
肉羊心型脂肪酸结合蛋白基因第2外显子多态性与肉质性状相关分析
2013-08-08 12:24于永生张立春郭振刚金海国
东北农业大学学报 2013年9期
曹 阳,赵 雪,,于永生,张立春,郭振刚,金海国
(1.吉林省农业科学院畜牧分院,吉林 公主岭 136100;2.延边大学农学院,吉林 延吉 133002;3.吉林农业大学动物科技学院,长春 130118)
肉品质是一个复杂经济性状,肌内脂肪是影响肉品质量和风味的重要指标之一。肌内脂肪(Intramuscular fat,IMF)含量直接影响肉品质和风味,特别是肉多汁性[1-2]。
脂肪酸结合蛋白(Fatty acid binding protein,FABP)是一族同源性较高小分子细胞内蛋白质,在心肌、小肠、肝脏、脂肪组织、脑、表皮等组织细胞中均存在,在脂类代谢过程中起重要作用。心脏型脂肪酸结合蛋白(Heart fatty acid binding pro⁃tein,H-FABP)与脂肪酸结合密切相关。目前,猪、鸡、牛、羊等畜禽H-FABP基因已经被相继克隆和定位,并有研究表明H-FABP基因可作影响IMF含量候选基因[3-7]。本试验采用PCR-SSCP方法检测杜泊×小尾寒羊H-FABP基因单核苷酸多态性,并与肉质性状进行相关分析,以期发现相关位点用于肉羊育种。
1 材料和方法
1.1 试验材料
杜泊×小尾寒羊杂交羊29只,统一标准饲养于吉林省农业科学院畜牧分院羊场,颈静脉采血9 mL左右,ACD抗凝带回实验室,用AxyPrep血基因组DNA小量制备试剂盒提取血液基因组DNA,1%琼脂糖电泳检测基因完整性及纯度,-20℃冷冻保存。
1.2 引物设计与合成
参考绵羊H-FABP基因序列(GenBank登录号:AY157617)设计H-FABP基因exon 2引物,引物序列为:F:5'AGCTCATGCTCATACCCTTC 3',R:5'CCCTCAAGGTGCAACAAT 3',扩增片段长度为285 bp。引物由上海英骏生物技术有限公司合成。
1.3 PCR扩增
PCR扩增反应体系为25 μL,包括10×PCR Buffer 1.5 μL,上、下游引物(10 pmol·L-1)各0.5 μL,dNTP(10 mmol·μL-1)0.5 μL,Taq DNA Poly⁃merase(2.5 U · μL-1)0.5 μL, DNA 模 板(50~100 ng·μL-1)1.0 μL,ddH2O 补至 25 μL。反应条件:95℃预变性5 min;94℃变性30 s,60℃退火30 s,72℃延伸30 s,35个循环;72℃延伸7 min,4℃保存,产物用1%琼脂糖凝胶电泳检测。
1.4 SSCP检测
取2 μL PCR产物,加入5 μLLoading buffer,混匀,98℃变性10 min,然后迅速冰浴10 min,变性后PCR产物用8%非变性聚丙烯酰胺凝胶(Acr:Bis=29∶1)在1×TBE电泳液中进行检测。电泳条件为温度4℃、电压150 V,12~14 h。银染显色后,进行基因分型,不同基因型纯合子进行测序比对。
1.5 肉质、脂肪酸和氨基酸测定
