
106份狗牙根材料的SRAP分析
2013-05-13 08:11张延辉李江华帕提古丽阿不来提
草原与草坪 2013年2期
张延辉,李江华,孙 强,帕提古丽,阿不来提
(1.新疆农业大学 草业与环境科学学院/新疆草地资源与生态重点实验室,新疆 乌鲁木齐 830052;2.新疆维吾尔自治区乌鲁木齐市公安局,新疆 乌鲁木齐 830052;3.新疆维吾尔自治区草原总站,新疆 乌鲁木齐 830052)
狗牙根(Cynodon dactylon)是暖地型草坪草中最重要的草种之一,且在世界各地广泛种植,狗牙根属共有9个种10个变种[1]。我国华南、华中、西南、西北、华北南部均有狗牙根分布[2]。不同地域的狗牙根即使在同一种内仍然存在着及其丰富的遗传变异。评价一种种质资源的遗传多样性,对深入了解该资源的种群结构、变异程度、居群丰富度,开展核心种质库的构建及优良性状的选育工作具有重要的意义。新疆南北疆各地均有狗牙根分布,这些野生狗牙根的不同居群在形态、地域环境上差别很大,表现为不同的生态型,阿不来提将分布于南北疆平原绿洲不同生态条件下的狗牙根分为伊犁型(北疆型)和喀什型(南疆型)两大类[3]。目前,新疆狗牙根的研究工作涉及各方面,包括形态、抗性、生理[4,5]等,但分子水平方面的研究刚刚起步,笔者仅就部分野生材料进行了SRAP标记研究,而较全面的评价新疆地区的狗牙根材料,亟需在建立较完整种质资源圃的情况下利用分子标记技术探究其遗传变异,从DNA水平评价其遗传多样性。
相关序列扩增多态性(SRAP)标记方法具有简便、中等产量、高共显性、易于分离条带及测序,不需预知物种的序列信息等优点[6]。相比于SSR标记,其引物开发的成本大大降低,而且该技术主要针对ORF开放阅读框设计引物,对内含子、外显子及启动子区域进行扩增,比较稳定。