
饲粮代谢能水平对北京油鸡屠宰性能和肌内脂肪含量的影响
2013-09-20 03:05陈继兰李冬立岳文斌郑麦青赵桂苹
动物营养学报 2013年10期
白 洁 陈继兰 李冬立 岳文斌 郑麦青 赵桂苹 文 杰
(1.中国农业科学院北京畜牧兽医研究所,动物营养学国家重点实验室,北京 100193;2.山西农业大学动物科技学院,太谷 030801)
北京油鸡是我国著名优良地方品种之一,具有肉质细嫩、肌内脂肪(intramuscular fat,IMF)含量高等优点。现代优质肉鸡的生产不仅注重生长速度和饲料的利用效率,同样注重上市日龄的屠体性状和肉质风味。因此,提高北京油鸡的屠体性能和IMF含量,加强肉品风味成为营养与遗传研究的热点。饲粮能量对家禽有多种营养代谢作用,能量的摄入量直接影响肉鸡的胴体组成和机体脂肪含量[1-2]。饲粮能量能够影响肌肉蛋白质分解能力、脂肪沉积,改变家禽屠宰率及肌肉风味等肌肉品质指标[3-4],但目前尚缺乏北京油鸡代谢能需要的推荐量。
IMF是影响肉品质量和风味的重要指标之一。IMF的含量与肉的嫩度和多汁性有高度的相关性[5-6],是影响肉质的一项重要经济性状。哺乳动物上的研究发现,脂肪型脂肪酸结合蛋白(adipocyte fatty acid-binding protein,A-FABP)基因型与IMF含量存在高度相关性[7-8]。家禽A-FABP基因多态性位点产生的不同基因型肉鸡肌肉的脂肪性状有显著差异;鸡A-FABP基因可作为影响鸡IMF含量的候选基因[9]。北京油鸡鸡群中存在A-FABP基因P4位点的突变,突变型(NN)北京油鸡IMF含量显著高于未突变型(MM)和杂合型(MN)北京油鸡[10-11]。饲粮代谢能水平可影响肉鸡的腹脂沉积和IMF含量,遗传选育IMF含量高的A-FABP基因型突变个体,其表型可能受饲粮代谢能水平影响[2-4]。为此,本研究旨在探讨饲粮代谢能水平与A-FABP基因型对北京油鸡屠宰性能和IMF含量影响的交互效应,为不同基因型北京油鸡的饲粮适宜代谢能水平提供科学依据。
1 材料与方法
1.1 试验设计与试验动物
采用单因素随机区组试验设计,选用健康体重相近[(665.6±20.1)g]的42日龄北京油鸡公鸡(由中国农业科学院畜牧研究所家禽遗传育种研究室提供)480只,随机分成4组,分别饲喂代谢能为11.76、12.39、13.02 和13.65 MJ/kg 的饲粮(Ⅰ~Ⅳ组),每组6个重复,每个重复20只鸡。试验期49 d。试验饲粮参照我国《鸡饲养标准》(NY/T 33—2004)中的黄羽肉鸡营养推荐量配制,其组成及营养水平见表1,除代谢能外各组所有营养水平保持一致。

表1 试验饲粮组成及营养水平(风干基础)Table 1 Composition and nutrient levels of experimental diets(air-dry basis) %
1.2 饲养管理
饲养试验在中国农业科学院北京畜牧兽医研究所北京油鸡育种场进行,试验鸡采用3层阶梯式金属笼笼养,每个笼4只鸡,相邻的5个笼为1个重复。全期自由采食、饮水。饲养管理、光照、消毒和免疫按常规方法进行。试验期间每日观察试验鸡只的精神状态,记录耗料量。
1.3 样品采集
于试验结束(90日龄)后断料8 h称重,并称取剩料重;每只鸡单独标号,称量体重;每个重复随机选取7只鸡,翅静脉采血10 mL,用肝素钠抗凝,以备分析鸡只A-FABP基因多态性;采血后,将鸡只宰杀,立刻取心脏和腹部脂肪组织,于液氮中保存,以备分别分析心脏型脂肪酸结合蛋白(H-FABP)和A-FABP基因mRNA表达水平;测定每只鸡的屠宰性能指标,分析胸肌和腿肌IMF的含量。
1.4 测定指标及方法
1.4.1 屠宰性能指标测定
