of Applied Poultry Research,2001,10:385-393.
[9]杨宁,李辉,王宝维.家禽生产学[M].北京:中国农业出版社,2002.
[10]赵丽红,马秋刚,陈旭东,等.日粮代谢能和赖氨酸水平对AA肉鸡生长性能和屠宰性能的影响[J].中国畜牧杂志,2008,44(23):35-40.
[11]吕明斌,孙作为,李勇,等.代谢能和可消化赖氨酸水平及互作效应对樱桃谷肉鸭生产性能和胴体指标的影响[J].中国饲料,2007(20):11-14.
[12]刘升军,呙于明.日粮蛋氨酸及赖氨酸水平对雌性肉仔鸡胴体组成的影响[J].中国畜牧杂志,2001,37(2):5-8.
[13]石天虹,黄保华,刘辉,等.日粮蛋氨酸、赖氨酸、苏氨酸对0~12周龄乌骨鸡生产性能的影响[J].山东农业科学,2005(5):52-55.
[14]陈春梅,宋遥,唐茂妍,等.日粮蛋白质和赖氨酸水平对AA肉鸡生长性能及肌肉品质的影响[J].中国农业大学学报,2006,11(6):55-59.
[15]KIDD M T,KERR B J,HALPIN K M,et al.Lysine levels in starter and grower-finisher diets affect broiler performance and carcass traits[J].Journal of Applied Poultry Research,1998,7:351-358.
[16]LATSHAW J D.Dietary lysine concentrations from deficient to excessive and theeffects on broiler chicks[J].British Poultry Science,1993,34:951-958.
[17]CORZO A,KIDD M T.Starter dietary lysine level and strain cross effects on performance and carcass traits of broiler females[J].Brazilian Journal of Poultry Science,2004,6:93-97.
[18]BERRI C,BESNARD J,RELANDEAU C.Increasing dietary lysine increases final PH and decreases drip loss of broiler breast meat[J].Poultry Science,2008,87:480-484.
[19]BERRY C,WACRENIER N,MILLET N,et al.Effect of selection for improved body composition on muscle and meat characteristics of broilers from experimental and commercial lines[J].Poultry Science,2001,80:833-838.
[20]陈志敏,蔡辉益,于会民,等.不同赖氨酸添加水平对肉仔鸡血液生理生化指标的影响[J].黑龙江畜牧兽医,2006(11):68-69.
[21]OJEDAPO L O,AKINOKUN O,ADEDEJI T A,et al.Effect of strain and sex on carcass characteristics of three commercial broilers reared in deep litter system in the derived savannah area of nigeria[J].World Journal of Agricultural Sciences,2008,4(4):487-491.
[22]SHELTON J L,MAVROMICHALIS I,PAYNE R L,et al.Growth performance of different breed crosses of chicks fed diets with different protein and energy sources[J].Poultry Science,2003,82:272-278.
[23]DOZIER W A Ⅲ,CORZO A,KIDD M T,et al.Digestible lysine requirements of male and female broilers from fourteen to twenty-eight days of age[J].Poultry Science,2009,88:1676-1682.
猜你喜欢
青海草业(2022年2期)2022-07-23
园林科技(2020年2期)2020-01-18
小哥白尼(军事科学)(2019年5期)2019-08-27
山东林业科技(2019年2期)2019-06-03
乡村地理(2018年3期)2018-11-06
乡村地理(2018年4期)2018-03-23
广东饲料(2016年3期)2016-12-01
广西林业科学(2016年2期)2016-03-20
应用化工(2014年10期)2014-08-16
作物研究(2014年6期)2014-03-01