目前,在草坪草的选育上,运用该技术开展多样性研究还较少。Budak等[7]利用34对SRAP引物组合分析了53份野牛草种质的遗传多样性及其与表型的关系。易杨杰等[8]对四川、重庆、贵州、西藏4省区的野生狗牙根材料进行了遗传多样性分析。
利用SRAP标记技术对106份狗牙根材料进行遗传多样性研究,旨在揭示不同野生狗牙根材料间的亲缘关系,遗传背景、变异程度及遗传多样性水平,为选育优良的狗牙根新品种提供理论依据。
1 材料和方法
1.1 供试材料
供试的狗牙根材料共106份,部分采自新疆不同地区,部分引自国内外其他地区,狗牙根材料编号及来源见表1。2008年5月将供试的狗牙根材料种植于新疆农业大学试验场原始资源圃,小区面积1.0m×2.5 m,小区间距70cm,期间进行灌溉、杂草防除等田间管理工作。
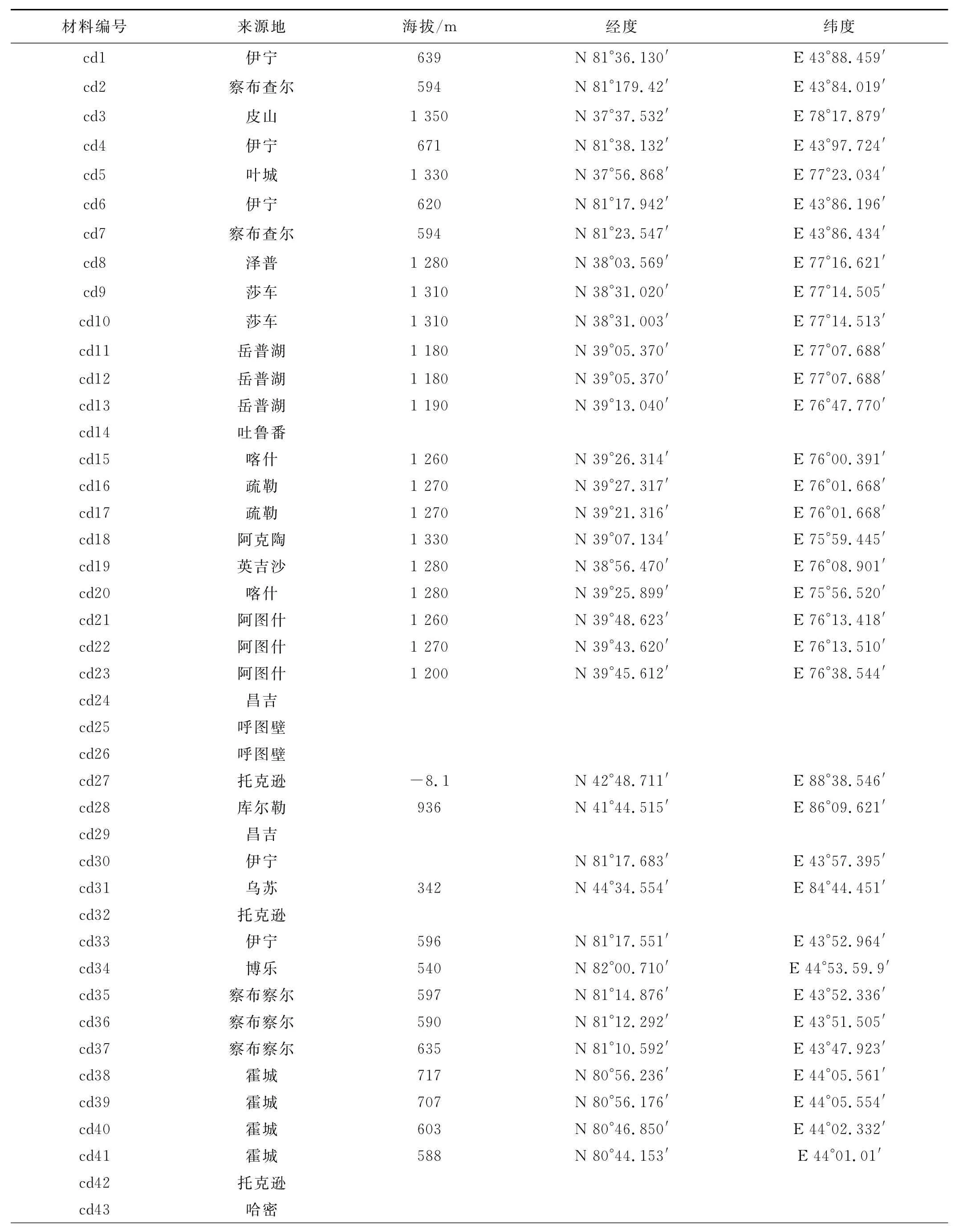
表1 106份野生狗牙根的采集地点Table 1 The collection areas of 106bermudagrass accessions
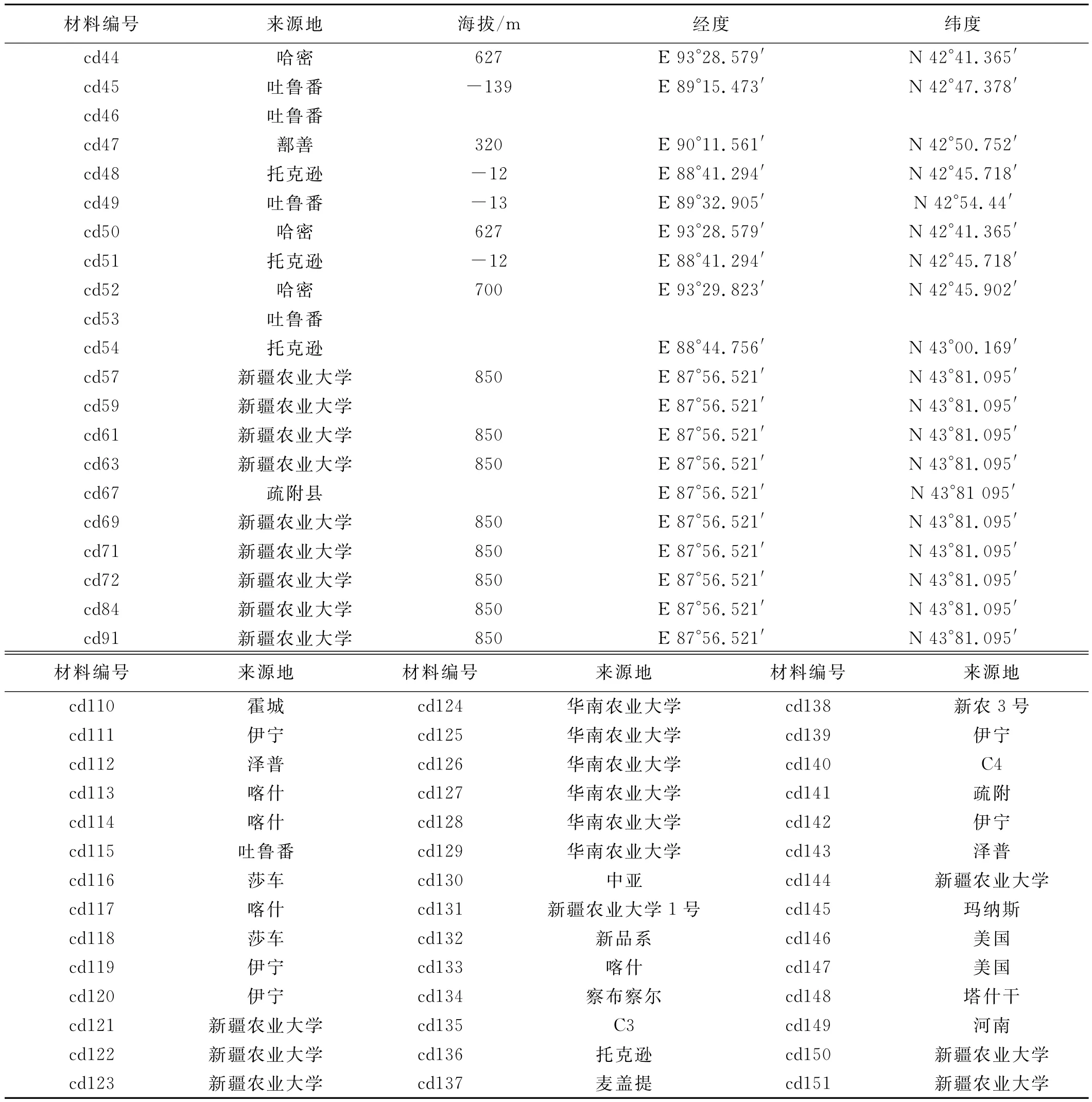
续表1
1.2 试验方法
采用CTAB法[12]提取样品总 DNA,从238对SRAP引物组合筛选出14对多态性较好的组合用于PCR扩增,采用10μL反应体系,反应液含1.4μL 10×Buffer,0.25mmol/L dNTP,1μmol/L引物,0.2U的Taq DNA聚合酶及15ng模板DNA。SRAP-PCR扩增在PTC2200(MJ,USA)PCR仪上完成。