试验结束时将所有鸡只宰杀,称量屠体重、全净膛重、胸肌重、腿肌重和腹脂重,计算屠体率、全净膛率、胸肌率、腿肌率和腹脂率。具体计算公式如下:

1.4.2 A-FABP基因型分析
利用PCR-单链构像多态性分析(PCR-single strand conformation polymorphism,SSCP)对A-FABP基因进行多态性分析。取30μL鸡全血用于鸡全基因组DNA的提取,操作参考文献[12]的方法。根据叶满红等[9]对北京油鸡P4位点C→T的突变发现,对A-FABP基因(AF 432506)采用Primer Premier 5.0软件进行引物设计,A-FABP上、下游引物分别为 A-FABP-F:5'-CAGCTGTGTCCAGCTTGGTA-3'和 A-FABP-R:5'-AAGCAATTTGTCCCTTGGTG-3'。对鸡基因组 DNA进行PCR扩增,PCR扩增12μL反应体系:基因组DNA 溶 液 1 μL(约 50 ng),A-FABP-F(10 mmol/L)0.3 μL,A-FABP-R(10 mmol/L)0.3μL,10× 缓冲液 1.2 μL,Mg2+(25 mmol/L)1.2 μL,dNTP(2.5 mmol/L)0.24 μL,Taq 酶(2.5 U/μL)0.8 μL,ddH2O 6.96 μL。A-FABP基因扩增条件为:95℃ 预变性5 min;95℃变性30 s,59.3 ℃退火30 s,72 ℃延伸30 s,30 个循环;72℃延伸7 min;4℃保存。将PCR产物在用2%的琼脂糖凝胶检测PCR产物大小,并进行回收和测序,验证引物的特异性。
取1μL PCR产物置于PCR管中,加5μL变性缓冲液[98%去离子甲酰胺,0.01 mol/L乙二胺四乙酸(ethylene diamine tetraacetic acid,EDTA)(pH 8.0),0.025%溴酚蓝,0.025%二甲苯青,20μL/mL甘油],离心混匀后于PCR仪上98℃变性10 min,经14%非变性聚丙烯酰胺凝胶120 V,4℃电泳14~16 h后,银染显带,拍照保存,以进行分析。
1.4.3 胸肌和腿肌IMF含量测定
取部分胸肌和腿肌样品,去除可见脂肪,鲜样粉碎后,烘干;采用索氏浸提法分别测定胸肌和腿肌IMF含量。

1.4.4 A-FABP和H-FABP基因mRNA表达水平测定
采用TRIzol一步法分别对约100 mg腹脂或心肌样品进行总RNA提取;利用琼脂糖凝胶电泳检测RNA的完整性,并通过分光光度计测定总RNA的浓度。将mRNA反转录成cDNA第1链,反转录反应体系为25μL:总RNA 2μL(400 ng),Oligo dT18(25 pmol/μL)2 μL, dNTP(10 mmol/L)1.25μL,DEPC 处理水 12.75μL,5×实时荧光定量PCR缓冲液5.0μL,RNA酶抑制剂1.0μL,M-MLVRT(200 U/μL)反转录酶1.0μL。反转录条件为:70℃ 5 min,冰浴后,迅速加入反转录酶后42℃温浴 30 min,72℃15 min;反应产物存放于-20℃冰箱保存。
实时荧光定量PCR反应体系为15μL,包括上游引物(Primer-F,10μmol/L)0.5 μL,下游引物(Primer-R,10 μmol/L)0.5 μL,TaqMan Mix 7.5μL(美国应用生物有限公司),ddH2O 5.0μL;实时荧光定量PCR所用基因及序列、产物大小及位置见表2。反应条件为:50℃ 2 min;95℃ 10 min;95℃ 20 s,60℃ 1 min,40个循环;4℃保存。根据PCR反应的动力曲线图、线性图谱和半对数图谱选择适当的Ct值和域值,以得到理想的标准曲线图。各样品的Ct值与标准曲线进行比较得出其起始即为目标基因的相对表达水平。
