
不同复种方式对水稻产量和土壤肥力的影响
2013-09-12 06:07周春火潘晓华吴建富石庆华
植物营养与肥料学报 2013年2期
周春火,潘晓华,吴建富,石庆华
(1江西农业大学国土资源与环境学院,南昌330045;2江西省作物生理生态与遗传育种重点实验室,南昌330045)
合理复种轮作是农田用地和养地相结合,提高作物产量的一项行之有效的耕作栽培措施。在长江中游双季稻区,冬闲+双季稻、紫云英+双季稻和油菜+双季稻是稻田主要的复种方式[1]。大量研究表明,不同复种轮作方式下的水稻生长发育和土壤肥力存在较大差异[2-6]。高菊生等[5]研究表明,与双季稻冬闲相比,双季稻与紫云英轮作种植可显著提高稻谷产量,提高稻田土壤有机质累积量,且土壤活性有机质组分显著高于双季稻冬闲,增加微生物碳在土壤有机碳中的比重的效果好于双季稻油菜处理。但以往的研究多将油菜作为绿肥作物种植,且双季稻生产中没有考虑冬作物带入土壤的养分,稻草也不还田。为此,本文通过2年定位试验,在稻草全量还田,并且考虑紫云英翻压还田带入和油菜收获带走的氮素养分的情况下,研究了3种复种方式(紫云英+双季稻、油菜+双季稻、冬闲+双季稻)对水稻产量及其构成和土壤肥力的影响,旨在为双季稻区合理复种提供科学依据。
1 材料与方法
1.1 试验设计与田间管理
于2009年9月开始,在江西省南昌市恒湖农场(北纬29°03、东经28°50')进行定位试验,试验地土壤属于冲积型水稻土。试验前土壤的基本理化性状为:有机质含量13.62g/kg、全氮1.38 g/kg、全磷1.036g/kg、全钾16.49g/kg、碱解氮 95.50 mg/kg、速效磷32.45 mg/kg、速效钾108.37 mg/kg、pH5.12。
试验设3个处理:1)紫云英 +早稻 +晚稻(ZDD);2)油菜+早稻+晚稻(YDD);3)冬闲+早稻+晚稻(XDD)。各处理小区面积为100 m2,4次重复。处理间筑埂隔开,以防串水串肥,稻草粉碎全量还田。供试早、晚稻品种分别为淦鑫203和五丰优T025。种植密度早稻为13.3×23.3cm,每蔸3粒谷苗;晚稻为13.3 cm×26.7cm,每蔸2粒谷苗。紫云英品种为“余江大叶”,油菜品种为“湘油5号”,紫云英于前一年晚稻收割前撒播,播种量为30 kg/hm2;油菜育苗移栽,密度97500 plant/hm2。
试验各处理施用的总氮量相同,即早、晚稻各施纯氮 180 kg/hm2、P2O572 kg/hm2和 K2O 180 kg/hm2,所用化学肥料分别为尿素、钙镁磷肥和氯化钾。在紫云英翻耕前、油菜收获前测定生物量(2010年和2011年紫云英地上部的生物量分别为1.125×104kg/hm2和4.35×104kg/hm2,油菜的生物量分别为4.76×104kg/hm2和5.38×104kg/hm2),并进行含氮量测定。XDD的施氮量为180 kg/hm2;ZDD的施氮量=180-[紫云英含氮量(%)×紫云英生物量];YDD的施氮量=180+(油菜含氮量-冬种油菜施氮量-油菜苗期含氮量)。施用的氮肥按基肥∶分蘖肥∶穗肥为5∶2∶3施用,磷、钾肥做基肥一次性施用,其中基肥在移栽前1 d施用,分蘖肥在栽后7 d施用,穗肥在2叶露尖时施用,其他大田管理措施一致。
1.2 测定项目及方法
水稻移栽后5 d开始,每隔5 d调查一次(每小区定株10蔸)总茎蘖数,直至抽穗为止。在每季水稻成熟期采用土柱法[7](取样直径20 cm)各处理取3蔸样,按0—5 cm、5—10 cm、10—20 cm 3层进行根量分析;每季水稻成熟期按5点法取耕层(0—20 cm)土样测定土壤有机质、全氮含量[8]和土壤酶活性[9];每年晚稻成熟期按5点法分0—5 cm、5—10 cm、10—20 cm三层取土壤样品,用于土壤微生物测定[10];每季水稻成熟时,每小区调查30蔸有效穗,按平均数法取5蔸进行考种;每小区实割30 m2进行产量测定。