扩增程序:94℃预变性5min;94℃变性1min,35℃复性1min,72℃延伸1min,循环5次;94℃变性1min,50℃复性1min,72℃延伸1min,循环35次;72℃延伸10min,4℃保存。扩增产物加3μL上样缓冲液,用6%非变性PAGE电泳检测,上样量3 μL,3μL 100bpLadderMarker。电压200V,电泳约60min,采用银染,数码相机照相记录结果。同时设置不含DNA样本的阴性对照(用ddH2O代替DNA),以确定每次PCR实验时材料的DNA是否存在自扩增或污染。
1.3 数据统计分析
根据扩增产物的条带有或无分别赋值,有带的记为1,无带的记为0,获得二元数据矩阵。利用NTSYS-pc2.1统计分析软件计算不同材料间的遗传相似系数(GS)。根据GS值按非加权平均距离(UPGMA)进行遗传相似性聚类分析,利用POPGENE软件进行各项遗传参数的统计分析。
2 结果与分析
2.1 引物筛选结果及扩增效果的比较
通过对14对正向引物,17对反向引物进行两两组合,共形成238对引物组合。通过对该引物组合进行筛选,共获得14对多态性较高、条带清晰、稳定性和重复性好的引物组合(表2),运用该引物组合分别对106份材料的基因组DNA进行扩增。结果表明,14个引物组合共产生133条带,平均每个引物产生9.5条,其中有123条显示多态性,多态性百分率为92.5%,Nei’s基因多样性指数(H)变异为0.244 2~0.499 8,平均为0.414 5(表3)。其中,引物组合Me21-Em13扩增的总位点最多,有12个,引物组合Me17-Em12和Me22-Em18多态性位点最少,仅有7个。供试的样品具有不同的SRAP带型,扩增片段在100bp~1 500bp,多态性检出率较高。表明SRAP能检测的遗传位点比较丰富,初步表明新疆野生狗牙根在DNA分子水平遗传多样性很丰富。

