
伊犁河谷围封草地群落组成及植物多样性的变化
2013-03-14 04:44李军保刘东林吐尔逊娜依热依木江朱进忠
草业科学 2013年5期
李军保,刘东林,吐尔逊娜依·热依木江,朱进忠
(1.陕西省治沙研究所, 陕西 榆林719000;2.新疆农业大学草业与环境科学学院 新疆草地资源与生态重点实验室,新疆 乌鲁木齐 830052)
草地退化已成为新疆地区一个严重的环境问题,春秋牧场的不足和过牧是新疆、尤其是北疆地区草地退化的症结和矛盾所在,退化草地植被的恢复对该区的生态环境与经济发展起着举足轻重的作用。该区的农业历来以草地畜牧业为主,草地畜牧业及其相关产业已成为其支柱产业,而该区生态环境的改善与优化在很大程度上依赖于草地植被的保护、发展与建设。围栏封育作为退化草地植被恢复的一种简单、经济、有效的措施在国内外已得到普遍推广与应用[1-2]。目前,我国开展了一些针对退化草地封育后植被特征变化的研究,结果均表明,封育改善了退化草地植物群落结构组成,增加了植物生物量,封育在退化草地的恢复治理中是有效的[3-8]。我国关于围栏封育方面的报道主要集中在宁夏盐池地区、内蒙古草原和青海湖地区,而对新疆地区围栏封育研究的报道较少。本研究以伊犁河谷昭苏马场春秋放牧地为研究对象,对不同围封年限草地植物群落组成和物种多样性进行调查分析,并探讨退化草地生态系统自然恢复过程中群落组成及其物种多样性的演替规律,以期为更加科学有效地开展该区退化草地植被恢复提供依据。
1 材料与方法
1.1研究区概况 研究区位于伊犁河谷西南部昭苏县的昭苏马场,81°03′~81°05′ E,40°55′~43°15′ N,属温带山区半湿润半干旱气候。年均气温2.7 ℃,≥10 ℃年积温1 416.8 ℃·d,无霜期85~100 d,年均降水量512 mm,降水集中于5-8月,约占全年总降水量的68%,年均蒸发量1 261.6 mm,是降水量的2.5倍,积雪期158 d,积雪厚度20~60 cm,土壤为黑钙土。研究区选在昭苏县境内特克斯河北岸的河滩阶地春秋放牧地,该区属典型的针茅(Stipacapillata)+杂类草草甸草原,主要植物有针茅、草原苔草(Carexliparocarpos)、阿克苏黄耆(Astragalusakeuensis)、黄花苜蓿(Medicagofalcata)、草原糙苏(Phlomispratensis)以及异株百里香(Thymusmarschalianus)等。由于研究区草地退化严重,分别于2002年、2003年春季和2006年秋季实行围栏封育措施,围封前均为放牧草地,未围封的地段照常放牧[9]。
1.2植被调查 于2006 年 7月,对2003年春季围封草地(围封4年)和未围封草地(未围封2006)进行调查,2007年7月对2006年秋季围封草地(围封1年)、2003年春季围封草地(围封5年)、2002年春季围封草地(围封6年)和未围封草地(未围封2007)进行调查,各样地随机设3条样带,样带间间距约50 m,在各样带设50个50 cm×50 cm小样方,间距约20 m,每样地有50个小样方。测定样方内各植物种类的盖度、频度、高度和生物量。盖度采用目测法测定;频度采用样圆法(直径35.6 cm),重复50次;生物量采用齐地面刈割法测定;高度量取自然高度,禾本科、豆科、莎草科草类为优良牧草。
1.3数据分析
物种重要值[10-12]:IV=(RC+RF+RY)×100/3;
丰富度指数[10-12]:S=n;
多样性指数[10-12]:H=∑PilnPi;
均匀度指数[10-12]:J=(-∑PilnPi)/ln S;
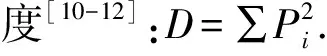
式中,RC表示相对盖度,RF表示相对频度,RY表示相对生物量(鲜质量);n表示样方内出现的物种总数;Pi为种i的相对重要值,Pi=Ni/N(Ni为种i的绝对重要值,N为种i所在样方各个种的重要值之和)。
Sorensen相似性指数[10-12]:Cs=2c/(a+b).