1.3 数据处理
用Excel和SPSS软件进行数据处理和显著性检验。
2 结果与分析
2.1 不同复种方式对水稻产量及其构成因素的影响
试验结果表明(表1),2010年由于早稻分蘖期低温导致迟发,晚稻受强寒露风影响,产量明显低于2011年。处理间,无论早稻还是晚稻,冬种紫云英处理的水稻产量最高。2010年早稻处理间差异不大,晚稻 ZDD比 YDD处理高 3.5%,比 XDD高4.1%,差异显著或极显著;2011年早、晚稻均以ZDD处理最高,早稻ZDD处理极显著高于YDD,增幅为6.4%,晚稻ZDD处理极显著高于XDD,幅度为4.6%。处理间产量的差异主要是有效穗数不同所致。相关分析表明,早、晚稻产量与有效穗呈正相关(r=0.7740*)。
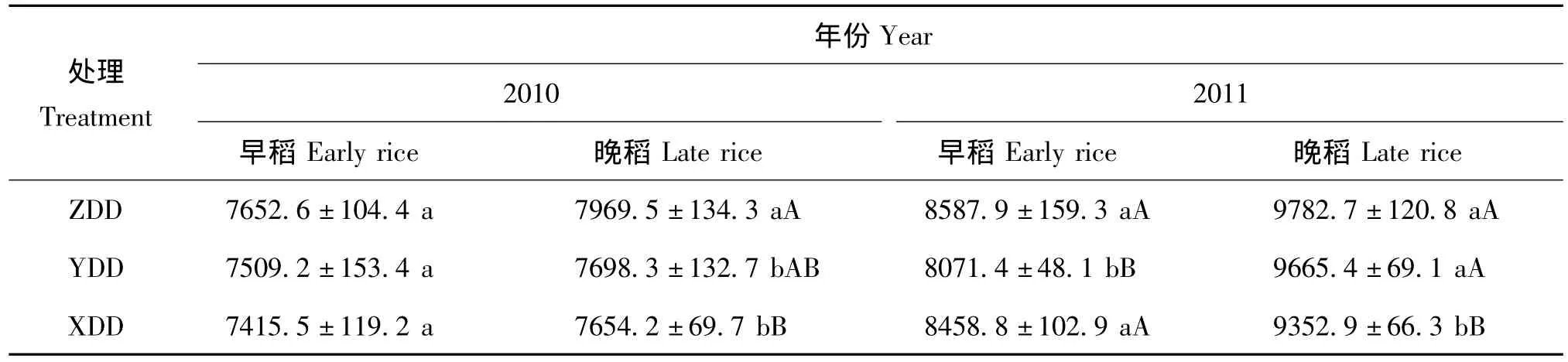
表1 不同处理对水稻产量的影响(kg/hm2)Table 1 Effect of different treatments on rice yield
从表2可以看出,2010年早、晚稻有效穗数均表现为ZDD>YDD>XDD,其中早稻处理间差异显著或极显著;早、晚稻每穗粒数呈 ZDD>YDD>XDD,晚稻ZDD与XDD处理差异显著;结实率早稻以XDD处理最高,YDD最低,差异显著或极显著,晚稻没有显著差异;早、晚稻千粒重和成穗率处理间差异不显著。2011年,早稻有效穗数ZDD处理显著高于YDD、XDD,晚稻ZDD与YDD处理相差不大,但极显著高于XDD;每穗粒数早稻以YDD处理最高,且显著高于XDD,晚稻ZDD显著高于其他处理;早、晚稻结实率、千粒重和成穗率处理间差异不显著。

表2 不同处理对水稻产量构成因素的影响Table 2 Effect of different treatment on yield components of rice
2.2不同复种方式对水稻分蘖动态的影响
由图1可以看出,年度间,各处理在2011年分蘖速率、最高茎蘖数和成穗率均高于2010年,这与两年的气候条件不同有关。
2010年早稻处理间分蘖数移栽后0~15 d差异不大,移栽25 d后分蘖数呈ZDD>YDD>XDD,分蘖达到高峰的时间ZDD处理比YDD和XDD处理晚3~5 d。2011年早稻,移栽后0~10 d处理间分蘖数几乎没有差异,随后YDD处理分蘖数增加最快,至移栽后25 d分蘖数已达最高值,而XDD和ZDD处理分别比其晚5和10 d;与XDD处理相比,ZDD处理早稻分蘖时间长、速率慢且下降快,YDD处理的早稻分蘖时间短、速度快且下降慢,但茎蘖数高和成穗率(表2)差异不大,使ZDD、YDD处理的有效穗得以提高;这主要是因为紫云英腐解使分蘖时间延长和冬种油菜收获推迟使水稻移栽时间推后的原因。
晚稻与早稻有所不同,处理间分蘖动态规律相似,最高分蘖数出现的时间相同。至移栽25 d处理间分蘖数相差不大,移栽25 d后分蘖数ZDD处理高于YDD、XDD处理。说明冬种紫云英对晚稻季生育前期生长仍有积极影响。

图1 不同处理水稻分蘖动态Fig.1 Dynamic of rice tiller under different treatments
2.3 不同复种方式对水稻根系的影响