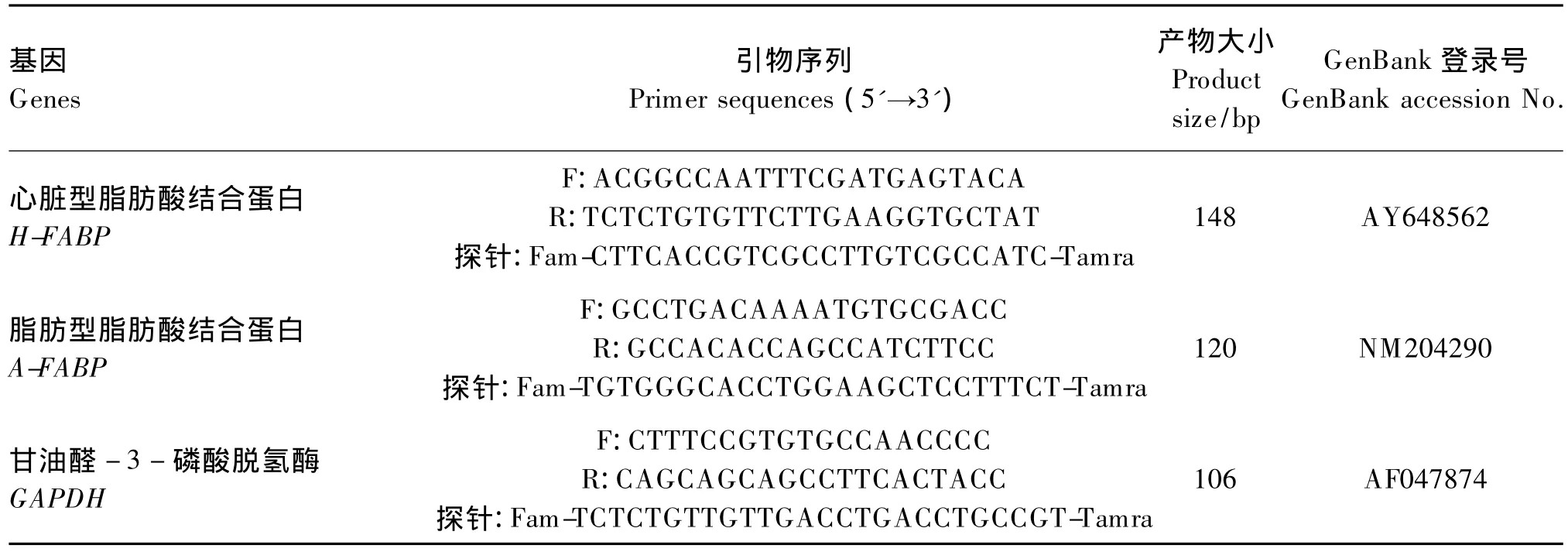
表2 实时荧光定量PCR所用基因的引物序列、产物大小及位置Table 2 The primer sequences,product size and position of the selected genes for the RT-qPCR
1.5 数据统计分析
所有数据均采用SAS 9.1.3统计软件分析;对单独代谢能效应的数据进行单因素方差分析(one-way ANOVA);对A-FABP基因型和基因频率采用NPAR1WAY程序进行卡方(χ2)检验;对屠宰性能、肉品质和基因mRNA表达水平采用GLM程序两因素的方差分析,模型采用Yij=μ+Ei+Gj+EGij+εij[式中:E 为代谢能水平(i=1,2,3,4),固定效应;G 为 A-FABP 基因型(j=1,2,3),固定效应;ε为残差],以每只鸡作为1个统计单元。方差分析显著者,利用Tukey法进行多重比较。对基因mRNA表达水平与屠宰性能指标间的相关性,采用Pearson相关分析。结果以平均值±标准差(mean±SD)表示,以P<0.05作为差异显著性判断标准。
2 结果与分析
2.1 饲粮代谢能水平对北京油鸡屠宰性能和肌肉IMF含量的影响
由表3可见,饲粮代谢能水平在不同程度上影响了北京油鸡的活体重、屠体率、全净膛率,但对腿肌率、胸肌率、腹脂率、腿肌和胸肌IMF含量无显著影响(P>0.05)。其中,Ⅲ组和Ⅳ组的活体重、屠体率和全净膛率显著高于Ⅰ组和Ⅱ组(P<0.05);Ⅲ组和Ⅳ组之间及Ⅰ组和Ⅱ组之间均差异不显著(P>0.05)。与其他3个代谢能水平相比,高代谢能(13.65 MJ/kg)有增加北京油鸡腹脂率的趋势(P<0.10)。此结果表明,北京油鸡饲粮代谢能提高到13.02 MJ/kg时可提高北京油鸡的活重、屠体率和全净膛率,但对胸肌率和腹脂率等其他屠宰性能指标无显著影响;而代谢能提高到13.65 MJ/kg时对活体重和屠体性能指标无进一步的改善作用,但有提高腹脂率的趋势。
2.2 A-FABP基因各基因型及等位基因频率的分析
本试验PCR-SSCP的结果证实,在A-FABP基因外显子1第51位碱基(P4位点)存在碱基C→T的突变。据单基因突变形成了 NN、MM和MN 3种基因型。由表4可见,经χ2适合性检验表明,选取的各代谢能组北京油鸡中A-FABP基因3种基因型在组内以及各组间的基因型频率和等位基因频率无显著差异(P>0.05),提示各组A-FABP基因的基因型分布平衡。
2.3 饲粮代谢能水平和基因型对北京油鸡屠宰性能的影响
由表5可见,北京油鸡活体重和屠体重受饲粮代谢能水平的显著影响(P<0.05)。基因型显著影响北京油鸡全净膛率(P<0.05),其中A-FABP基因MM和NN全净膛率均高于MN(P<0.05)。饲粮代谢能水平和A-FABP基因型均显著影响了腹脂率(P<0.05),但对胸肌率和腿肌率无显著影响(P>0.05),其中13.65 MJ/kg代谢能组北京油鸡腹脂率分别是11.76、12.39、13.02 MJ/kg代谢能组的1.65、2.43和2.50倍,差异显著(P<0.05);NN显著高于MM和MN(P< 0.05)。 与 中 等 代 谢 能 (12.39、13.02 MJ/kg)组相比,高代谢能(13.65 MJ/kg)能增加NN北京油鸡的腹脂率,但对MM和MN无显著影响(P>0.05);中等代谢能时,3种基因型北京油鸡腹脂率无显著差异(P>0.05)。由此可见,饲粮代谢能对北京油鸡活体重和屠体重的影响与不考虑A-FABP基因型的结果一致。
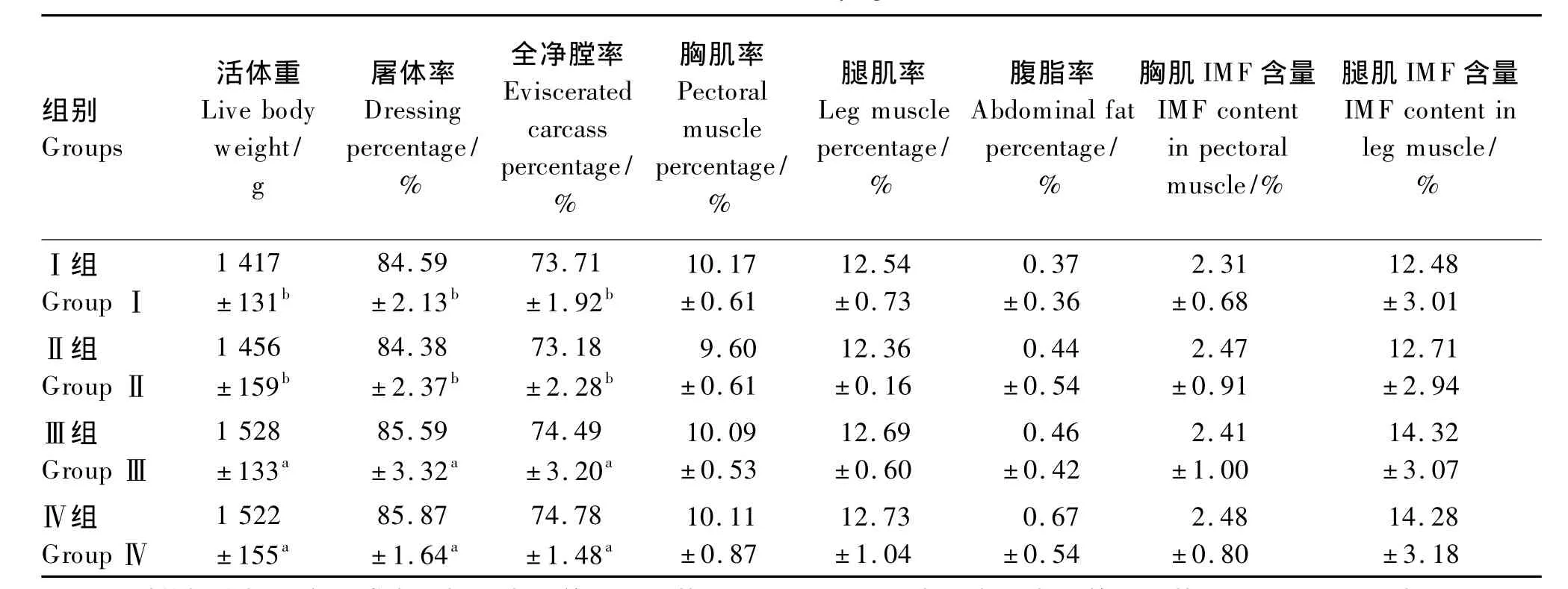