29只杜泊×小尾寒羊杂交羊进行屠宰,并测定肉质性状,包括pH、失水率、熟肉率和剪切力,测定方法参照文献[8],并且在屠宰过程中取12~13肋骨处背最长肌,用自封袋封好后送往吉林省农业科学院农业资源与环境研究所测量脂肪酸和氨基酸含量。
1.6 数据统计分析
根据基因分型结果计算基因型频率和基因频率,并进行卡方检验。利用SPSS 13.0统计软件对各生产性状进行方差分析,不同标记基因型之间生产性状指标(平均值±标准差)进行差异显著性检验和多重比较。
2 结果与分析
2.1 PCR-SSCP检测结果
PCR扩增产物大小与预期一致,无非特异性条带,可以用于SSCP分析。PCR-SSCP检测结果发现有3种基因型AA、BB和AB(见图1)。
2.2 序列分析结果
利用DNAStar 7.0软件与绵羊H-FABP基因序列(GenBank登录号:AY157617)比对发现,AA基因型与BB基因型均在在778位置发生碱基C缺失。AA基因型在1013处发生A→C突变,BB基因型在第939处、980处和1018处分别发生A→G、G→A和A→C单碱基突变。

图1 H-FABP基因PCR产物1%琼脂糖凝胶电泳和SSCP检测结果Fig.1 1%agarose gel electrophoresis and SSCP test results of PCR products in H-FABP gene
2.3 基因型频率和基因频率
经SSCP分型后计算基因型频率和等位基因频率,结果见表1。由表1可知,B等位基因为优势等位基因,基因型BB为优势基因型,并且经卡方检验发现,杜泊×小尾寒羊杂交羊处于Hardy-Weinberg平衡状态(P>0.05)。
2.4 不同基因型对肉质、脂肪酸、氨基酸性状影响
表2结果表明,不同肉质性状均值差异显著性检验发现,不同基因型剪切力存在显著差异,即基因型BB显著低于AB型(P<0.05),说明基因型BB对剪切力有一定负效应。
表3为H-FABP基因不同基因型各脂肪酸性状均值差异显著性检验结果,不同基因型棕榈酸、硬脂酸、油酸和亚油酸存在显著差异:基因型BB在棕榈酸含量上显著高于AA和AB型(P<0.05),说明基因型BB对棕榈酸含量有显著正效应(P<0.05);基因型BB在硬脂酸、亚油酸含量上极显著低于AA、AB型(P<0.01),AA和AB型之间差异不显著,说明BB型对硬脂酸、亚油酸含量有极显著负效应。
H-FABP基因各基因型之间不同氨基酸含量均值差异显著性比较结果发现(表4),不同基因型谷氨酸、丝氨酸、苏氨酸、丙氨酸和脯氨酸含量存在显著性差异。基因型AA、AB在谷氨酸、丝氨酸、苏氨酸、丙氨酸含量以及氨基酸总和上显著高于BB型(P<0.05),说明BB型对谷氨酸、丝氨酸、苏氨酸、丙氨酸含量、氨基酸总和有显著负效应;基因型BB在脯氨酸含量上极显著低于AA、AB型(P<0.01),说明BB型对脯氨酸含量有极显著负效应。

表1 杜泊×小尾寒羊杂交羊H-FABP基因exon2基因型频率及等位基因频率分布Table 1 Distribution of genotype and allele frequencies at the exon 2 of H-FABP gene in Dorper×Small-Tailed Han sheep