式中,c为两样地或群落共有物种数,a和b分别为样地A和B的物种数。
2 结果
2.1不同围封年限草地植物群落变化 对退化草地的围封消除了放牧的干扰,为植物生长发育和其他已消退物种的侵入营造了良好的生境,使群落的物种组成和结构发生变化。草地植被在自然恢复过程中,群落盖度、生物量和平均株高的增加变化很明显,围封4、5、6年草地与围封1年和未围封草地间差异显著(P<0.05)。围封第1年草地生物量变化很大,显著高于未围封草地生物量(P<0.05),约是未围封草地的3倍。围封6年草地生物量略有下降,但优良牧草的比例呈增加趋势。未围封草地草群低矮,几乎没有垂直结构,且生长季节不能形成华丽的开花季相,受放牧采食和践踏的影响,植物的生长和发育均受到抑制,不能达到最佳生长状态,其生物量和物种数最低(表1)。

表1 不同围封年限草地植物群落特征Table 1 Properties of plant community at different enclosed stages
2.2围封对退化草地植物群落物种组成和结构的影响 随着围封草地的自然恢复及其生境条件的改善,草地植物群落由未围封的异株百里香+草原苔草逐步向适应当地气候条件的针茅+草原苔草方向演进。针茅的重要值增幅最大,由未围封的11.82、12.19增加到围封6年的30.48,成为群落的优势种,而未围封草地由于正常放牧,针茅草丛生长低矮,枝叶量较少,株丛营养不足,分配不均,造成株丛中央老蘖的死亡率加大,导致大部分株丛中心死亡和破碎,制约了针茅的正常生长发育。围封过程中异株百里香的重要值逐渐减小,由未围封草地的优势种退至次要地位,但围封1年变化较小,这与其在围封前有较多个体有关(图1)。
羊茅(Festucaovina)、洽草(Koeleriacristata)和草地早熟禾(Poapratensis)属于根系较浅的小丛禾草,围封后与其他植物在光照和土壤水分养分的竞争中处于不利地位,所以在围封草地植物群落中逐渐减少甚至消失;草原糙苏和千叶蓍(Achilleamillefolium)因其适口性差受放牧干扰较小,在未围封草地生长发育较好,围封后生长发育受到其他种群的竞争抑制,重要值减小;根茎型禾草具有较强的根茎繁殖能力,围封草地土壤物理性状的改善,有利于根茎的生长和营养物质的贮存,因此,拂子茅(Calamagrostisepigejos)重要值增加较无芒雀麦(Bromusinermis)明显,黄花苜蓿在未围封草地为应对牲畜采食,分枝较多且匍匐生长,围封后分枝数减少并直立生长,其生物量的增加补偿了盖度的减小,所以重要值变化不明显;草原苔草为根茎疏丛型草本植物,因植株低矮家畜不便采食,且返青早生长快,植株贮存了大量营养,在春季放牧开始时已进入生殖生长期,植株生长缓慢,所以家畜的采食对草原苔草的影响较小,因而在未围封草地其重要值较大。