表3 饲粮代谢能水平对北京油鸡屠宰性能和肌肉IMF含量的影响Table 3 Effects of dietary metabolizable energy level on carcass performance and muscle IMF content in Beijing You chickens

表4 不同代谢能水平组A-FABP基因型频率和等位基因频率分布Table 4 The genotype and allele frequencies of A-FABP gene in different metabolizable energy groups
2.4 饲粮代谢能水平和基因型对北京油鸡肌肉IMF含量和FABP基因mRNA表达水平的影响
由表6可见,饲粮代谢能水平显著影响了胸肌IMF含量(P<0.05),但A-FABP基因型和代谢能与基因型的互作对胸肌IMF含量无显著影响(P>0.05)。12.39 MJ/kg代谢能组北京油鸡胸肌IMF含量分别是11.76、13.02和13.65 MJ/kg代谢能组的1.35、1.34和1.32倍,差异显著(P<0.05)。基因型显著影响了腿肌IMF含量和脂肪组织中A-FABP基因mRNA表达水平(P<0.05),其中MM 显著高于MN和NN(P<0.05)。饲粮代谢能水平、基因型和二者的互作对H-FABP基因mRNA表达水平无显著影响(P>0.05)。

表5 饲粮代谢能水平和A-FABP基因型对北京油鸡体重和屠宰性能的影响Table 5 Effects of dietary metabolizable energy level and genotypes of A-FABP gene on body weight and carcass performance in Beijing You Chickens
2.5 H-FABP、A-FABP基因mRNA表达水平与相关性状的相关分析
由表7可见,固定代谢能水平效应,A-FABP基因mRNA表达水平与腿肌IMF含量和基因型均存在显著相关性(P<0.01),相关系数分别为0.411 4和0.377 4;但H-FABP基因mRNA表达水平与各指标间的相关性均未达到显著水平(P>0.05)。
3 讨论
3.1 饲粮代谢能水平对北京油鸡体重和屠宰性能的影响
本试验的结果表明,饲粮代谢能低于13.02 MJ/kg时,北京油鸡活体重随着饲粮代谢能升高而增大;当代谢能大于13.02 MJ/kg时,活体重即达到平台期。快大型肉鸡的研究表明,肉鸡增重与饲粮代谢能水平间呈二次曲线关系;在一定范围内肉鸡增重随着饲粮代谢能水平的升高而升高[13-14]。刘蒙等[15]和周桂莲等[16]在地方优质肉鸡上的研究也得到相似的结果。但也有研究表明,饲粮的能量水平对地方慢速生长肉鸡体重无显著影响[17-18]。家禽具有为能而食的特点,能通过采食量和饲料转化效率调节平衡能量摄入[2-3,19-20]。肉鸡饲粮能量浓度超出了采食量的调节范围,过低或过高的能量水平均能降低肉鸡的体重,造成养殖效益的降低[2-3,19]。在三黄鸡肉种鸡的育成前期,低能量饲粮(10.5~11.7 MJ/kg)可提高肉鸡的生产性能,而在育成后期,随着饲粮能量水平的增加(10.5~12.5 MJ/kg),肉鸡增重和饲料转化率显著提高[21]。陈继兰等[22]对石岐黄鸡的研究表明,随着饲粮代谢能水平的增加(11.3~13.2 MJ/kg),试验鸡的平均日增重和饲料效率显著提高。因此,不同品种、不同生长阶段的肉鸡饲粮的适宜能量水平也有所差异。我国《鸡饲养标准》(NY/T 33—2004)中给出黄羽肉种鸡代谢能需要量为11.70 MJ/kg。本试验结果表明,增加饲粮代谢能水平到13.02 MJ/kg,提高了北京油鸡的活体重,而继续增加饲粮代谢能水平其活体重不再增加,这说明7~13周龄北京油鸡公鸡适宜的饲粮代谢能水平为13.02 MJ/kg。

