
白桦人工林土壤主要养分元素时空变异1)
2013-03-05 09:58赵淑苹庞凤艳
东北林业大学学报 2013年5期
孟 春 罗 京 赵淑苹 庞凤艳
(森林持续经营与环境微生物工程黑龙江省重点实验室(东北林业大学),哈尔滨,150040) (哈尔滨市林业科学研究院)
土壤养分是土壤肥力的重要物质基础,它不仅是植物营养元素的主要来源,同时也是影响土壤与空气间气体交换(尤其是CO2)的重要因素。因此,国内外学者多年来对土壤营养元素及时空变异进行了大量的研究[1-2]。对于土壤营养元素的空间变异性,研究者多采取对不同利用类型和不同深度(层次)土壤进行取样来讨论其空间变异性[3-6];对于土壤营养元素的时间变异性,研究者多以年为时间跨度来讨论其时间变异性[7-11]。对不同利用类型和不同深度(层次)土壤进行取样研究,可以较好地揭示土壤营养元素的空间变异,而以年为时间跨度研究土壤营养元素的时间变异,则忽视了植物生长季节性变化对土壤营养元素的影响。本文以白桦(Betula platyphylla Suk.)人工林地为研究对象,以月为时间跨度,并结合前人对该林地土壤营养元素的研究结果来揭示土壤营养元素的时空变异性。
1 材料与方法
1.1 样地概况
东北林业大学哈尔滨实验林场位于哈尔滨市区内马家沟河西岸(N45°43. 463' ~N45°43. 464',E126°37.457' ~E126°37.458'),地形平缓,土壤为地带性黑土,水分条件良好。属于温带季风性气候,年平均气温3.5 ℃,年积温2 757 ℃,年降水量534 mm。
本次试验选取该林场内白桦(Betula platyphylla Suk.)人工林为研究样地。该林地于1960年春季以2年实生苗造林,初植密度0.5 m×1 m,当年成活率95%。前3年按常规方法进行除草、抚育等田间管理。1964年隔一行去一行,变株距为1 m ×1 m,1990年进行疏伐。目前,每0.5 hm2林地上活立木493 株,平均高19 m,平均胸径16 cm,活立木蓄积53.81 m3。下草盖度20%。
1.2 观测方法
在选定样地上,以对角线形式随机选取5 个取样点,在每个取样点用铁锹挖出深约50 cm 的土壤剖面。分别于2011年5月15日、6月24日、8月8日、9月15日和10月2日分5 次在每个剖面上从下向地表分别在45、30、15、5 cm 处取样,土样编号后放入取样袋中,带回室内进行自然风干、粉碎、过筛,制备土样供测定土壤化学元素之用。土壤全N 采用半微量凯氏法测定,土壤水解性N 采用碱解—扩散法测定,土壤全P 采用氢氧化钠—钼锑抗比色法测定,土壤有效性P 采用0.025 mol/L 硫酸浸提法测定,土壤全K、速效性K 采用火焰光度法测定,土壤C 元素采用水合热法测定,土壤pH 值采用水浸电位法测定。除K 元素进行4 次测定外,其他各元素均进行了5 次测定。