随着围封草地的自然恢复,草原苔草的重要值随种间竞争的增大逐渐减小,但变化不明显,说明草地恢复演替对其影响较小,这也体现了草原苔草在草地群落中伴生种的地位(图1)。
根据组成群落的各种植物在群落中的表现,可以将其变化趋势归纳为衰退型、增长型和波动型。

图1 植物种重要值消长趋势Fig.1 The variation trend in important values of plant species
衰退型:特点为随围封时间的延长,优势度降低,如异株百里香、草地早熟禾、羊茅、洽草、草原糙苏和千叶蓍。这些植物种耐牧性强,能够忍受较强的干旱及家畜的采食和践踏,常出现在资源贫瘠的生境中,大部分能量用来维持和延长寿命,从生态对策来看属于S对策种。
增长型:特点为随放围封时间的延长重要值升高,如针茅、无芒雀麦和拂子茅等。他们多为高大禾本科植物,能够充分利用群落空间资源环境而在种间竞争中处于优势地位,属于C对策种。
波动型:在草地围封群落演替过程中,种群重要值在各群落中有起伏,但总体上保持稳定,属于恒有种,如银背委陵菜(Potentillaargenta)、森林草莓(Fragariavesa)、黄花苜蓿、大车前(Plantagemajor)等。多属群落伴生种,生态适应性较强,种群变化对群落演替影响不大。
2.3植物经济类群的变化 草地植物经济类群是以草地植物的自然和经济特性为依据,综合考虑其饲用价值、生态价值与经济价值来划分,可分为禾本科、豆科、莎草科、菊科、藜科和杂类草六大类[2,13](表2)。
草地植被围封恢复过程中,禾本科增加了无芒雀麦、拂子茅和鹅观草(Elymuskamoji),而草地早熟禾在群落中很快退出。禾本科重要值增加幅度较大,围封6年的重要值是未围封的2.2~2.5倍,说明围封草地中禾本科牧草在群落中的资源利用能力、竞争力等较强。白花车轴草(Trifoliumrepens)和葫芦巴(Trigonellafocnum-graecum)的消失使围封草地群落豆科类减少,但其重要值并没有减小而是逐渐增加;菊科变化不大,重要值较小,随着围封年限的延长呈减小趋势,在群落中处于次要地位;藜科和杂类草逐渐增加且变化较大,重要值减小。表明退化草地围封自然恢复促进了针茅等禾草的生长发育,抑制了异株百里香、草原糙苏、千叶蓍等杂类草的生长发育。