表6 饲粮代谢能水平和A-FABP基因型对北京油鸡肌肉IMF含量和FABP基因mRNA表达水平的影响Table 6 Effects of dietary metabolizable energy level and genotypes of A-FABP gene on muscle IMF content and FABP gene mRNA expression level of Beijing You Chickens

表7 H-FABP、A-FABP基因mRNA表达水平与屠宰性能相关指标的相关分析Table 7 The correlation analysis between H-FABP or A-FABP gene mRNA expression level and related parameters of carcass performance in Beijing You chickens
快大型肉仔鸡[2]、中速型黄羽肉鸡[23]和慢速型石岐杂肉鸡[17]上的研究表明,高能(代谢能>13.00 MJ/kg)饲粮显著提高了肉鸡的屠体重、全净膛重、腹脂重等屠宰性能的绝对指标。Zhao等[24]在北京油鸡上的研究结果也表明,随着饲粮代谢能水平的升高,屠体率、全净膛率和腹脂率显著升高。本试验结果与上述研究报道相一致。但Dozier等[25]发现增加代谢能水平并未显著增加快大型肉鸡屠体重和胸肌重;Skinner等[1]研究发现,增加肉鸡饲粮代谢能水平显著降低了腹脂率。这可能是由于高能饲粮降低了肉鸡的采食量,使氨基酸摄入不足造成的[4]。刘蒙等[15]发现提高饲粮中代谢能水平能显著提高北京油鸡胸肌和腿肌中IMF含量。Summers等[4]在保持蛋能比与氨基酸和能量比的情况下提高代谢能,发现高能饲粮对腹脂率无显著影响,但增加了屠体重的脂肪沉积。本研究发现提高饲粮代谢能水平显著提高了腹脂率和胸肌中IMF含量,但未显著提高腿肌中的IMF含量,与上述研究不完全一致。蛋氨酸和赖氨酸来源的高能饲粮能显著增加脂肪在肌肉组织中的沉积,而未显著增加其在腹脂中的沉积;脂肪等能量饲料来源的高能饲粮能显著增加腹脂沉积[1,4,26]。故饲粮能量来源及与蛋白质的比例不同可能是造成了以上试验结果差异的原因。
3.2 A-FABP基因型对北京油鸡屠宰性能和肌肉IMF含量的影响
本实验室前期研究表明,北京油鸡A-FABP基因第1个外显子上第51位编码碱基处C→T突变形成的不同基因型与肌肉IMF含量、腹脂率等相关性状有显著的关联性[9]。Wang 等[26]也发现A-FABP基因型与鸡屠体重、腹脂率有高度相关。A-FABP基因型可以遗传选育肉鸡脂肪性状的主效基因或与主效基因连锁的候选基因[9,27]。本试验也表明,NN北京油鸡腹脂率显著高于MM和MN,与叶满红等[9]研究结果一致。但本试验中NN未显著增加胸肌中的IMF含量,而叶满红等[9]发现NN能显著增加胸肌中IMF含量。试验群体中基因型的频率是影响基因多态性与性状关联可靠性的一个重要因素,群体中A-FABP等位基因纯合型频率偏低会影响基因型与性状关联性分析的准确性[28]。叶满红等[9]采集北京油鸡 AFABP基因未突变型与突变型频率相似(30%),而本试验中A-FABP基因未突变型频率(16%)偏低;这可能是造成2个试验结果存在差异的原因。因此,阐明A-FABP基因型对北京油鸡肌肉IMF含量的影响需要增加等位基因纯合型样本含量展开进一步的研究。脂肪酸结合蛋白在机体中参与细胞内脂肪酸的摄取,可协助将脂肪酸运至其进行β-氧化的场所(如线粒体或过氧化物酶体),促进甘油三酯和磷脂的合成[26]。A-FABP作为FABPs的家族成员,其基因只在脂肪细胞中表达[29]。A-FABP被认为是组织脂类累积的标示物[30]。