1.3 数据统计
采用经典统计方法计算每个样地每次取样同层次各项指标的平均值、标准差。采用变异系数(CV)说明各项指标的变异程度:CV=S/(S 为标准差;为均值)。CV≤0.1 属于弱变异性,0.1 <CV<1 属于中等变异性,CV≥1 属于强变异性[12]。采用方差分析说明各指标时空差异的显著性。
2 结果与分析
2.1 N 元素
观测期内土壤全N 质量分数平均值为1 101.74 mg·kg-1(表1),低于1983年该林地土壤全N 观测值[13]。土壤全N 质量分数虽然不同土层深度处差异性并不显著(p >0.05),即全N 质量分数最低的45 cm 土层处较最高的5 cm 土层处仅低了17.44%,但全N 却呈现随土层深度增加而减小的变化趋势,这种状况与陈喜全[13]对该林地土壤全N沿土层深度方向变化趋势的研究结果完全相同。各次取样间土壤全N 质量分数差异性显著(p <0.05)。8月8日和9月15日两次取样中各层次土壤全N 质量分数较低,其余3 次取样中各层次土壤全N 质量分数较高。全N 质量分数最低的8月8日较最高的6月24日低了40.81%。观测期内土壤全N质量分数平均变异系数为0.37,属中等变异。变异系数随土层深度的增加有减小的变化趋势,各次取样间变异系数无明显规律性变化。
观测期内土壤水解性N 质量分数平均值为360.89 mg·kg-1(表1)。虽然不同土层深度处土壤水解性N 质量分数差异并不显著(p >0.05),即水解性N 质量分数最低的30 cm 土层处比质量分数最高的45 cm 土层处仅低了3.91%,但却呈现随土层深度的增加先减小后增加的变化趋势。各次取样间土壤水解性N 质量分数差异性显著(p <0.05):5月15日和10月2日两次取样的土壤水解性N 质量分数较高,其余3 次取样的土壤水解性N 质量分数较低。水解性N 质量分数最低的8月8日较质量分数最高的5月15日低了40.31%。观测期内土壤水解性N 质量分数平均变异系数为0.36,属中等变异。变异系数随土层深度的增加有先增加后减小的变化趋势,各次取样间变异系数呈现先增加后减小的变化趋势。
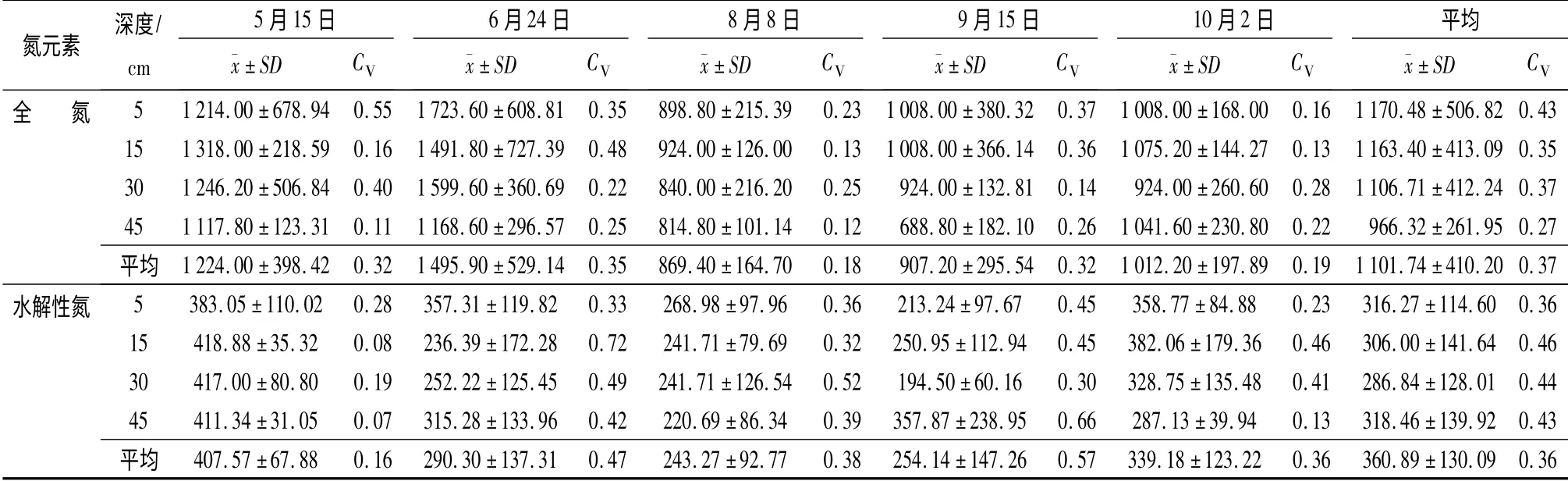
表1 土壤N 元素统计值
2.2 P 元素
