
金线莲菌根真菌的分离和鉴定
2013-04-17 02:27朱丹陈卫国杜婵娟付岗黄兴振付书婕赵有林蒋伟哲
环球中医药 2013年4期
朱丹 陈卫国 杜婵娟 付岗 黄兴振 付书婕 赵有林 蒋伟哲
金线莲[Anoectochilusroxburghii(Wall.) Lindl]为兰科的药用植物,生长在土壤腐殖质较多、光线较弱、相对湿度大的阔叶林中[1],具有保肝、防癌、抗氧化、抗炎等多种药理作用[2]。近年来,因其生长环境遭到严重破坏,野生资源被过度采挖,加之具有繁殖率低、生长缓慢等特点,使得金线莲野生资源濒临灭绝[3]。在野生状态下,所有兰科植物都与真菌共生[4-5],野生金线莲亦与内生真菌形成菌根共生关系,筛选促进金线莲生长发育的益生菌,研究它们的相互作用具有重大的理论意义。本课题组在广西桂林等地采集野生金线莲,并从该植物根中分离出多株菌根真菌,研究发现,这些真菌可以增加植物芽和根的数量,促进金线莲试管苗的生长。为了进一步研究菌根真菌对金线莲生长发育的影响,课题组对金线莲菌根真菌进行分离,并采用分子技术结合形态学特征对分离物进行鉴定,为下一步研究奠定基础。
1 材料与方法
1.1 菌根真菌的分离和形态学观察
采集野生金线莲植株,取其根部用无菌水冲洗干净,切成0.5~1 cm小段,75%乙醇表面消毒3~5秒,无菌水冲洗净消毒液后,再用0.1%HgCl2表面消毒6~8分钟,无菌水冲洗后将根段接种于PDA平皿内,25℃恒温培养,待从根内长出真菌菌丝后,挑取尖端菌丝纯化培养。将纯化的6株真菌菌株(编号分别为2、5、6、A12、A46、A55)在PDA培养基(马铃薯200 g、葡萄糖20 g、琼脂17 g、水1000 ml)平板上于28℃恒温下黑暗培养,每天观察菌落的形态特征和颜色变化,并于显微镜下进行形态学观察、鉴定。
1.2 分子生物学特征测定