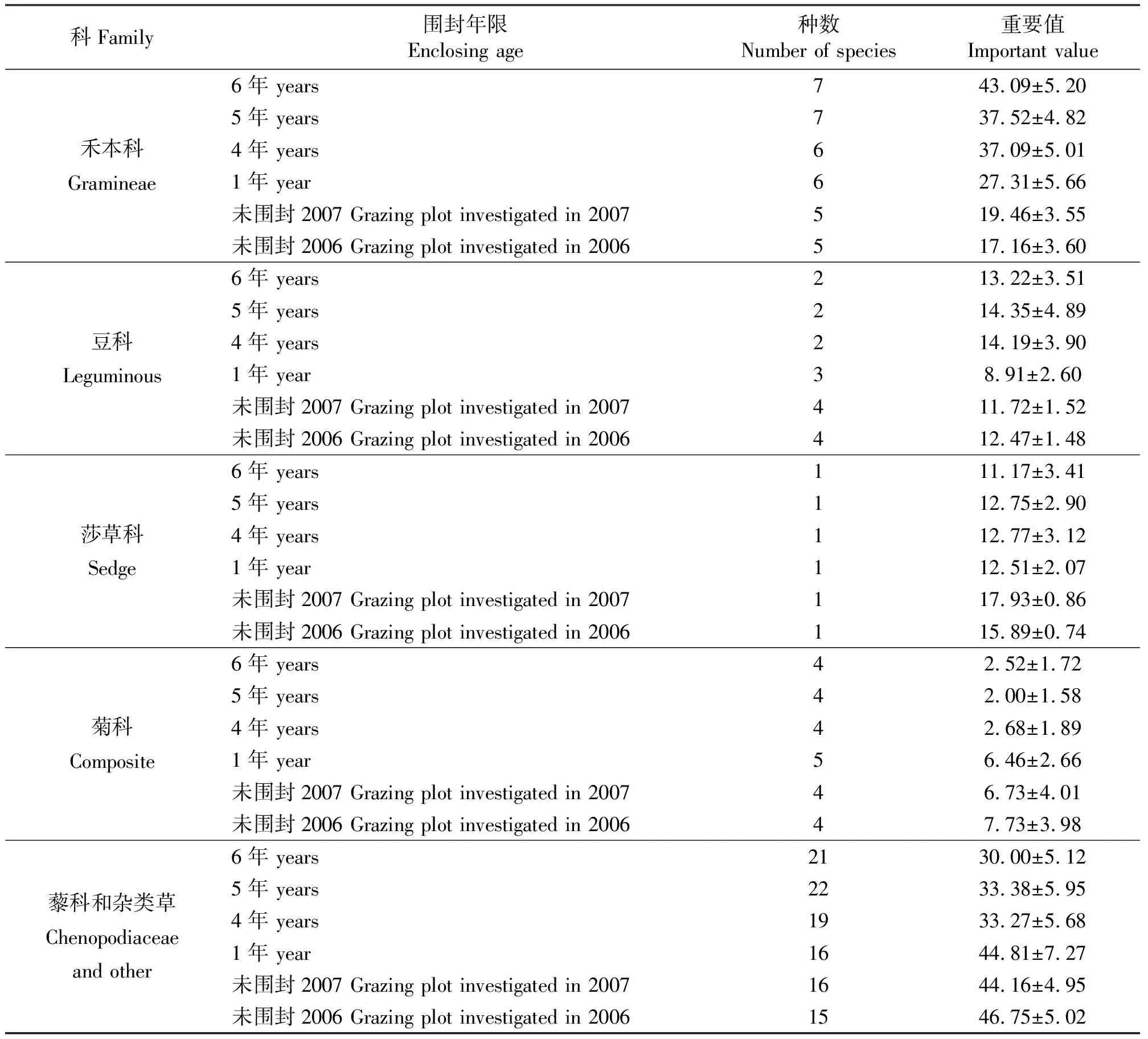
表2 植物经济类群重要值变化Table 2 Important value change of economic herbage group
2.4围封对草地植物群落物种多样性的影响 植物多样性恢复是植被恢复的直接体现[13]。物种丰富度随围封年限的延长逐渐增加(表3),分析结果表明,围封6年草地与围封5、4、1年和未围封草地间的差异显著(P<0.05),而围封1、5年和4年草地间的差异不显著(P>0.05)。物种均匀度指数随着围封时间的延长呈减小趋势,其在围封1年时达到最大。
围封恢复草地的主要优势种为针茅,群落优势度随围封年限的增加逐渐增大,表明针茅在群落中的作用逐渐增强;而未围封草地异株百里香较多,其优势度与围封4、1年草地间的差异不显著(P>0.05)。物种多样性指数随着围封年限的增加呈增大趋势,以围封1年的最大,与围封5、6年草地间差异不显著(P>0.05),但与未围封草地之间差异性显著(P<0.05)。因为多样性指数受物种丰富度和均匀度的共同影响,一个具有较低物种丰富度指数和较高均匀度指数的群落,其物种多样性指数可能和一个物种丰富度指数大而均匀度指数低的群落相同[14-15]。
2.5群落的相似性分析 每一群落总是与其围封年限最邻近的群落具有最高的相似度(表4)。围封时间长的草地与未围封草地的相似性指数较低,说明其群落物种组成差异不断增大;围封6年和5年草地的相似性指数为0.95,表明草地恢复到一定阶段后群落的物种趋于稳定;未围封草地物种数受放牧干扰的影响变化较大。不同围封年限草地群落间的相似性指数都大于0.65,群落的相似性程度较高。说明退化草地植被恢复是中途演替过程,群落中主要植物种并没有消失,而是原有植物种的消长变化,这也是退化草地植被短期可恢复的重要原因。

表3 各围封年限草地植物物种多样性指数变化Table 3 Species diversity of plant community in different enclosed stages
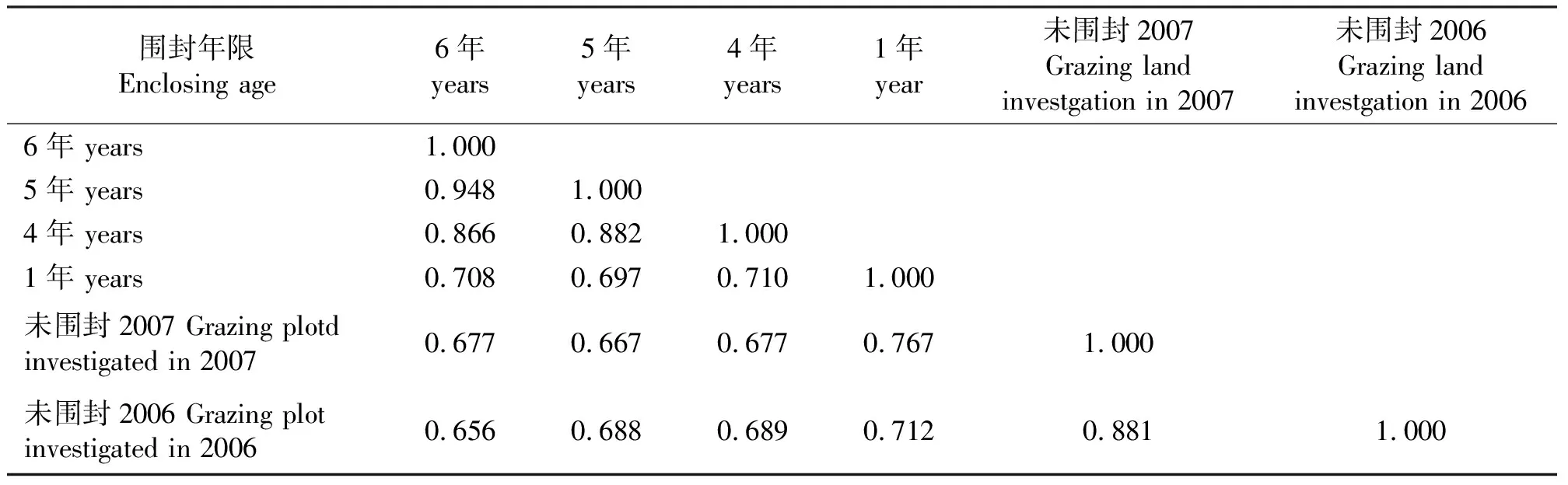
表4 群落相似性指数比较Table 4 Comparison of similarity indices of plant communities
3 讨论