观测期内土壤全P 质量分数平均值为249.11 mg·kg-1(表2),低于1983年该林地土壤全P 的观测值[13]。土壤全P 质量分数在不同深度处差异性不显著(p >0.05),无明显变化趋势。全P 质量分数最小值(45 cm 土层深度处)较最大值(30 cm 土层深度处)小了20. 91%,这一结果与陈喜全1983年[13]对该林地土壤全P 沿土层深度方向的变化趋势的研究结果基本相同。各次取样间土壤全P 质量分数差异性显著(p <0.05):从5月15日第1 次取样至10月2日第5 次取样,土壤全P 质量分数呈现先减小后增加的变化趋势。9月15日取样的土壤全P 质量分数较5月15日取样的土壤全P 质量分数减少了44.81%。观测期内土壤全P 质量分数平均变异系数为0.34,属中等变异。变异系数随土层深度的增加呈先减小后增加的变化趋势,各次取样间变异系数无明显的规律性变化。
观测期内土壤速效P 质量分数平均值为4.12 mg·kg-1(表2)。不同土层深度处土壤速效P 质量分数差异显著(p <0.05),呈现随土层深度的增加而减小的变化趋势,45 cm 土层深度处速效P 质量分数较5 cm 土层深度处减小了53.69%。各次取样间土壤速效P 质量分数差异性显著(p <0.05):5月15日取样时速效P 的质量分数最低;6月24日至10月2日,速效P 则呈现先减小后增加的变化趋势。土壤速效P 质量分数最低的5月5日较最高的6月24日低了65.93%。观测期内土壤速效P 质量分数平均变异系数为0.63,属中等变异。变异系数随土层深度的增加有增加的趋势,各次取样间变异系数表现为先增加后减小的变化趋势。
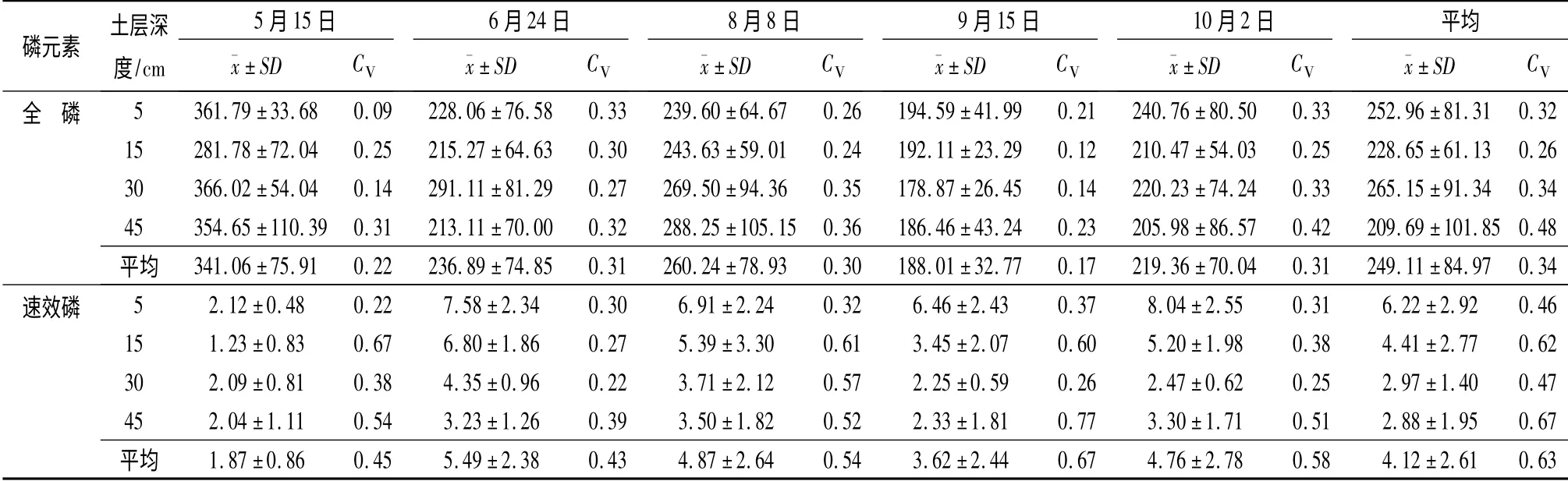
表2 土壤P 元素统计值
2.3 K 元素
观测期内土壤全K 质量分数平均值为803.60 mg·kg-1(表3)。不同深度处土壤全K 质量分数差异性虽不显著(p >0.05),但全K 质量分数呈现随土层深度增加而减小的变化趋势,最小值(45 cm土层深度处)比最大值(5 cm 土层深度处)小了5.72%。各次取样间土壤全K 质量分数差异性不显著(p >0.05),土壤全K 质量分数最低的9月15日较最高的6月24日低了7.32%。从6月24日第1次取样至10月2日第4 次取样,土壤全K 质量分数呈现出先减小后增加的变化趋势。观测期内土壤全K 质量分数平均变异系数为0.11,属中等变异。变异系数随土层深度的增加呈现先增加后减小的变化趋势,各次取样间变异系数亦呈现先增加后减小的变化趋势。
观测期内土壤速效K 质量分数平均值为67.42 mg·kg-1(表3)。不同土层深度上土壤速效K 质量分数差异性不显著(p >0.05),但呈现出随土层深度的增加而增加的变化趋势,最大值(45 cm 土层深度处)比最小值(15 cm 土层深度处)大了21.90%。各次取样间土壤速效K 质量分数差异性不显著(p >0.05),各次取样土壤速效K 质量分数平均值差异不大,最大值(8月8日)较最小值(6月24日)仅增加了9.61%,但有先减小后增加的变化趋势。观测期内土壤速效K 质量分数平均变异系数为0.21,属中等变异。变异系数随土层深度的增加表现为先增加后减小,而在各次取样间变异系数则表现为先减小后增加。