表2 14对SRAP引物序列及其扩增结果Table 2 Sequences of 14SRAP primers and amplification results
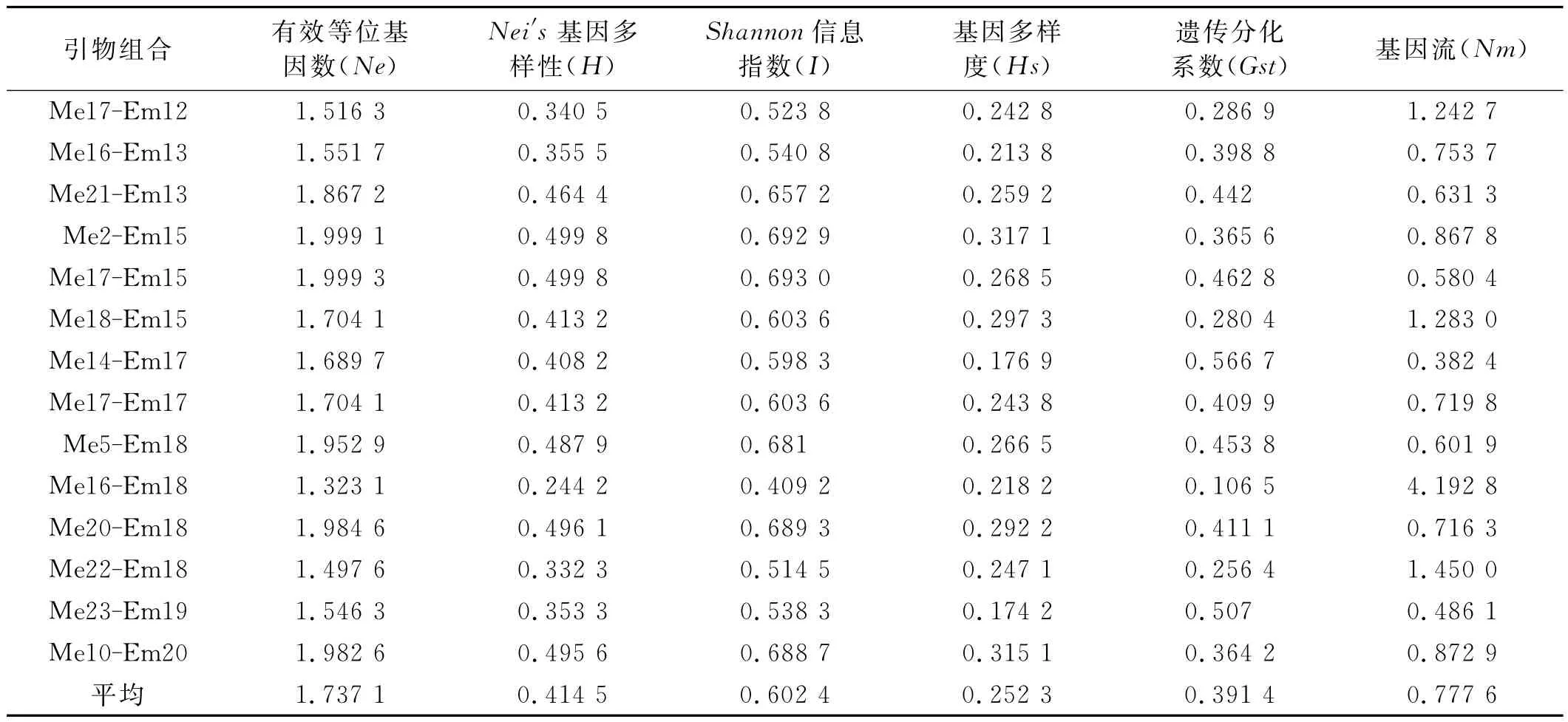
表3 研究所用SRAP引物组合的扩增结果及多态性信息Table 3 Results and the polymorphism information of SRAP primer combinations used in the present study
2.2 遗传多样性分析
对SRAP-PCR扩增结果采用 Nei-Li相似系数(GS)的计算方法,得到相似性矩阵。供试材料之间差异明显,具有较为丰富的遗传多样性(表4),有效等位基因数(Ne)在1.236 3~1.593 8,Nei′s基因多样性指数(H)在0.159 0~0.347 4,Shannon信息指数(I)在0.254 0~0.516 7,表明新疆野生狗牙根材料种内存在广泛的变异,其中以材料cd01多样性最为丰富,Nei′s基因多样性(H)为0.347 4,Shannon信息指数(I)为0.516 7。
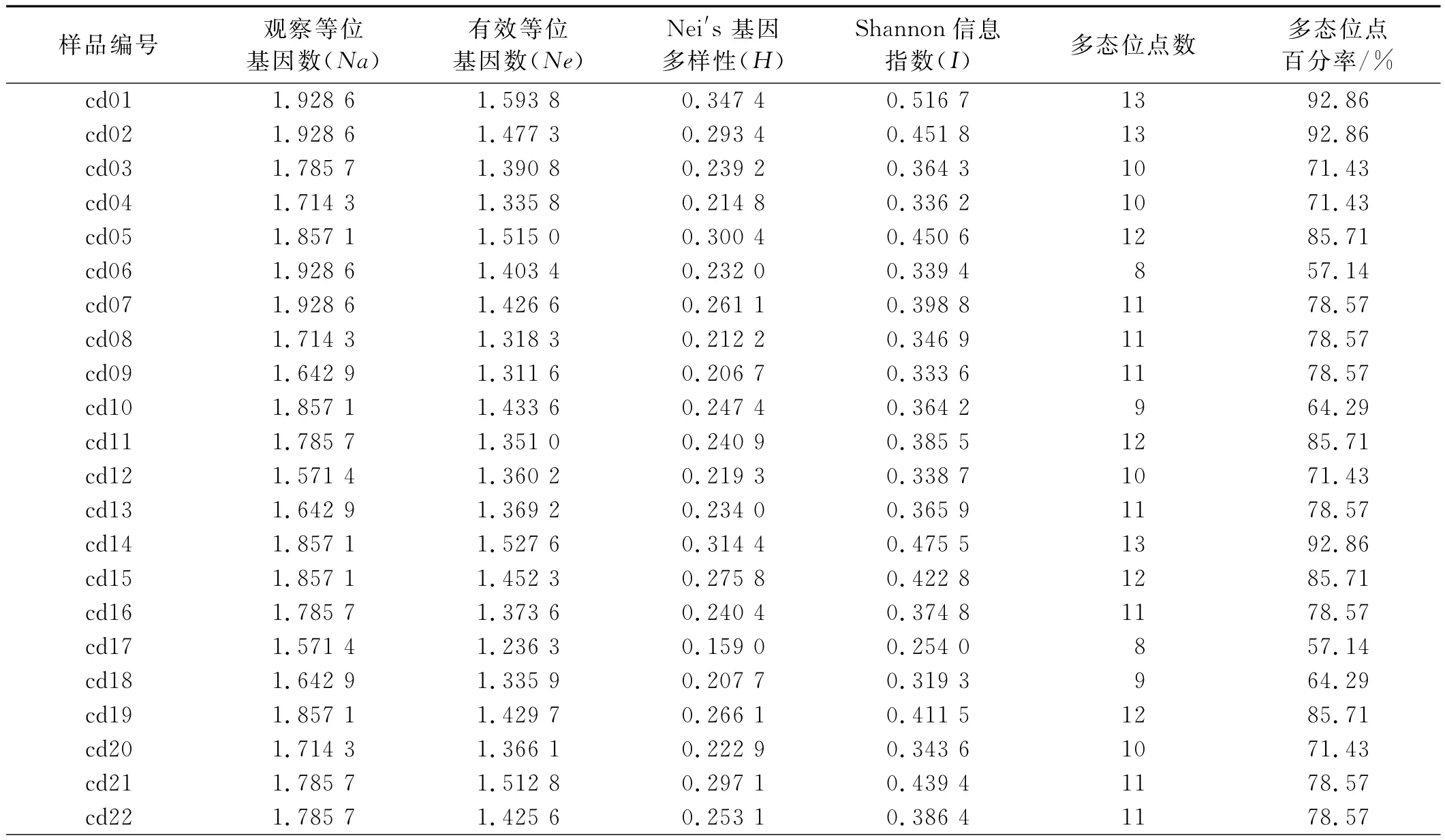
表4 SRAP标记分析狗牙根的遗传多样性Table 4 Genetic diversity of native bermuda grass by SRAP mark
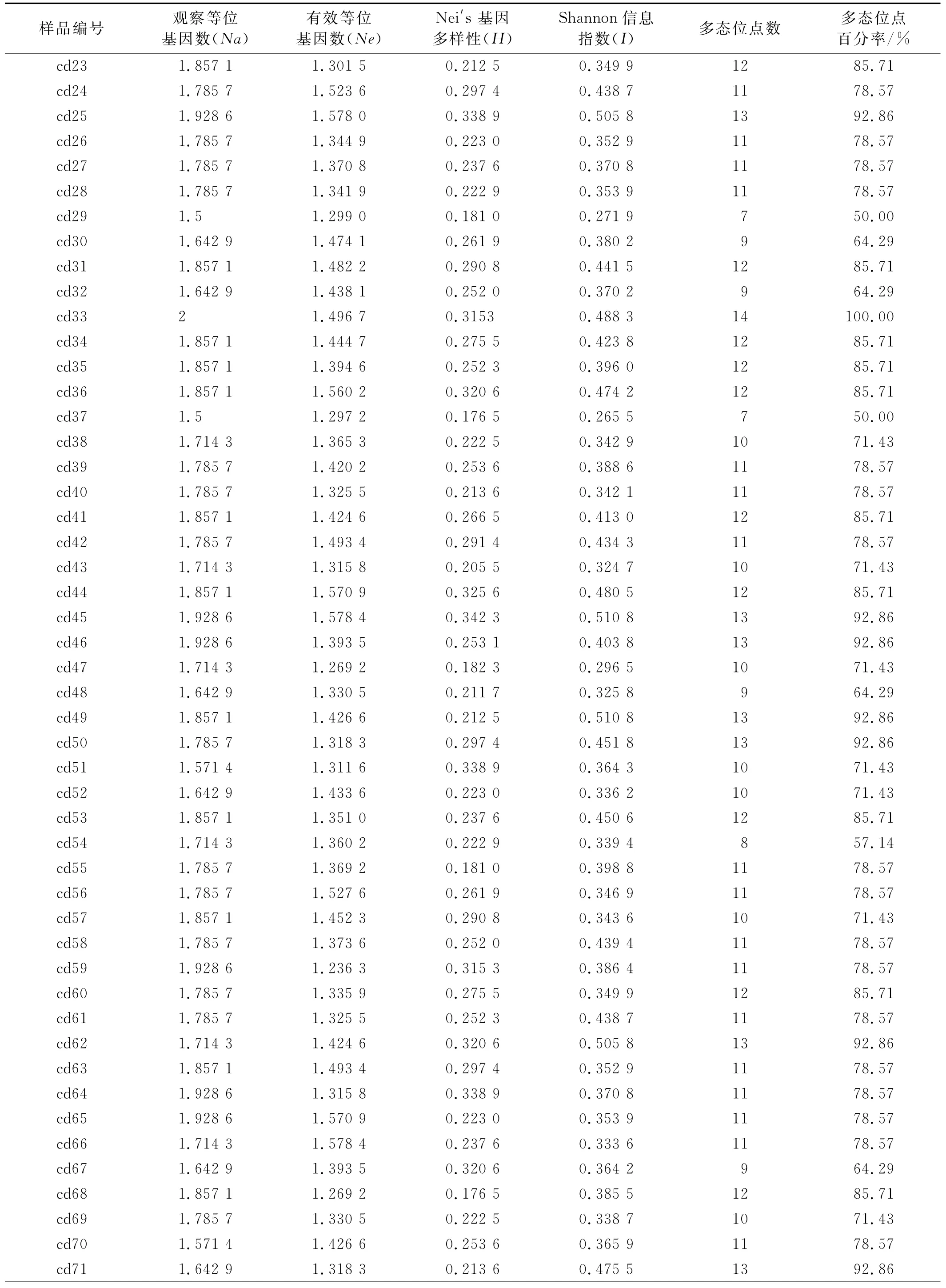
续表4

续表4
2.3 聚类分析