围封措施主要是人为地降低或完全排除家畜对草地生态系统的影响,使草地生态系统在自身弹性下得以恢复和重建[2,6,15]。草地围封恢复过程中,群落物种组成、盖度和生物量均呈现不同的变化特征。
随着草地围封的自然恢复,针茅替代异株百里香成为群落的优势种且数量逐渐增加,在群落中处于优势地位,是群落演替的标识。群落由未围封的异株百里香+草原苔草逐步向适应当地气候条件的针茅+草原苔草方向演进。群落高度、生物量围封5年时最大,之后降低,群落盖度和优良牧草比例继续增加,围封6年时最大。由于地区环境的差异和草地退化程度的不同,植物生物量恢复时间也不尽相同,农牧交错带的草原,封育3年其生物量最大[16];退化羊草草原,围封两年草地上生物量最高[17];适当放牧利用或刈割,可刺激草丛分蘖,促进草丛再生,改良草地[18]。本研究表明,伊犁河谷春秋放牧地退化草地围封5年时,从生产力或生物量的角度上看已适宜再度进行割草或放牧利用。
由于草原类型的复杂性以及围封时间、放牧强度等的差异,对草地恢复过程中多样性研究结果也不尽相同,内蒙古典型草原植被围封26年时,其丰富度指数、多样性指数均为最高,均匀度指数最低[8];随着围封年限的延长,高寒草地植物群落丰富度指数和多样性指数均为在围封1年的草地中达到最大,而后逐渐降低;物种均匀度指数不断增大,优势度指数在未围封草地中最大[19]。物种丰富度和均匀度指数变化趋势并不一致,是因为物种丰富度指群落内种的绝对密度,而物种均匀度指群落内种的相对密度[20],物种丰富度指数与特定群落的物种总数成正比,与个体在群落内的分布无关,而物种均匀度指数则强调个体在群落内的分布,分布越均匀该指数越大。生态优势度指一个种在群落内出现的频率和作用,其值越大表明优势种的作用越大,摄取资源的能力也就越大,其值越小说明群落内种的作用差别不大[21]。本研究表明,随着围封年限的延长,物种丰富度和优势度逐渐增大,多样性指数和物种均匀度以围封1年的最大。由于禾本科牧草春季返青较早而杂类草较晚,围封消除放牧干扰,客观上增强了禾本科牧草对杂类草在资源上的竞争能力,有利于禾本科牧草的生长发育,其在群落的重要值逐渐增加,抑制了藜科植物和杂类草的生长发育,这与前人的研究结果[7,22-23]相似。群落的相似性与其围封年限邻近的群落有最高的相似度,围封6年与围封5年草地的相似性最大,说明草地恢复到一定阶段群落的物种趋于稳定,未围封草地受放牧干扰影响物种变化波动较大,与围封草地群落的相似性指数较小。
[1] 王小利,干友民,张力,围栏内禁牧与轻牧对高寒草原群落的影响[J].甘肃农业大学学报,2005,40(3):368-375.