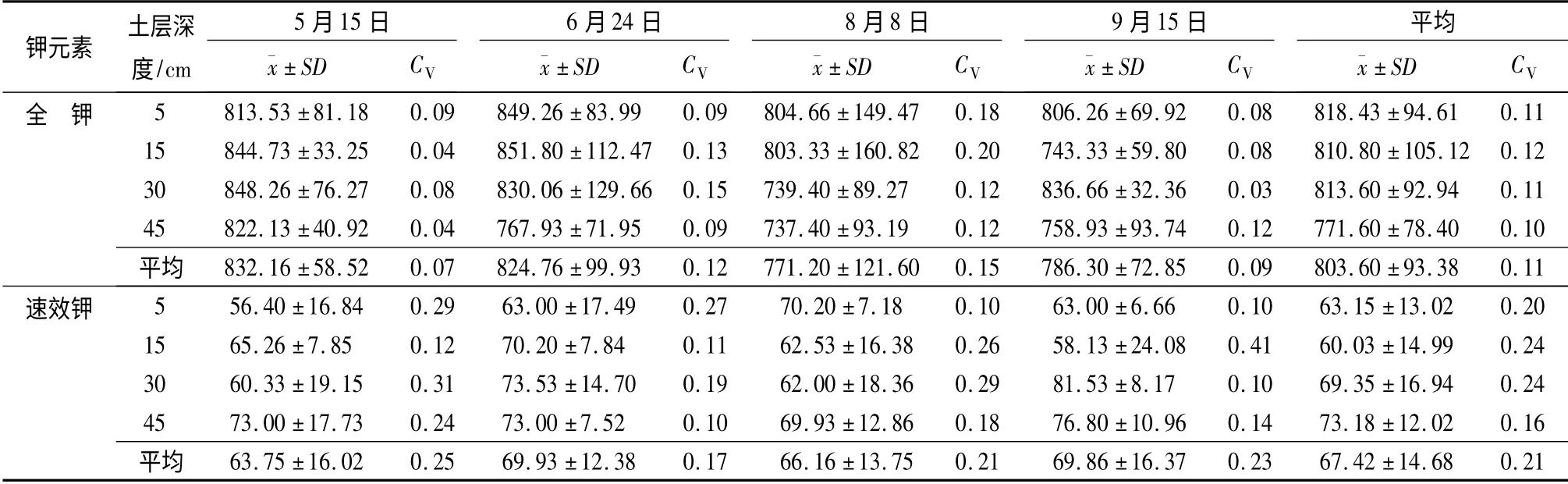
表3 土壤K 元素统计值
2.4 C 元素
观测期内土壤C 元素质量分数平均值为18.63 g·kg-1(表4),略高于1983年[13]该林地土壤C 元素的观测值。不同土层深度处土壤C 元素差异性显著(p <0.05),最小值(15 cm 土层深度处)比最大值(30 cm 土层深度处)小了19.29%,表现出随土层深度的增加先减小后增加的波动变化趋势,这与陈喜全[13]对该林地土壤C 元素的观测值随土层深度变化的趋势有相似之处。各次取样间C 元素差异性不显著(p >0.05):从5月15日第1 次取样至10月2日第5 次取样,土壤C 元素质量分数呈现递增的变化趋势,10月2日取样的土壤C 元素质量分数较5月15日取样时增加了12.50%。观测期内土壤C 元素平均变异系数为0.21,属中等变异。变异系数随土层深度的增加波动变化,各次取样间变异系数呈减小的变化趋势。

表4 土壤C 元素统计值
2.5 pH 值
观测期内土壤pH 值平均值为5.65(表5),低于1983年该林地土壤pH 值的观测值[13]。各层pH值虽然变化很小,即pH 最小值(15 cm 土层深度处)比最大值(45 cm 土层深度处)仅小了5.46%,但不同土层深度处土壤pH 值差异性显著(p <0.05),并表现出随土层深度增加先减小后增加的变化趋势,这与陈喜全[13]对该林地土壤pH 值随土层深度变化的趋势略有不同。各次取样间土壤pH 值差异性显著(p <0.05):从5月15日第1 次取样至10月2日第5 次取样,土壤pH 值呈现先减小后增加的变化趋势,最小值(8月8日)较最大值(5月15日)减小了7.44%。观测期内土壤pH 值平均变异系数为0.05,属弱变异。变异系数随土层深度的增加有增加的趋势,在各次取样间变异系数则表现为先减小后增加。

表5 白桦林地土壤pH 值统计结果
3 结论与讨论
C 元素是构成植物有机体的基本物质。通过光合作用,植物将大气中的CO2合成为有机物,以碳水化合物的形式构成植物各个器官的化学组分。同时,植物正常生长需要吸收必要的营养元素。这些营养元素除N 元素可以从大气中吸收一部分外,N元素的大部分和其他营养元素如P、K 的吸收均来自土壤。因此,研究样地从1960年造林开始至今,尽管每年有枯枝落叶分解而致营养元素回归,但土壤主要养分元素一直处于“净支出”状态,地力有“衰退”的趋势[14],在有对比数据的几个土壤营养元素中,土壤全N、全P 质量分数均较1983年的观测值下降。