根据14对引物组合的PCR扩增结果,基于遗传相似系数,利用UPGMA法绘制出供试材料SRAP标记遗传多样性分子聚类图(图1)。从聚类图上可以看出,106份供试材料在L=0.51时,可以分成3个组:在第1大组中,共计90份材料,被分成了3个亚类。其中,来自伊犁的一种材料和采自华南的材料单独聚为一类(cd001和cd128),而且距离其他材料较远。聚类结果显示,不同来源地的材料也有聚在一起,如cd16、cd20;也有部分来源地相同的材料聚在不同的亚类中,如cd15、cd114、与cd117,说明来源地相同的材料有可能是同一种材料,其遗传相似度极高,但不同来源的材料也聚在一大组中,说明这些材料有一定程度的变异。在第2组中,共计15份材料,cd37自为一类,而cd32、cd49、cd50聚在一起,遗传基础较为相似,这些材料普遍来自气候较热的地区,比如哈密、吐鲁番,说明具有较一致的生态型的材料容易聚到一起。在第3组中cd129自成一类,同其他材料遗传距离较大,亲缘关系比较远。从聚类结果也可以看出,SRAP标记对材料的区分能力及鉴别的准确性比较好。

图1 106份狗牙根材料UPGMA聚类图Fig.1 Dendrogram of 106genotypes from the UPGMA cluster analysis
3 讨论与结论
3.1 新疆狗牙根的多态性
利用SRAP标记技术分析了106份狗牙根材料的遗传多样性,从238个引物组合中筛选出多态性高的14个引物组合,共扩增出123条多态性条带,定位了523个多态性位点,其中多态性条带比例为92.5%,表明新疆野生狗牙根资源具有较高的遗传多样性。已有的一些研究结果在多态性统计上有一些差异。凌瑶测定的中国西南5省区的44份野生狗牙根及8份非洲狗牙根材料的多态性位点为87.29%[9];易杨杰对野生狗牙根种质研究,多态性条带比例为79.8%[8]。周少云等[10]应用ISSR标记对广东地区30份野生狗牙根进行遗传多样性研究,多态性条带比率达93.7%。这些研究结果的差异主要与不同地域野生狗牙根材料本身具丰富的遗传变异,以及材料样本选取量大小有关。