A-FABP基因mRNA表达水平升高可显著增加细胞内脂肪的分解,降低脂肪的合成[29-30]。本试验发现,MM A-FABP基因显著提高了A-FABP基因mRNA表达水平,这可能是MM北京油鸡腹脂率显著低于NN的原因。本试验还发现,A-FABP基因mRNA表达水平与腿肌IMF含量、A-FABP基因型显著相关;MM A-FABP基因mRNA表达水平和腿肌IMF含量显著高于MN和NN。A-FABP基因可促进脂肪沉积,并参与IMF的生成[1,4],但刘蒙等[15]研究表明 IMF 受多基因调控,并可能受多基因的连锁效应影响。因此,将A-FABP基因作为影响畜禽肉质性状的候选基因并应用于选育还需做大量研究。
3.3 饲粮代谢能水平与基因型互作对北京油鸡腹脂率的影响
肉鸡腹脂增加是脂肪细胞的增殖或细胞内甘油三酯的累积造成的[31]。本试验发现,与中等代谢能(12.39和13.02 MJ/kg)相比,饲粮高代谢能(13.65 MJ/kg)水平显著增加了NN北京油鸡的腹脂率,但对MM和MN北京油鸡无显著影响;A-FABP基因型与腹脂率具有高度相关,但饲粮代谢能水平与腹脂率无显著的相关性。肉鸡腹脂的增加能显著增加A-FABP基因mRNA表达水平[30],但本试验中NN北京油鸡腹脂率显著高于MM和MN,而NN北京油鸡A-FABP基因mRNA表达水平却显著低于MM,与MN无显著差异。A-FABP基因mRNA表达水平与蛋白质水平低度相关[32],A-FABP蛋白可能存在转录后的修饰作用,这可能是本试验中NN北京油鸡腹脂率增加而A-FABP基因mRNA表达水平未增加的原因。已有研究表明,饲粮中添加0.1%的油酸能显著降低A-FABP基因 mRNA 表达水平[33],而 A-FABP基因mRNA表达水平与长链脂肪酸的量密切相关[4]。本试验饲粮代谢能的提高主要通过增加玉米和玉米油的含量来增加代谢能浓度,饲粮中油酸含量的增加可能是本试验饲粮代谢能水平的升高降低了MM北京油鸡A-FABP基因mRNA表达水平的主要原因。
3 结论
①北京油鸡饲粮13.02 MJ/kg代谢能显著提高了北京油鸡的活体重、屠体率、全净膛率和腹脂率。
②高代谢能饲粮(13.65 MJ/kg)显著增加了A-FABP基因突变型北京油鸡的腹脂率。
③饲粮代谢能可显著影响胸肌IMF含量,基因型可显著影响腿肌IMF含量,但二者无交互作用。
[1]SKINNER J T,WALDROUP A L,WALDROUP P W.Effects of dietary nutrient density on performance and carcass quality of broilers 42 to 49 days of age[J].Journal of Applied Poultry Research,1992,1(4):367-372.
[2]DOZIER W A,PRICE C J,KIDD M T,et al.Growth performance,meat yield,and economics of broilers fed diets varying in metabolizable energy from thirty to fifty-nine days of age[J].The Journal of Applied Poultry Research,2006,15:367-382.
[3]DOZIER W A,CORZO A,KIDD M T,et al.Dietary apparent metabolizable energy and amino acid density effects on growth and carcass traits of heavy broilers[J].The Journal of Applied Poultry Research,2007,16:192-205.