不同处理早、晚稻成熟期根系测定结果(表3)表明:2年4季水稻根系的总干重以及在0—5 cm、5—10 cm、10—20 cm土层中的分布均呈 ZDD>YDD>XDD的趋势,且处理间差异达显著或极显著水平,说明冬种紫云英和油菜有利于水稻根系的生长。各处理水稻根系在0—5 cm和5—10 cm土层中分布较多,占根系总量的90%以上,10—20 cm土层中分布根系较少,其中早稻10—20 cm土层中分布的根系ZDD和YDD处理是XDD处理的2.1倍和1.8倍,晚稻为1.9倍和1.5倍。说明冬种紫云英和油菜不仅使各土层水稻根系总量增加,还促进水稻根系向下延伸,有利于水稻对养分的吸收。
2.4 不同复种方式对土壤有机质和全氮的影响
由图2可以看出,2年4季水稻成熟期土壤有机质含量均呈ZDD>YDD>XDD的趋势。复种1年(2季)后,与试验前相比,ZDD处理增加了0.1 g/kg,YDD 增加0.01 g/kg,而 XDD 略有减少,ZDD与XDD处理差异达显著水平;复种2年(4季)后,ZDD处理土壤有机质含量分别比YDD、XDD高0.58%、0.95%,差异显著。土壤全氮含量亦呈ZDD>YDD>XDD的趋势。复种1年后处理间差异不显著;复种2年后,与试验前相比,各处理土壤全氮含量均有提高,ZDD、YDD和XDD处理的提高幅度分别为 6.52%、2.17%和 0.72%,且 ZDD和XDD处理差异显著。可见,相对冬闲处理,冬种紫云英和油菜种植均有利于土壤有机质和全氮的积累。试验结果还可以看出,相对试验前,XDD处理土壤有机质和全氮含量变化不大,可能与稻草还田对土壤肥力的维持有关。

表3 不同复种方式对水稻根量及土层分布的影响Table 3 Effects of different multiple cropping patterns on root dry weight and distribution in soil layer of hybrid rice
2.5 不同复种方式对土壤微生物的影响
试验结果表明(表4),复种 1年后,ZDD和YDD处理0—5 cm土层中的微生物数量差异较小,但极显著高于XDD处理,增幅分别为31.5%和27.1%;5—10 cm、10—20 cm微生物呈ZDD>YDD>XDD,处理间的差异达显著或极显著水平;复种2年后,不同土层中的微生物数量ZDD处理均极显著地高于YDD和XDD,YDD和XDD两处理的微生物数量,在0—5 cm和10—20 cm土层中差异不显著,但在5—10 cm土层中差异极显著。
从不同类群微生物(表5)来看,复种1年后,土层中的细菌数量ZDD>YDD>XDD,差异达极显著水平;真菌以YDD处理最高,XDD最少,处理间差异极显著;放线菌ZDD与XDD处理差异不大,但均极显著高于YDD处理。复种2年后,土层中的细菌数量与第一年规律相同;真菌数量变化较大,以ZDD处理最高,YDD处理最少,处理间差异显著或极显著;放线菌数量XDD>ZDD>YDD,处理间达显著或极显著差异。年度间,随复种时间延长,细菌和放线菌数量各处理明显增加,这与紫云英腐解量和稻草还田有关,而真菌数量YDD处理明显减少,其原因有待进一步研究。

图2 不同复种方式对水稻成熟期土壤有机质和全氮的影响(2010~2011年)Fig.2 Effects of different multiple cropping patterns on Soil organic matter and soil total nitrogen in maturity stage of rice(2010-2011 year)

表4 不同复种方式对水稻土壤微生物的影响(×106cfu/g)Table 4 Effects of different multiple cropping patterns on soil microbial