3.2 狗牙根的遗传相似性
根据此次试验的结果,106份狗牙根的遗传相似性GS值在0.36~0.94,平均为0.65。齐晓芳等[11]利用AFLP标记对采自非洲以及中国四川、重庆、云南、贵州和西藏5省区的共44份野生狗牙根材料进行遗传多样性分析,遗传相似系数(GS)为0.64~0.95,平均GS值为0.76。新疆野生狗牙根部分材料间的相似系数较低,遗传距离很远,表明狗牙根种内具有大量可供遗传变异选择的基因型,这为选育优良品种提供了良好的基础。
3.3 新疆狗牙根遗传变异与地理来源的相关性分析
物种的遗传变异和地理生态环境之间的关系一直是植物种群遗传学普遍关注的问题,目前,多数研究认为物种的基因与种质的地理分布间存在有一定的相关性[12]。刘伟等[13]将西南区42份野生狗牙根材料的遗传距离矩阵和与之对应的地理距离矩阵进行相关分析,结果表明大部分野生狗牙根材料的遗传距离与地理来源之间无严格的一致性关系。笔者所得结论认为材料与地理之间有一定的相关性,聚类结果可以在一定程度上反映与地理距离之间的联系。聚类结果比较发现,在大的类群划分上表现出一定的一致性,但在类群内表现出较大的差异。生境相近的大部分品种,在SRAP分子标记上也能聚在一起,而且在地理位置上越接近的品种,在二者的一致性上表现得越显著,相同或相邻生境较易聚在一起的。如吐鲁番、托克逊、哈密等地的材料。在所有的材料中,以来自伊犁的材料遗传多样性最丰富,分布在两个大组中,这可能与伊犁地区地域特点有关。因此,在做聚类分析时应结合形态、生理生化数据及地理特征进一步分析。