C 元素得益于光合作用的积累,通过细根的分解[15-16]而使土壤C 元素质量分数增加。pH值的下降使林地土壤从中性转变为弱酸性,白桦林出现类似情况的不多[17],其原因可能是由于该林地与落叶松林地相邻,落叶松落叶进入白桦林地后分解造成土壤酸性化[18],从pH 值在不同土层深度处差异性显著及随土层深度增加先减小后增加的变化趋势也可以印证这一原由。
土壤营养元素在土壤剖面垂直方向上质量分数的变化既与成土过程有关,也与植物生长、枯落物和残体的分解以及不同元素的特性有关,所以,即使土壤类型相同,但利用方式不同,土壤营养元素在土壤剖面垂直方向上质量分数的变化趋势也会明显不同。相对于1983年而言,经过近30a 的生长,白桦林已从中龄林生长为成熟林,生长速度降低、根系发达。此时,分解周期极短的细根成为影响土壤营养元素变化的一个重要因素[19]。在细根分解过程中,对于土壤有机质、全N、全P、全K 而言是个增值的过程[20]。有研究表明[16],白桦林细根的垂直分布表现出随土层深度的增加细根生物量减少的趋势,92.8%和93.1%的活、死根分布在0 ~20 cm 的土层中。因此,土壤全N、水解性N、全P、速效P 和全K质量分数表现出的随土层深度增加而减小的变化趋势与白桦细根的垂直分布特点相符合。速效K 则由于其对植物体新鲜程度的选择性而使其在根部质量分数较小,出现与全K 相反的变化趋势。C 元素则表现为在20 cm 土层以下的质量分数高于0 ~20 cm 土层的质量分数,这与不同土层深度活细根与死细根的比例以及淋溶作用产生的沉积作用有关。
N、P、K 元素作为植物体的营养元素,生长初期(6月份)在植物体内都有较高的积累[21],导致经7、8月份植物速生期的波动变化后土壤中N、P、K 元素质量分数降低,至非主要植物生长季节的9、10月份其质量分数又上升。因此,全N、全P、全K 以及水解性N、速效K 和速效P 均表现出了与植物生长对营养元素的季节性吸收特点相一致的变化趋势。C 元素的月际变化呈现增加的趋势与细根分解导致土壤C 元素的积累直接相关。pH 值的月际变化则与细根的生长及分解过程中微生物的活性有关,微生物活性高的月份pH 值较低。
研究样地是经由松花江、阿什河的长期侵蚀、搬运和堆积作用下形成的复微度分割的冲积平原[13],土壤和营养元素的空间差异不大,研究样地在栽植白桦纯林后,由于树种单一,对土壤营养元素的空间差异影响甚微,因此,除pH 值为弱变异外,其他各元素也仅为中等变异,林地土壤均值性较好。
[1] 黄绍文,金继运. 土壤特性空间变异研究进展[J]. 土壤与肥料,2002(1):8 -14.