表5 复种方式对水稻土壤微生物类群的影响Table 5 Effects of different multiple cropping patterns on soil microbes
2.6 不同复种方式对土壤酶活性的影响
土壤酶催化土壤中的一切生物化学反应,对土壤肥力有重要影响。由表6可以看出,水稻收获后,土壤中脲酶、转化酶和过氧化氢酶的活性均表现为ZDD>YDD>XDD。其中脲酶活性处理间早稻达极显著差异,且晚稻ZDD处理极显著高于YDD和XDD处理;转化酶活性处理间2年表现不同,2010年早稻ZDD处理极显著高于YDD、XDD处理,晚稻ZDD、YDD与XDD处理间差异显著;2011年早、晚差异不显著;过氧化氢酶活性处理间2年也有不同差异,2010年早稻处理间差异显著或极显著,晚稻ZDD与YDD差异不大,但ZDD极显著高于XDD。2011年早、晚稻过氧化氢酶活性与脲酶活性的规律相同。土壤酶的活性年际间有一定变化,其原因可能与年度间取样时的土壤水分、温度和养分消耗程度等因素有关。土壤酶活性的大小与土壤理化性质有关,尤其与土壤肥力的关系十分密切。相关分析表明,土壤有机质、全氮含量与土壤中脲酶、过氧化氢酶活性呈显著或极显著正相关;土壤有机质、全氮含量与转化酶活性呈正相关,但没有达到显著水平。

表6 不同复种方式对土壤酶活性变化的影响Table 6 Effects of different multiple cropping patterns on soil enzyme activity changes
3 讨论
本试验结果表明,与冬闲处理相比,ZDD处理2年4季水稻产量明显高于YDD、XDD处理,紫云英替代部分化肥能增加分蘖总量,优化穗粒结构而提高产量,其原因是ZDD处理有效穗的提高和每穗粒数的增加,而YDD和XDD处理相差不大。从养分供应过程分析,早稻生育前期紫云英腐解由于温度较低肥效缓慢,随着时间延长,土壤养分含量逐渐提高,较好满足水稻生长对养分的需求,至晚稻季仍产生后效;油菜种植消耗土壤一定养分,至水稻季虽然氮素得到补充,水稻产量并没有表现明显优势,但也没有减产,值得一提的是由于油菜生育期较长,致使早稻移栽时间推迟可能对周年产量有一定的影响,因而生产中应注意茬口安排。关于复种方式对产量的影响有较多的报道,大部分研究认为绿肥作物与双季稻轮作种植后,水稻产量显著高于冬闲对照,且以紫云英处理表现最好,但结论是在不减施化肥的条件下[4,11-13]。另有研究认为紫云英替代化肥对产量亦有促进作用,但需要控制好紫云英用量[14-15]。刘春增[13]等研究了紫云英与当地常规施肥量100%、80%、60%、40%配施,结果显示稻谷产量比仅施化肥处理分别增产4.5%、5.2%、2.8%、0.5%,紫云英还田与当地化肥施用量的80%和60%配施是最优组合,这与本文结果相似。
稻田复种轮作由于水旱交替引起土壤肥力的变化,土壤有机质是土壤的重要组成物质,与作物产量和土壤肥力密切相关。在水作条件下由于还原细菌的作用增加了有机质的积累,而在旱作时有机质分解加快。复种轮作模式中,由于冬种作物的茎秆还田,一方面提高土壤有机质的含量,使有效养分源源不断地分解释放,另一方面易氧化有机质增加,增强土壤有机质化学活性,提高土壤养分的供应强度[11]。绿肥作物紫云英为豆科植物,能够利用根瘤菌共生固氮,紫云英还田既增加土壤氮,也增加了土壤有效磷、钾和有机质,提高了土壤肥力[5]。冬种绿肥作物对土壤外源有机物料的增加,有利于增加微生物数量,这是因为有机肥本身含有一些微生物,同时有机肥刺激微生物繁殖[16-17];冬种绿肥作物在生长过程中所产生的分泌物、凋落物均增加了土壤酶的含量,同时绿肥还田引起来自土壤微生物的土壤酶活性发生改变[18]。本试验结果也表明,与冬闲处理相比,冬种紫云英和油菜提高了土壤有机质、全氮含量和微生物总量,促进了土壤微生物活动和酶活性的增加。