[1] Boune J M.Morocco’s native perennial turf grasses are a treasure trove of diversity and they’re disappearing[J].Diversity,2000,16:53-54.
[2] 刘建秀,贺善安.暖季型草坪草种质资源的研究与改良[J].国外畜牧学-草原与牧草,1996(3):12-30.
[3] 阿不来提,石定燧,杨光,等.新疆野生狗牙根研究初报[J].新疆农业大学学报,1998(2):124-127.
[4] 孙宗玖,阿不来提,齐曼,等.狗牙根新品种抗寒性评价及生理基础研究[J].中国草地,2003(4):25-30.
[5] 孙宗玖,阿不来提,齐曼,等.冷害胁迫下3个狗牙根品种抗寒性比较研究[J].草业科学,2004(1):39-42.
[6] Ferriol M,Pico B,Nuez F.Genetic diversity of a germplasm collection of Cucurbita pepo using SRAP and AFLP markers[J].Theoretical and Applied Genetics,2003,107:271-282.
[7] Budak H,Shearman R C,Parmaksiz I,et al.Molecular characterization of Buffalo grass germplasm using sequence-related amplified polymorphism markers[J].Theor Appl Genet,2004,108:325-361.
[8] 易杨杰,张新全,黄琳凯,等.野生狗牙根种质遗传多样性的SRAP研究[J].遗传,2008,30(1):94-100.