1.2.1 基因组DNA的提取 收集PDA平板上的菌丝体,采用SDS-CTAB法分别提取各菌株的基因组DNA。具体操作如下:取0.1 g新鲜菌丝体,在液氮中研磨成粉末,迅速转入1.5 ml的离心管中,加入750 μl提取缓冲液,置65℃水浴30分钟;室温下12 000 r/min离心10分钟,取上清液,加入1/10的3 mol/L KAc,冰浴20分钟;室温下12 000 r/min离心10分钟,取上清液,加入等体积的异丙醇,冰浴20分钟;12 000 r/min离心10~20分钟;弃上清,加入400 μl ddH2O及5 μl RNAase,37℃水浴1小时;加入等体积的酚/氯仿(1:1)颠倒混匀40次;室温下12 000 r/min离心10分钟,取上清液,再加入 1/10 KAc,冰浴20分钟(重复1次);用2倍体积无水乙醇沉淀20分钟,12 000 r/min离心10分钟;用-20℃ 预冷的75%乙醇洗涤沉淀3次,室温干燥5~10分钟后加入TE溶液溶解沉淀。
1.2.2 PCR扩增及纯化测序 以各菌株基因组DNA为模板,扩增ITS区的保守区段,选用真菌通用引物ITS1(5′-TCCGTAGGTGAACCTGCGG-3′)和ITS4(5′-TCCTCCG CTTATTGATATGC-3′)进行PCR扩增。PCR扩增体系为20 μl,其中含10×PCR buffer 2 μl,dNTP0.5 μl,ITS1 0.6 μl,ITS4 0.6 μl,无菌超纯水15.1 μl,模板DNA 1 μl,Taq酶0.2 μl。反应程序为:94℃预变性3分钟;94℃变性55秒,56℃退火50秒,72℃延伸l分钟,32个循环;72℃延伸7分钟。扩增产物通过琼脂糖凝胶电泳,以DNA Marker DL2000作为分子量标准,稳压120 V下电泳30分钟。电泳结果用紫外检测仪观察,置于图像处理仪中处理并保存。目的片段用UNIQ-10柱式胶回收试剂盒回收纯化,纯化样品进行基因序列测定。