两年试验结果表明,在总施氮量相同的条件下,不同复种方式对水稻产量和土壤肥力有明显影响,其中以紫云英+双季稻复种方式表现最佳,对双季稻区减少化肥施用和缓解环境压力有重要意义。
[1] 黄国勤.中国南方稻田耕作制度的发展[J].耕作与栽培,2006,(3):1-5.Huang G Q.Inada Kosaku system development in South China[J].Till.Cult.,2006,(3):1-5.
[2] 陈福兴,张马祥,邹长明,等.稻田不同轮作方式对培肥地力的影响[A].中国农业科学院红壤实验站.红壤丘陵区农业发展研究[M].北京:中国农业科技出版社,1995.188-190.Cheng F X,Zhang M X,Zou C M et al.Paddy field different rotation methods on the influence of the fertilizer[A].Station of Red SoilResearch ofCAAS.Red soilhilly agricultural development research[M].Beijing:China’s Agricultural Science and Technology Press,1995.188-190.
[3] 王先华.稻田不同轮作方式对培肥地力的作用[J].耕作与栽培,2002,(6):9-10.Wang X H.Effect of the paddy-field different rotations on soil fertility[J].Till.Cultivat.,2002,(6):9-10.
[4] 高菊生,刘更另,秦道珠,等.红壤稻田不同轮作方式对水稻生长发育的影响[J].耕作与栽培,2002,(2):1-2.Gao J S,Liu G L,Qin D Z et al.Effect of red soil paddy field of different rotations on rice growth and development[J].Till.Cult.2002,(2):1-2.
[5] 高菊生,曹卫东,李冬初,等.长期双季稻绿肥轮作对水稻产量及稻田土壤有机质的影响[J].生态学报,2011,31(16):4542-4548.Gao J S,Cao W D,Li D C et al.Effects of long-term double-rice and green manure rotation on rice yield and soil organic matter in paddy field[J].Acta Ecol.Sin.,2011,31(16):4542-4548.
[6] 王辉,屠乃美.稻田种植制度研究现状与展望[J].作物研究,2006,(5):498-503.Wang H,Tu N M.Research and prospect of rice cropping system[J].Crop Res.,2006,(5):498-503.
[7] 吴朝晖,周建群,青先国.水稻根系分布形态研究法现状及展望[J].湖南农业科学,2008,(5):11-14.Wu Z H,Zhou J Q,Qing X G.Advances in methods of studying rice root system[J].Hunan Agric.Sci.,2008,(5):11-14.