[9] 凌瑶,张新全,齐晓芳,等.西南五省区及非洲野生狗牙根种质基于SRAP标记的遗传多样性分析[J].草业学报,2010,19(2):196-203.
[10] 周少云,黄春琼,刘国道.广东地区野生狗牙根遗传多样性的ISSR分析[J].热带作物学报,2010,31(5):797-802.
[11] 齐晓芳,张新全,凌瑶,等.野生狗牙根种质资源的AFLP遗传多样性分析[J].草业学报,2010,19(3):155-161.
[12] Wilson B L,Kitzmiller J,Rolle W,et al.Isozyme variation and its environmental correlates in Elymusglaucusfrom the California Floristic Province[J].Canadian Journal of Botany,2001,79:139-153.
[13] 刘伟,张新全,李芳,等.西南区野生狗牙根遗传多样性的ISSR标记与地理来源分析[J].草业学报,2007,16(3):55-61.
猜你喜欢
今日农业(2022年13期)2022-09-15
世界科学技术-中医药现代化(2022年3期)2022-08-22
中国麻业科学(2018年6期)2018-04-09
西南农业学报(2016年5期)2016-05-17
西南农业学报(2016年6期)2016-04-16
广西林业科学(2016年3期)2016-03-16
中国卫生标准管理(2015年16期)2016-01-20
中央民族大学学报(自然科学版)(2015年1期)2015-06-11
中国当代医药(2015年20期)2015-03-01
西南军医(2015年1期)2015-01-22