1.2.3 数据分析 序列经BioEdit软件分析和手工校正后,用NCBI的BLAST程序将测得序列与GenBank中已有菌株的ITS序列进行同源性比对,采用MAGA (3.1)软件进行系统发育树的构建。
2 结果
2.1 形态学特征观察结果
各菌株在PDA培养基上生长,菌落初期灰白色,渐变为淡褐色,平整,生长速度快,气生菌丝繁茂,28℃下培养3天后,直径可达80 mm,7天即可产生菌核(见图1)。菌丝初期灰白色,渐变为淡褐色,直径12~14 μm;菌丝分支处有缢缩,近分支处形成隔膜。菌核由菌丝纠集生成,褐色,内外颜色一致,表面粗糙,大小均一,可聚集联结;无性态不产生孢子(见图2)。
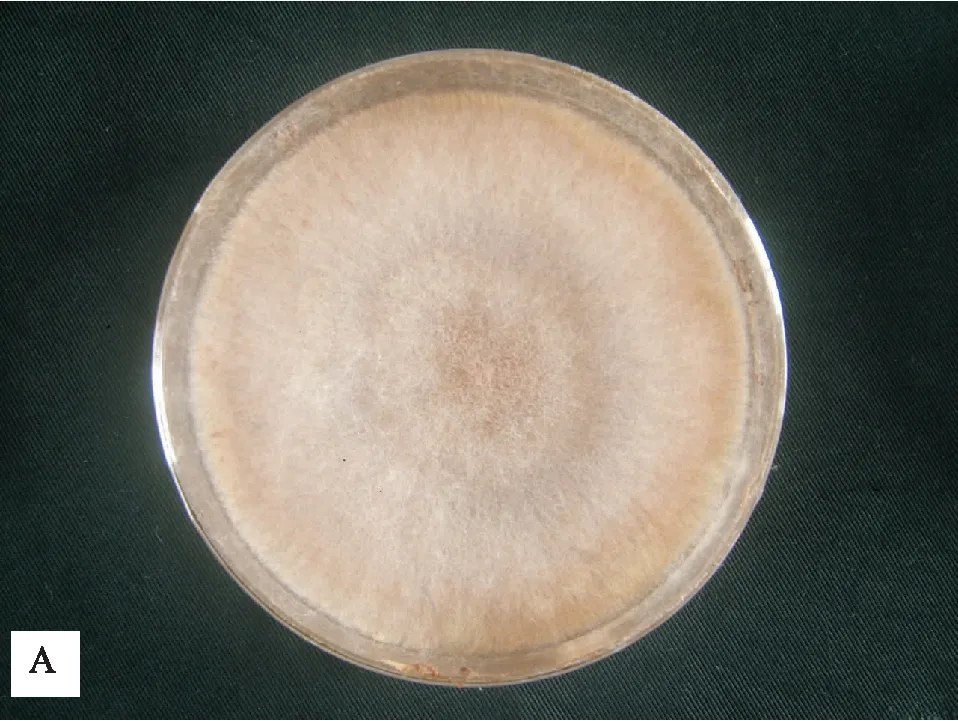
A 金钱莲菌根真菌早期菌落
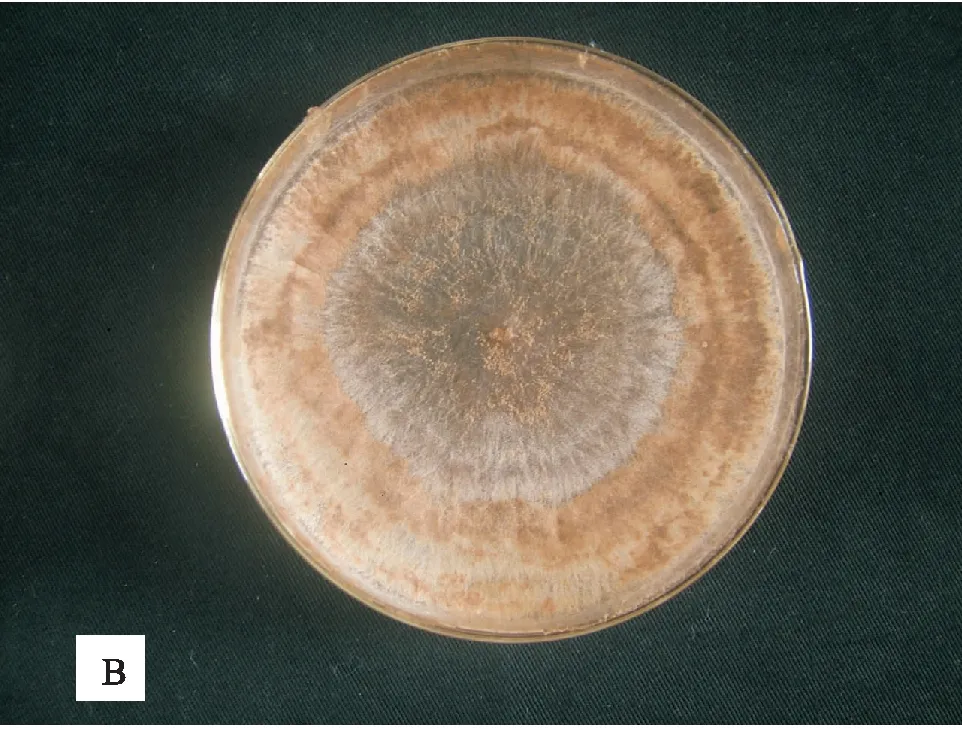
B 金钱莲菌根真菌后期菌落图1 金钱莲根真菌菌落特征
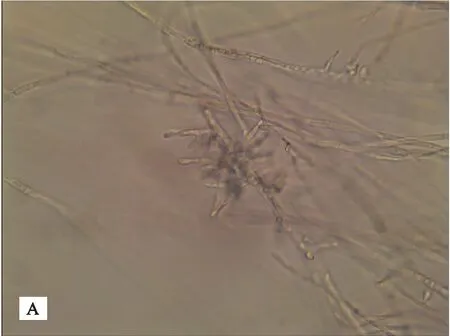
A 幼嫩菌丝

B 老熟菌丝

C 菌核图2 金线莲菌根真菌菌株的显微形态
2.2 ITS序列分析及种类鉴定
各菌株的基因组DNA经PCR扩增,回收后,通过序列测定得到分别为668bps~686bps的各菌株的ITS碱基序列,见表1。
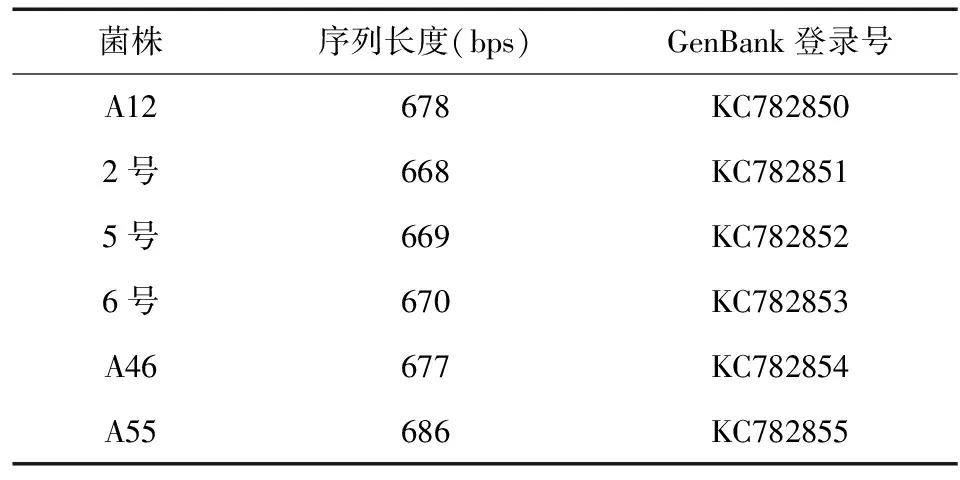
表1 菌株rDNA ITS区序列测定结果
2.3 基于ITS序列的同源性比较
在GenBank中与已有的相关菌株的ITS序列进行同源性比较,用MAGA(3.1)软件将以上菌株与来自GenBank的27株真菌一起构建基于ITS序列的系统发育树。发育树的构建,以亲缘关系相对较远的低等真菌(Pythiumsp.)为外群,见图3。