[4]SUMMERS J D,SPRATT D,ATKINSON J L.Broiler weight gain and carcass composition when fed diets varying in amino acid balance dietary energy and protein level[J].Poultry Science,1992,71(2):263-273.
[5]WOOD J D,ENSER M,MONCRIEF C C,et al.Effects of carcass fatness and sex on the composition and quality of pig meat[C]//The 34th international congress of meat science and technology.Brisbane:[s.n.],1988:562-564.
[6]孙玉民,罗明.畜禽肉品学[M].济南:山东科学技术出版社,1993:25-48.
[7]GERBENS F,JANSEN A,VAN ERP A J,et al.The adipocyte fatty acid-binding protein locus:characterization and association with intramuscular fat content in pigs[J].Mammalian Genome,1998,9(12):1022-1026.
[8]GERBENS F,VERBURG F J,VAN MOERKERK H T,et al.Associations of heart and adipocyte fatty acidbinding protein gene expression with intramuscular fat content in pigs[J].Jounral of Animal Nutrition,2001,79:347-354.
[9]叶满红,文杰,曹红鹤,等.脂肪型脂肪酸结合蛋白基因多态性与鸡肉品质性状的关系研究[J].畜牧兽医学报,2007,38(6):526-532.
[10]罗桂芬,陈继兰,文杰,等.鸡A-FABP基因多态性分析及其与脂肪性状的相关研究[J].遗传,2006,28(1):39-42.
[11]李文娟,李宏宾,文杰,等.鸡H-FABP和A-FABP基因表达与肌内脂肪含量相关研究[J].畜牧兽医学报,2006,37(5):417-423.
[12]萨姆布鲁克J,弗里奇E F,曼尼阿蒂斯T.分子克隆实验指南[M].2版.金冬雁,里猛,黎孟枫,等译.北京:科学出版社,1992:453-456.
[13]FARRELL D J.Effects of dietary energy concentration on utilisation of energy by broiler chickens and on body composition determinted by carcass analysis andpredicted using tritium[J].British Poultry Science,1974,15(1):25-41.
[14]WALDROUP P W,MITCHELL R J,PAYNE J R,et al.Characterization of the response of broiler chickens to diets varying in nutrient density content[J].Poultry Science,1976,55(1):130-145.