[8] 史瑞和.土壤农化分析[M].北京:农业出版社,1981.Shi R H.Soil agrochemical analysis[M].Beijing:Agricultural Science,1981.
[9] 关松荫.土壤酶及其研究法[M].北京:农业出版社,1986.294-329.Guan S M.Soil enzyme and its study method[M].Beijing:Agricultural Science,1986.294-329.
[10] 陈华癸,李阜棣,陈文新.土壤微生物[M].上海:上海科学技术出版社,1981.80-232.Chen H K,Li F D,Chen W X.Soil microorganisms[M].Shanghai:Shanghai Science and Technology Press,1981.180 -232.
[11] 刘英,王允青,张祥明,等.种植紫云英对土壤肥力和水稻产量的影响[J].安徽农学通报,2007,13(1):98-99.Liu Y,Wang Y Q,Zhang X M et al.Effect of planting Astragalus sinicus on soil fertility and rice yield[J].Acta Agric.Anhui,2007,13(1):98-99.
[12] 王允青,张祥明,刘英,等.施用紫云英对水稻产量和土壤养分的影响[J].安徽农业科学,2004,(4):699-700.Wang Y Q,Zhang X M,Liu Y et al.Effect of application of Astragalus sinicus on rice yield and soil nutrient[J].Anhui Agric.Sci.,2004,(4):699-700.
[13] 刘春增,李本银,吕玉虎,等.紫云英还田对土壤肥力、水稻产量及其经济效益的影响[J].河南农业科学,2011,40(5):96-99.Liu C Z,Li B Y,Lü Y H et al.Effect of incorporation of astragalus sinicus on soil fertility,rice yield and economic efficiency[J].J.Henan Agric.Sci.,2011,40(5):96-99.
[14] 何春梅,李昱,李清华,等.紫云英还田下水稻适宜施肥量研究[J].福建农业科技,2010,(5):77-78.He C M,Li Y,Li Q H et al.Study of rice suitable fertilizer under Astragalus sinicus back to field[J].Fujian Agric.Sci.Technol.,2010,(5):77-78.
[15] 诸海焘,余廷园,田吉林,等.绿肥-水稻轮作体系中氮肥适宜用量研究[J].上海农业学报,2008,24(4):60-64.Zhu H T,Yu T Y,Tian J L et al.Study on suitable nitrogen fertilizer dose under green manure crop-rice rotation system[J].Acta Agric.Shanghai,2008,24(4):60-64.
[16] 肖嫩群,张洪霞,成壮,等.紫云英还田量对烟田土壤微生物及酶的影响[J].中国生态农业学报,2010,18(4):711-715.Xiao N Q,Zhang H X,Cheng Z.Effect of incorporation of Astragalus sinicus on microbe and enzyme dynamics in tobacco cultivated soils[J].Chin.J.Ecol.,2010,18(4):711-715.
[17] 樊军,郝明德.长期轮作与施肥对土壤主要微生物类群的影响[J].水土保持研究,2003,(1):88-89,114.Fan J,Hao M D.Effects of long-term rotations and fertilizations on soil microflora[J].Res.Soil Water Conserv.,2003,(1):88-89,114.
[18] 姜培坤,徐秋芳,周国模,等.种植绿肥对板栗林土壤养分和生物学性质的影响[J].北京林业大学学报,2007,(3):120-123.Jiang P K,Xu Q F,Zhou G M et al.Effects of green manure on soil nutrients and bio-properties of Castanea mollissima blume plantations[J].J.Beijing For.Univ.,2007,(3):120-123.
猜你喜欢
今日农业(2021年21期)2022-01-12
中国土壤与肥料(2021年5期)2021-12-02
农业资源与环境学报(2021年1期)2021-01-27
湖南农业大学学报(自然科学版)(2020年3期)2020-07-11
上海农业科技(2020年2期)2020-04-02
作物研究(2019年2期)2019-04-02
中学时代(2018年6期)2018-11-20
海峡姐妹(2018年7期)2018-07-27
作文与考试·小学高年级版(2016年14期)2016-09-10
营销界(2015年23期)2015-02-28