结果显示A12菌株与5号菌株出现在同一最小分支,说明这两菌株的亲缘关系最近;2号菌株与6号菌株出现在同一最小分支,说明这两菌株的亲缘关系最近;A46菌株和A55菌株与前述4株真菌的亲缘关系相对较远,但仍同处于同一较小分支内。所得菌株均与Thanatephorussp.及Rhizoctoniasp.具有较高的同源性。结合各菌株的形态特征和ITS区域基因序列特征,将A12菌株、5号菌株、2号菌株、6号菌株、A46菌株和A55菌株鉴定为丝核菌(Rhizoctoniasp.),其有性态为Thanatephorussp.或Ceratobasidiumsp.。
3 讨论
丝核菌(Rhizoctoniasp.)属的有些成员是农作物的病原菌,如立枯丝核菌(Rhizoctoniasolani)能引起棉花的立枯病[6]和水稻、玉米的纹枯病[7]。也有报道某些种类的丝核菌可与兰科植物共生,利于植株的生长[8]。本研究获得的6株丝核菌从金线莲根部分离,可促进金线莲的生长发育,对其他兰科植物是否具有促生作用,还有待进一步研究。
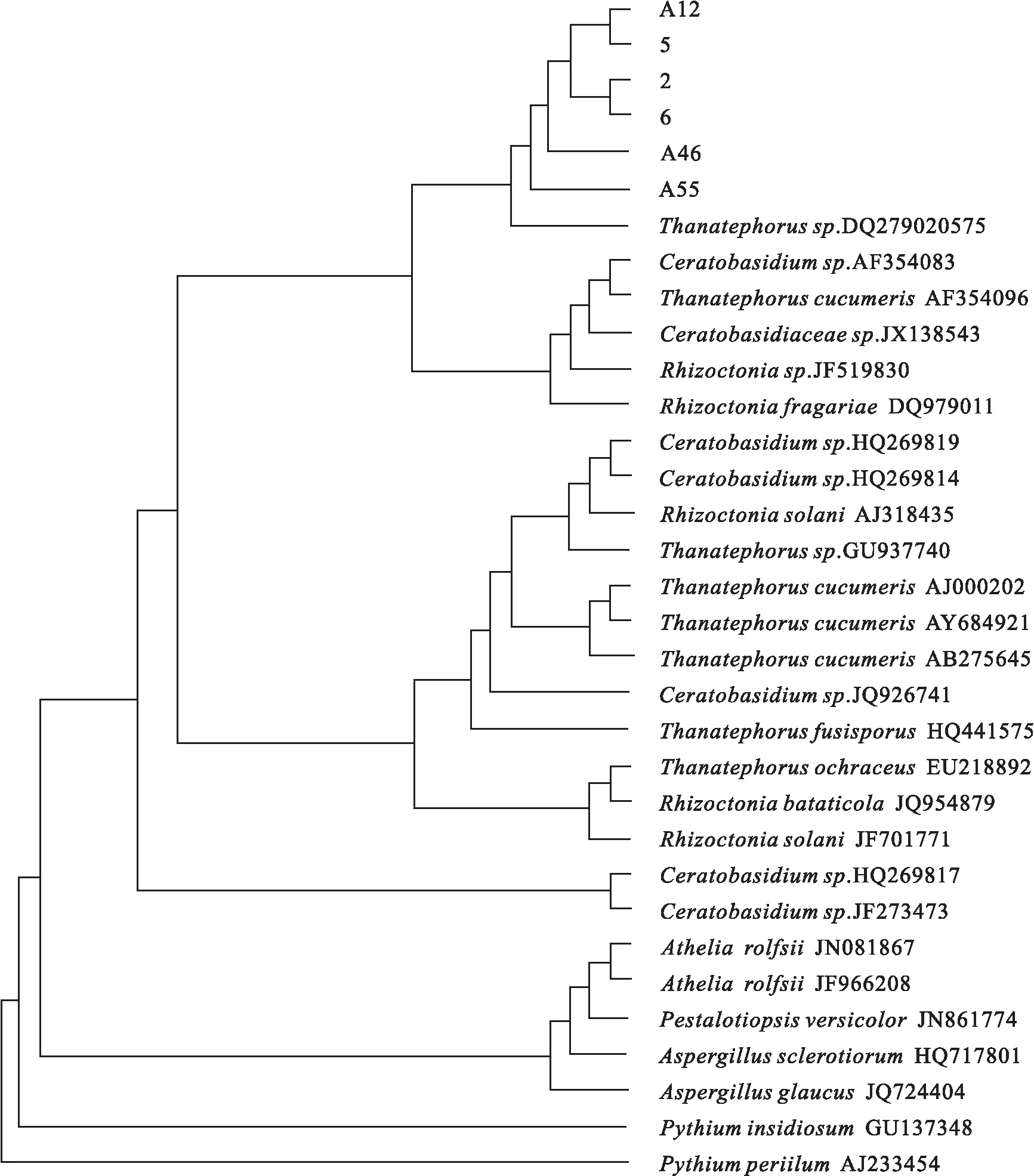
图3 基于ITS序列同源性的各菌株的系统发育树
[1] 谭小明,郭顺星,周雅琴,等.广西金线莲内生真菌的分离及抗菌活性研究[J].中国药学杂志,2010,45(3):178-181.
[2] Huang L, Chen T, Ye Z, et al. Use of liquid chromatography-atmospheric pressure chemical ionization-ion trap mass spectrometry for identification of oleanolic acid and ursolic acid in Anoectochilus roxburghii (wall.) Lindl[J]. J Mass Spectrom, 2007, 42(7): 910-917.