[15]刘蒙,文杰,宋代军,等.日粮能量水平与肌内脂肪遗传选择对北京油鸡生产性能的互作影响[J].安徽农业大学学报,2009,36(3):456-460.
[16]周桂莲,蒋宗勇,林映才,等.22~42日龄黄羽肉鸡饲粮代谢能需求参数的研究[J].动物营养学报,2004,16(1):57-64.
[17]何翔,高振华,孔鹏,等.不同能量和粗蛋白水平日粮对石岐杂鸡生产性能、屠宰性能及养分表观代谢率的影响[J].河南农业科学,2011,40(12):153-156.
[18]蒋桂韬,戴求仲,胡艳.能量和蛋白水平对湘黄鸡生长性能及胴体品质的影响[J].湖南畜牧兽医,2004(5):6-8.
[19]HIDALGO M A,DOZIER W A Ⅲ,DAVIS A J,et al.Live performance and meat yield response of broilers to progressive concentrations of dietary energy maintained at a constant metabolizable energy-to-crude protein ratio[J].Journal of Application Poultry Research,2004,13:319-327.
[20]LESSON S,CASTON L,SUMMER J D.Broiler response to diet energy[J].Poultry Science,1996,75(4):529-535.
[21]鞠科,肖从兴.日粮不同能量水平对广西三黄肉鸡种鸡育成期的生产性能和血液生化指标影响[J].畜牧与饲料科学,2009,30(10):25-26.
[22]陈继兰,吕连山,赵玲,等.石岐黄肉鸡前期日粮适宜的能量和蛋白质水平的研究[J].中国畜牧杂志,1998,34(4):10-12.
[23]蔺淑琴,李金录,史兆国,等.日粮不同营养水平对黄羽肉鸡屠宰性能及肉品质的影响[J].中国畜牧兽医,2000,35(8):9-12.
[24]ZHAO J P,CHEN J L,ZHAO G P,et al.Live performance,carcass composition,and blood metabolite responses to dietary nutrient density in two distinct broiler breeds of male chickens[J].Poultry Science,2009,88(12):2575-2584.
[25]DOZIER W A,GEHRING C K,CORZO A,et al.Apparent metabolizable energy needs of male and female broilers from 36 to 47 days of age[J].Poultry Science,2011,90(4):804-814.
[26]WANG Q L,LI H,LI N,et al.Tissue expression and association with fatness traits of liver fatty acid-binding protein gene in chicken[J].Poultry Science,2006,85:1890-1895.
[27]WANG Q,LI H,LI N,et al.Identification of single nucleotide polymorphism of adipocyte fatty acid-binding protein gene and its association with fatness traits in the chicken[J].Poultry Science,2006,85:429-434.
[28]王启贵,关天竹,王守志,等.A-FABP基因多态性与肉鸡生长和体组成性状的关联[J].遗传,2011,33(2):153-162.
[29]VEERJAMP JH,VAN MOERKERK H T.Fatty acidbinding protein and its relation to fatty acid oxidation[J].Molecular and Cellular Biochemistry,1993,123(1/2):101-106.
[30]SHI H,WANG Q,ZHANG Q,et al.Tissue expression characterization of chicken adipocyte fatty acid-binding protein and its expression difference between fat and lean birds in abdominal fat tissue[J].Poultry Science,2010,89(2):197-202.
[31]SOUKAS A,SOCCI N D,SAATKAMP B D,et al.Distinct transcriptional profiles of adipogenesis in vivo and in vitro[J].Journal of Biology Chemistry,2001,276(36):34167-34174.
[32]WANG D,WANG N,LI N,et al.Identification of differentially expressed proteins in adipose tissue of divergently selected broilers[J].Poultry Science,2009,88(11):2285-2292.
[33]XING J,KANG L,JIANG Y.Effect of dietary betaine supplementation on lipogenesis gene expression and CpG methylation of lipoprotein lipase gene in broilers[J].Molecular Biology Reports,2011,38(3):1975-1981.
猜你喜欢
动物营养学报(2022年9期)2022-10-12
动物营养学报(2022年8期)2022-09-01
中国饲料(2021年17期)2021-11-02
家禽科学(2021年8期)2021-09-23
畜牧兽医科学(2021年21期)2021-06-27
中国食用菌(2020年12期)2020-12-18
中国饲料(2020年16期)2020-11-06
数学大王·趣味逻辑(2018年11期)2018-12-14
江苏农业科学(2017年14期)2017-10-10
广东饲料(2016年6期)2016-12-01