表2 H-FABP基因各基因型肉质性状均值差异显著性检验Table 2 Significance test of meat trait means for different genotypes of H-FABP gene

表3 H-FABP基因各基因型脂肪酸性状均值差异显著性检验Table 3 Significance test of fatty acid means for different genotypes of H-FABP gene
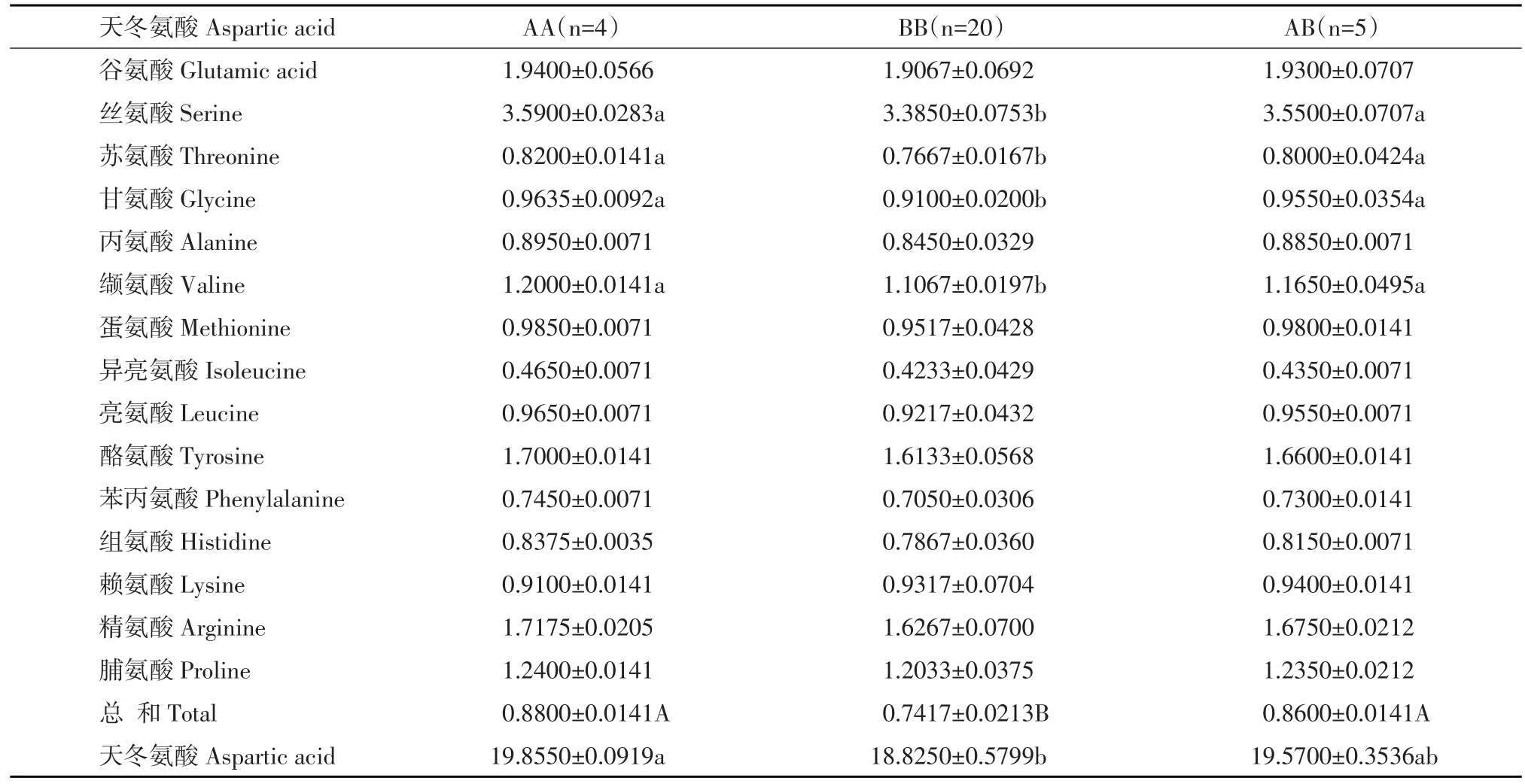
表4 H-FABP基因各基因型氨基酸均值差异显著性检验Table 4 Significance test of amino acids means for different genotypes of H-FABP gene
3 讨论与结论
H-FABP主要在脂肪代谢中起作用,参与长链脂肪酸摄取或利用,把脂肪酸从细胞膜运输到线粒体β氧化位点[9]。此外,通过与脂酰辅酶A和酰基-L-肉碱绑定长链脂肪酸胞内运输,H-FABP基因也参与胞质疏水配体结合蛋白和脂质代谢[10]。目前H-FABP基因与肉质性状关联分析多见于猪和鸡上。李武峰等在不同品种牛群体中发现H-FABP基因多态性对嫩度有一定影响[11-12];余刚等研究表明H-FABP基因可以作为陕北白绒山羊生长及胴体性状候选基因以及绒山羊肉用性能分子标记[13];关于羊H-FABP基因多态性与肉质性状关系报道较少,本研究进行H-FABP基因多态性与肉质性状关联分析,发现BB基因型对剪切力有显著负效应,剪切力越大,嫩度越差,所以说BB基因型对嫩度有显著正效应,这与李武峰和周国利结果相似。应用此标记进行标记辅助育种将有望提高羊肉嫩度,但还需进一步验证。
研究表明不饱和脂肪酸(如油酸、亚油酸和亚麻酸等)含量与肌肉风味和营养价值显著相关[14]。本研究中在肌肉组织中不饱和脂肪酸含量高于饱和脂肪酸含量,这可能是羊肉特有风味物质基础。饱和脂肪酸含量主要取决于棕榈酸和硬脂酸含量,棕榈酸含量与多汁性呈负相关;硬脂酸可引起羊肉膻味,并且随着含量升高而增加[15,16]。本试验对不同基因型脂肪酸含量进行研究,不同基因型棕榈酸、硬脂酸和亚油酸含量存在显著性差异,其结果对进一步研究H-FABP基因与脂肪酸含量关系奠定基础。
H-FABP基因各基因型之间不同氨基酸含量显著性比较结果表明,不同基因型间谷氨酸、丝氨酸、苏氨酸、丙氨酸、脯氨酸含量以及氨基酸总和存在显著性差异,说明H-FABP基因对氨基酸含量有一定影响。氨基酸中丝氨酸、谷氨酸、甘氨酸、亮氨酸、异亮氨酸、丙氨酸和脯氨酸是形成肉香味所必需前体氨基酸,尤其谷氨酸是肉中最主要鲜味物质[17],本试验中不同基因型谷氨酸含量最高,而谷氨酸可增加羊肉鲜味和香味[18],由此说明HFABP基因对羊肉鲜味和香味有影响,
通过对与生产性状有显著相关的位点进行标记辅助选择,将导致经济性状性能产生遗传反应。但由于试验方法及判型标准、统计方法不同,结论存在差异,需要尽快寻求一种更为灵敏的通用检测方法及分析手段进一步验证,以获得可靠数据用于生产。
[1] 王兰萍,耿荣庆,冀德君,等.江苏地方山羊品种H-FABP基因SNPs检测及群体遗传学分析[J].中国畜牧兽医,2012,39(3):159-161.