[2] 李军保,朱进忠,吐尔逊那依·热依木江,等.围栏封育对昭苏马场春秋草场植被恢复的影响[J].草原与草坪,2007(6):45-48.
[3] Meissner R A,Facelli J M.Effects of sheep exclusion on thesoil seed bank and annual vegetation in chenopods shrublandsof south Australia[J].Journal of Arid Environments,1999,42:117-128.
[4] 李政海,王炜,刘钟龄.退化草原围封恢复过程中草场质量动态的研究[J].内蒙古大学学报(自然科学版),1995,26(3):334-338.
[5] 杨晓晖,张克斌,侯瑞萍,等.封育措施对半干旱沙地草场植被群落特征及地上生物量的影响[J].生态环境,2005,14(5):730-734.
[6] 李永宏.内蒙古典型草原地带退化草原的恢复动态[J].生物多样性,1995,3(3):125-130.
[7] 闫玉春,唐海萍,常瑞英,等.典型草原群落不同围封时间下植被、土壤差异研究[J].干旱区资源与环境,2008,22(2):145-151.
[8] 周国英,陈桂深,赵以莲,等.施肥和围栏封育对青海湖地区高寒草原影响的比较研究Ⅰ群落结构及其物种多样性[J].草业学报,2004,13(1):26-31.
[9] 李军保,马存平,刘洪来,等.围封对昭苏马场春秋草场种群生态位的影响[J].中国草地学报,2009,31(2):86-91.
[10] 孙涛,毕玉芬,赵小社.围栏封育下山地灌草丛草地植被植物多样性与生物量的研究[J].云南农业大学学报,2007,22(2):246-250,279.
[11] 王小利,张力,张德罡,等.青海湖地区线叶嵩草型中度与重度退化草地群落的比较研究[J].草业科学,2006,23(10):8-13.
[12] 张继义,赵哈林,张铜会,等.科尔沁沙地植被恢复系列上群落演替与物种多样性的恢复动态[J].植物生态学报,2004,28(1):86-92.
[13] Fang W,Peng S L.Development of species diversity in the restoration process of establishing a tropical man-made forest ecosystem in China[J].Forest Ecology and Management,1997,99:185-196.
[14] 阎桂琴,赵桂仿,胡正海.秦岭太白红杉群落特征及其物种多样性的研究[J].西北植物学报,2001,21(3):497-506.
[15] 王明玖,李青丰,青秀玲.贝加尔针茅草原围栏封育和自由放牧条件下植物结实数量的研究[J].中国草地,2001,23(6):21-26.
[16] 李瑞,张克斌,王百田,等.农牧交错带不同封育时间对植物特征值及多样性的影响[J].干旱区资源与环境,2007,21(7):106-110.
[17] 左万庆,王玉辉,王风玉,等.围栏封育措施对退化羊草草原植物群落特征影响研究[J].草业学报,2009,18(3):12-19.
[18] 程积民,邹厚远,Akio H.封育刈割放牧对草地植被的影响[J].水土保持研究,1998,5(1):36-54.
[19] 胡玉昆,高国刚,李凯辉,等.巴音布鲁克草原不同围封年限高寒草地植物群落演替分析[J].冰川冻土,2009,31(6):1186-1194.
[20] 岳天祥.生物多样性研究及其问题[J].生态学报,2001,21(3):462-467.
[21] 郭正刚,梁天刚,刘兴元,等.新疆阿勒泰地区草地类型及植物多样性的研究[J].西北植物学报,2003,23(10):1719-1724.
[22] 刘忠宽,汪诗平,陈佐忠,等.不同放牧强度草原休牧后土壤养分和植物群落变化特征[J].生态学报,2006,26:2048-2056.
[23] 李青云,董全民.围栏封育对高寒草甸退化植被的作用[J].青海草业,2002,11(3):1-3.
猜你喜欢
天津农林科技(2022年2期)2022-04-19
河南畜牧兽医(2022年3期)2022-04-13
绿色科技(2021年10期)2021-06-23
今日农业(2020年19期)2020-12-14
河南畜牧兽医(2020年21期)2020-01-10
草业学报(2019年2期)2019-02-25
北京航空航天大学学报(2017年3期)2017-11-23
中国科技信息(2015年2期)2015-11-16
中国药业(2014年17期)2014-05-26
植物营养与肥料学报(2014年1期)2014-03-11
