
桃蛀果蛾2种寄主的挥发物鉴别
2013-07-19 07:57孔维娜贺润平
山西农业科学 2013年3期
李 捷 ,王 慧 ,孔维娜 ,赵 飞 ,贺润平
(1.山西省农业科学院果树研究所,山西太谷030815;2.山西省农业科学院高寒区作物研究所,山西大同037008;3.山西省农业科学院植物保护研究所,山西太原030032)
桃蛀果蛾(Carposina niponensis),又名桃小食心虫,广泛分布在我国北方果区,其食性杂,可为害苹果、枣、梨、杏、海棠等10多种果树的果实。近年来果树套袋在一定程度上控制了该虫在苹果和梨上的危害,但由于枣果和杏等不能套袋,则有为害加重的趋势。
植物在自然界中释放出大量的信息化合物,这些化合物在植食性昆虫寻找、识别和定位寄主植物的过程中扮演着重要的角色,而且在成虫产卵和取食选择中起至关重要的作用[1-2]。另外,在植物防御方面,它可通过释放挥发物吸引天敌、驱避害虫和阻止害虫取食,进行有效的直接或间接防御,以保证自身的安全[3-4]。可见,植物挥发物作为信息化合物在植物与昆虫的协同进化过程中对昆虫行为产生了十分重要的影响,并在害虫的及时、有效测报及其综合治理中发挥着十分重要的作用[5]。近10多年来,植物挥发物的研究已取得很大进展,特别是挥发物顶空吸附和GCMS分析技术以及近年来广泛应用的固相微萃取(SPME)技术,为该领域的研究提供了重要的技术支撑[6]。
本试验以桃蛀果蛾寄主选择期(即寄主幼果期)的枣、杏为材料,采用固相微萃取和GC-MS分析鉴别桃蛀果蛾不同寄主的挥发性组分及其含量,旨在为进一步研究桃蛀果蛾与寄主间化学通讯机理提供依据。
1 材料和方法
1.1 供试材料
本试验所用寄主植物为枣和杏,采集地点为山西省农业科学院果树研究所。分别在2010年4月下旬采集杏的幼果、6月下旬采集枣吊。将所采集的样品用蒸馏水清洗、晾干,待用。
1.2 挥发物的收集与分析
取待测材料100 g,放入500 mL的广口瓶内,用封口膜封口后,在室温下,将固相微萃取装置(DVB/CAR/PDMS)的针头穿透封口膜,插入容器内上方的空白处,然后小心地推出纤维头,同时用固定装置固定固相微萃取装置的手柄,自萃取纤维头推出后开始计时(吸附45 min),待吸附完毕后立即进行GC-MS分析,每个样品重复 3 次[7]。
挥发物的鉴定利用美国Thermo-Finnigan公司的TraceDSQ气相色谱-质谱仪(GC-MS)分析,色谱柱利用SE-54毛细管柱(30 m×0.25 mm ID,膜厚 0.25 μm),载气氦气;流速 1 mL/min,进样1 μL,分流比50∶1。起始温度50℃,停留3 min,以10℃/min升到220℃。质谱条件:EI离子源,电离能70 eV。