[2] 乔海云,赵倩君,姚娜,等.绵羊H-FABP基因单核苷酸多态性研究[J].遗传,2009,31(7):725-731.
[3] Gerbens F,Van A E,Harders F L,et al.Effect of genetic variants of the heart fatty acid-binding protein gene on intramuscular fat and performance traits in pigs[J].Journal of Animal Science,1999,77(4):846-852.
[4] Gerbens F,Verburg F J,Van H M,et al.Associations of heart and adipocyte fatty acid-binding protein gene expression with intra⁃muscular fat content in pigs[J].Journal of Animal Science,2001,79(2):347-354.
[5] 李文娟,李宏宾,文杰,等.鸡H-FABP和A-FABP基因表达与肌内脂肪含量相关研究[J].畜牧兽医学报,2006,37(5):417-423.
[6] 郝莉,李齐发,乔永,等.湖羊肌肉组织H-FABP和PPARγ基因表达水平与肌内脂肪含量相关研究[J].中国农业科学,2008,41(11):3776-3783.
[7] 姜延志,刘晓研,李芳琼,等.猪H-FABP基因多态性及其与肌内脂肪含量相关研究[J].畜牧兽医学报,2010,41(7):792-796.
[8] 赵有璋.羊生产学[M].北京:中国农业出版社,2005.
[9] Gerbens F,Verburg F J,Van Moerkerk H T B,et al.Associations of heart and adipocyte fatty acid-binding protein gene expression with intramuscular fat content in pigs[J].J Anim Sci,2001,79:347-354.
[10] Glatz J F,Schaap F G,Binas B,et al.Cytoplasmic fatty acidbinding protein facilitates fatty acid utilization by skeletal muscle[J].Acta Physiol Scand,2003,178:367-371.
[11] 李武峰,许尚忠,曹红鹤,等.3个杂交牛种H-FABP基因第二内含子遗传变异与肉品质性状相关分析[J].畜牧兽医学报,2004,35(3):252-255.
[12] 周国利,朱奇,郭善利,等.鲁西黄牛H-FABP基因多态性及其与肉质性状关系分析[J].西北农业学报,2005,14(3):5-7.
[13] 余刚,罗军,韩雪峰,等.陕北白绒山羊H-FABP基因SNPs及其与生长、胴体性状相关研究[J].畜牧兽医学报,2007,38(11):1154-1159.
[14] Lunt D K,Smith S B.Wagyu beef holds profit potential for US[J].Feedlots,1991,(8):18.
[15] 马文习.脂蛋白与载脂蛋白[J].生物学通报,1998,33(6):17-18.
[16] 钱文熙.滩羊肉品质研究[D].银川:宁夏大学,2005.
[17] 宋社果,安小鹏,赵海波,等.藏香猪屠宰特性及肉品质分析[J].西北农业学报,2011,20(12):26-32.
[18] 热孜瓦古丽·米吉提,买买提伊明·巴拉提,依巴代提·米吉提,等.绵羊羊肉氨基酸成分比较研究[J].新疆农业科学,2012,49(8):1552-1556.
猜你喜欢
今日农业(2022年2期)2022-11-16
世界科学技术-中医药现代化(2022年3期)2022-08-22
美食(2022年4期)2022-04-16
昆明医科大学学报(2021年3期)2021-07-22
美食(2020年5期)2020-06-01
中央民族大学学报(自然科学版)(2015年1期)2015-06-11
现代检验医学杂志(2015年6期)2015-02-06
作物研究(2014年6期)2014-03-01
中国神经精神疾病杂志(2013年4期)2013-03-11
中国糖料(2013年1期)2013-01-22