[3] 王雅俊,孟志霞,于雪梅,等.促进福建金线莲生长发育的内生真菌筛选研究[J].中国药学杂志,2009,44(13):976-979.
[4] 郭顺星,陈晓梅,于雪梅.金线莲菌根真菌的分离及其生物活性研究[J].中国药学杂志,2000,35(7):443-445.
[5] Látalová K and Balá M. Carbon nutrition of mature green orchid Serapias strictiflora and its mycorrhizal fungus Epulorhiza sp[J]. Biol Plant,2010, 54: 97-104.
[6] 王伟娟,鹿秀云,李宝庆,等.河北省棉花立枯丝核菌菌丝融合群及其致病性研究[J].华北农学报,2010,25(增刊):274-278.
[7] 冯典兴,郑爱萍,王世全,等.四川省不同寄主立枯丝核菌的遗传分化和致病力研究[J].植物病理学报,2005,35(6):520-525.
[8] 伍建榕,韩素芬,朱有勇,等.春兰与丝核菌共生菌根及结构研究[J].南京林业大学学报(自然科学版),2005,29(4):105-108.
猜你喜欢
小学生作文(低年级适用)(2022年11期)2022-12-02
浙江农林大学学报(2022年5期)2022-10-12
园林科技(2020年2期)2020-01-18
福建农业科技(2018年11期)2019-01-17
中成药(2018年10期)2018-10-26
现代食品(2018年5期)2018-06-06
中国酿造(2017年11期)2017-12-06
化学与生物工程(2017年11期)2017-11-29
中成药(2017年7期)2017-11-22
中国果业信息(2014年2期)2014-01-23