[2] 冯晓,乔淑,胡峰,等.土壤养分空间变异研究进展[J].湖北农业科学,2010,49(7):1738 -1741.
[3] Li X F,Chen Z B,Chen H B,et al. Spatial distribution of soil nutrients and their response to land use in eroded area of south China[J]. Procedia Environmental Sciences,2011,10(A):14 -19.
[4] Wei Yichang,Bai Youlu,Jin Jiyun,et al. Spatial variability of soil chemical properties in the reclaiming marine foreland to Yellow Sea of China[J]. Agricultural Sciences in China,2009,8(9):1103 -1111.
[5] 于洋,刘吉平,徐艳艳.东北典型黑土区土壤养分空间分异影响因素分析[J].水土保持研究,2009,16(5):66 -69.
[6] 李双异,刘慧屿,张旭东,等.东北黑土地区主要土壤肥力质量指标的空间变异性[J].土壤通报,2006,37(2):221 -227.
[7] Huang Biao,Sun Weixia,Zhao Yongcun,et al. Temporal and spatial variability of soil organic matter and total nitrogen in an agricultural ecosystem as affected by farming practices[J]. Geoderma,2007,139(3/4):336 -345.
[8] Yang Z P,Zhang Q,Wang Y L,et al. Spatial and temporal variability of soil properties under Caragana microphylla shrubs in the northwestern Shanxi Loess Plateau,China[J]. Journal of Arid Environments,2011,75(6):538 -544.
[9] Prasolova N,Xu Zhihong,Saffigna P G,et al. Spatial-temporal variability of soil moisture,nitrogen availability indices and other chemical properties in hoop pine (Araucaria cunninghamii)plantations of subtropical Australia[J]. Forest Ecology and Management,2000,136(1/3):1 -10.
[10] Cui Baoshan,Zhao Hui,Li Xia,et al. Temporal and spatial distributions of soil nutrients in Hani terraced paddy fields,Southwestern China[J]. Procedia Environmental Sciences,2010,2:1032 -1042.
[11] 汪景宽,李双异,张旭东,等.20年来东北典型黑土地区土壤肥力质量变化[J].中国生态农业学报,2007,15(1):19 -24.
[12] 刘淑娟,张伟,王克林,等. 桂西北喀斯特峰丛洼地土壤物理性质的时空分异及成因[J]. 应用生态学报,2010,21(9):2249 -2256.
[13] 陈喜全.关于哈尔滨林场土壤发生分类的探讨[J].东北林学院学报,1983,11(3):12 -18.
[14] 崔国发.人工林地力衰退机理及其防止对策[J].世界林业研究,1996(5):61 -69.
[15] 郭忠玲,郑金萍,马元丹,等. 长白山几种主要森林群落木本植物细根生物量及其动态[J].生态学报,2006,26(9):2855-2863.
[16] 汪洋,代力民,于大炮,等. 长白山白桦林细根的生产力及其周转[J].湖南农业科学,2010(7):113 -115.
[17] 王树力,别志远,张先庆,等. 凉水自然保护区4 种红松林型下土壤性质的对比分析[J].林业科技,1997,22(2):1 -3.
[18] 陈喜全,郭秋慧,王政权.落叶松纯林与落叶松胡桃楸混交林下土壤理化性质的研究(Ⅰ)[J]. 东北林业大学学报,1991,19(水胡黄椴专刊):258 -267.
[19] 张秀娟,吴楚,梅莉,等. 水曲柳和落叶松人工林根系分解与养分释放[J].应用生态学报,2006,17(8):1370 -1376.
[20] 邢存旺,张艳华.胡桃楸与落叶松人工混交林土壤化学性质的研究[J].东北林业大学学报,1995,23(5):64 -71.
[21] 李俊清,宫伟光.东北主要造林树种的营养元素含量特征分析[J].植物生态学与地植物学学报,1991,15(4):380 -385.
猜你喜欢
建材发展导向(2022年10期)2022-07-28
农业工程技术(2021年28期)2021-12-30
当代陕西(2020年24期)2020-02-01
当代陕西(2020年24期)2020-02-01
当代陕西(2020年24期)2020-02-01
现代园艺(2017年23期)2018-01-18
现代园艺(2017年21期)2018-01-03
绿色科技(2016年16期)2016-10-11
亚热带资源与环境学报(2015年1期)2015-01-22
水土保持研究(2014年4期)2014-09-21