各成分通过与谱库(NIST2002版)标准化合物的质谱图核对并分析后,进行定性,根据峰面积归一化法进行定量[8-9]。
2 结果与分析
桃蛀果蛾2种寄主挥发物成分的比较如表1所示。
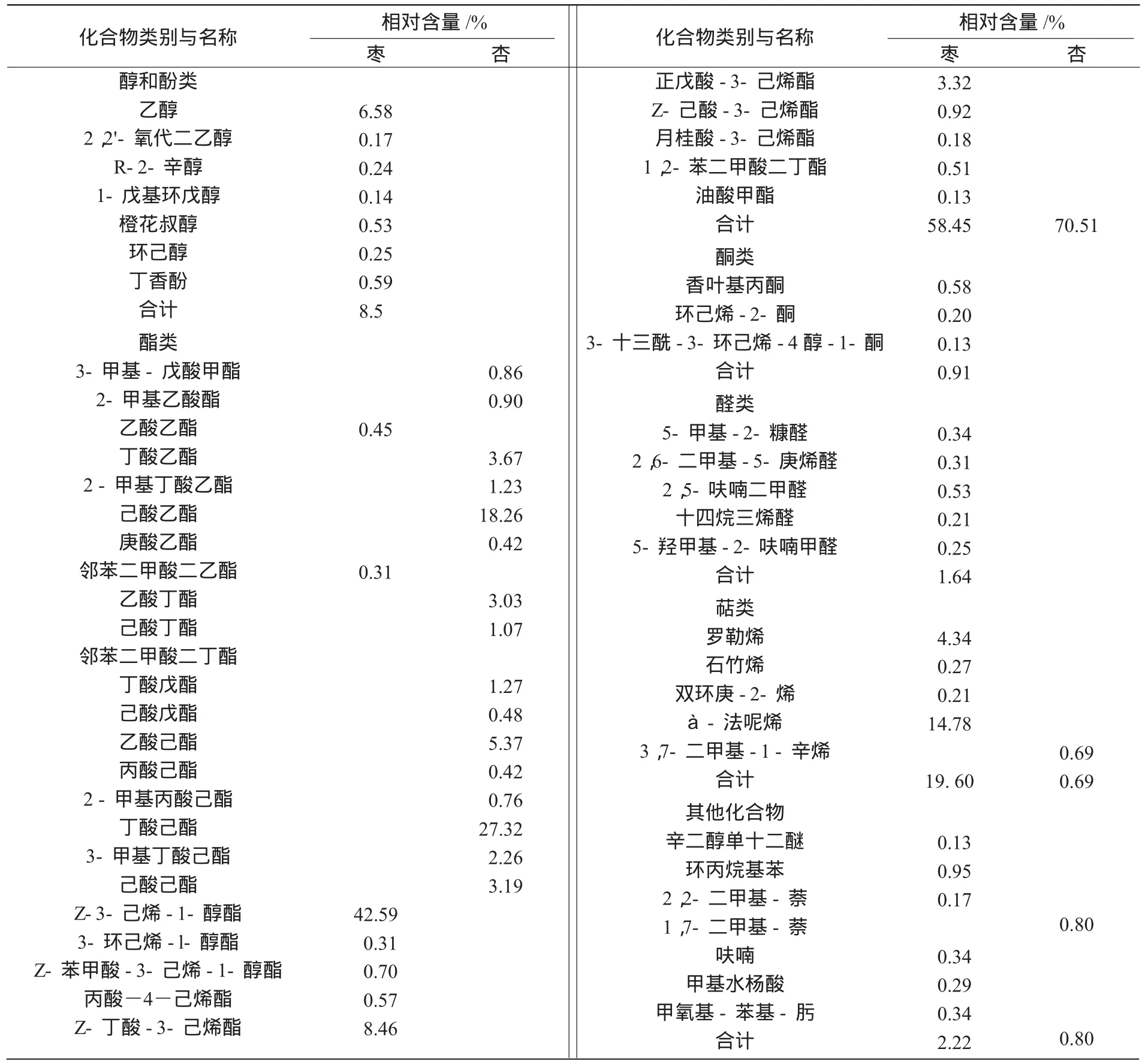
表1 桃蛀果蛾2种寄主挥发物成分的比较
从表1可以看出,桃蛀果蛾2种寄主枣、杏在幼果期挥发物(除烃类化合物)有显著差别,分别为37种和18种,主要包括醇(酚)类、醛类、酯类、萜类和其他类等。枣和杏在幼果期挥发物组成存在很大差异,枣幼果期挥发物成分以酯类和萜类化合物为主,分别占到挥发物总成分的58.45%和19.60%,Z-3-己烯-1-醇酯的相对含量最高,占总量的42.59%,à-法呢烯的相对含量次之,为14.78%;杏幼果期挥发物成分主要是酯类,约占挥发物总成分的70.51%,丁酸己酯的相对含量最高,为27.32%,己酸乙酯的相对含量次之,为18.26%。
3 结论与讨论
本试验采用固相微萃取法采集桃蛀果蛾2种寄主的挥发物,该方法集采样、萃取、浓缩、进样于一体,便于携带,真正实现了样品的现场采集和富集。
SPEM结果表明,枣和杏幼果期挥发物组成有很大差异,分别为37种和18种,枣幼果期以Z-3-己烯-1-醇酯和à-法呢烯、杏幼果期以丁酸己酯和己酸乙酯含量较高。
不同寄主挥发物其功能不同,目前的研究表明,Z-3-己烯-1-醇酯与性信息素以1∶1混合所引诱的菜蛾数量为仅用性激素的6~7倍[10],à-法呢烯是苹果蠹蛾(Laspeyresia pomonella)寄主趋向的主要挥发物[11],GC-EAD测试表明,苹果蠹蛾对乙酸己酯、丁酸己酯和à-法呢烯有明显反应[12],罗勒烯是叶螨天敌的引诱物质[13],而罗勒烯和Z-3-己烯-1-醇酯是枣镰翅小卷蛾(Ancylis sativa)的主要寄主趋向物质[14];此外,甲基水杨酸不但是引诱昆虫天敌的重要信息物质[15],而且是植物间通信的重要信号物质[16],其他化合物的功能,目前仍不清楚。2种寄主共鉴定出55种化合物,哪些是桃蛀果蛾寄主趋向的主要物质,还有待于进行进一步深入研究。
[1]Bernays E A,Chapman R F.Host-plant selection by phytophagousinsects[M].London:Chapman&Hall,1994:206-223.
[2]Tholl D B,Hansel W,Loreto A F,et al.Practical approaches to plant volatile analysis[J].The Plant Journal,2006,45(4):540-560.
[3]NosovAM.Functions ofplant secondarymetabolites in-vivoand in-vitro[J].Journal ofPlant Physiology,1994,41(6):767-771.
[4]Heath M C.In this issue:secondary metabolites and plant defense[J].Physiological and Molecular Plant Pathology,2002,60(6):273-274.
[5]莫圣书,赵冬香,陈青.植物挥发物与昆虫行为关系研究进展[J].热带农业科学,2006,26(6):84-89.
[6]阎凤鸣.化学生态学[M].北京:科学出版社,2003.
[7]王明林,乔鲁芹,张莉,等.固相微萃取-气相色谱/质谱测定植物叶片中的挥发性物质[J].色谱,2006,24(6):343-346.
[8]李新岗,刘惠霞,刘拉平,等.影响松果梢斑螟寄主选择的植物挥发物成分研究[J].林业科学,2006,42(6):71-78.
[9]李新岗,刘惠霞,刘拉平,等.油松球果对外源茉莉酸甲酯和虫害诱导的生化反应[J].林业科学,2007,43(3):66-72.
[10]Reddy G V P,Guerrero A.Behavioral responses of the diamondback moth,Plutella xylostella,to green leaf volatiles of Brassica Oleracea subsp.Capitata[J].Journal of Agricultural and Food Chemistry,2000,48(12):6025-6029.
[11]Witzgall P,Stelinsk L,Gut I L,et al.Codlingmoth management and chemical ecology[J].Annual Review of Entomology,2008,53:503-522.
[12]CasadoD,GemenoC,Avilla J,et al.Day-night and phonological variation of apple tree volatiles and electroantennogram responses in Cydia pomonella(Lepodoptera:Tortricidae)[J].Environmental Entomology,2006,35(2):258-267.
[13]Dicke M,Abelis M W,Takabayashi J,et al.Plant strategies of manipulating predator-prey interactions through allelochemicals:Prospects for application in pest control[J].Chemistry and Ecology,1990,16:3091-3117.
[14]杨立军.基于寄主挥发物的枣镰翅小卷蛾寄主选择研究[D].杨凌:西北农林科技大学,2011.
[15]Degenhardt J,Gershenzon J,Baldwin I T,et al.Attracting friends to feast on foes:engineering terpene emission to make crop plants more attractive to herbivore enemies[J].Plant Biotechnology,2003,14:169-176.
[16]ShulaevV,Silverman P,Raskin I.Airborne signalingbymethyl salicylate in plant pathogen resistance[J].Nature,1997,385:718-721.
猜你喜欢
中国饲料(2021年17期)2021-11-02
今日农业(2021年10期)2021-07-28
当代水产(2021年2期)2021-03-29
传染病信息(2021年6期)2021-02-12
河北果树(2020年4期)2020-11-26
化工管理(2020年26期)2020-10-09
烟草科技(2015年8期)2015-12-20
山西大同大学学报(自然科学版)(2015年1期)2015-01-22
食品工业科技(2014年7期)2014-03-11
中国造纸(2014年1期)2014-03-01
