
宁波地区蔬菜主要病原线虫的分离和种类鉴定
2013-02-28 07:48姚红燕顾建锋王江岭陈若霞
植物保护 2013年6期
姚红燕, 顾建锋, 王江岭, 陈若霞
(1.宁波市农业科学研究院 宁波 315040;2.宁波检验检疫科学技术研究院 宁波 315010)
由植物寄生线虫侵袭和寄生引起的植物病害称为植物病原线虫病。据Esser的统计,到1990年为止全世界已报道发现植物寄生线虫207个属,共4 832种[1]。危害植物的线虫主要有根结线虫、孢囊线虫、滑刃线虫、茎线虫、粒线虫及短体线虫等。据Sasser &Freek man报道,植物寄生线虫每年造成世界农业生产损失约1 000亿美元[2]。其中危害最严重的是根结线虫(Meloidogyne Goeldi,1987),有90余种,而常见的4种根结线虫包括南方根结线虫、北方根结线虫(M.hapl a Chit wood)、爪哇根结线虫[M.j avanica(Treub)]和花生根结线虫[M.arenaria(Neal)],蔬菜一旦被侵染,植株生长缓慢、叶片发黄,植株萎蔫,一般减产10%~20%,严重时达到75%以上[3]。咖啡短体(根腐)线虫是三类检疫性线虫,属寄主根系皮层的迁徙性内寄生线虫,受害根系出现浅褐色病斑,伴随次生病原物,造成根系根腐。国内主要采用形态学特征即雌成虫会阴花纹进行植物病原线虫种类鉴定,国外多采用形态鉴定和分子生物学鉴定相结合的方法。病原线虫对不同的寄主植物具有一定的专化性,准确鉴定病原线虫种类对选育抗病作物品种和采用防治措施有重要指导意义。因此,本课题组对宁波地区蔬菜病原线虫发生情况进行调查并进行种类鉴定,以便针对性地进行防治,确保安全生产。
1 材料与方法
1.1 试验材料
在宁波地区选取镇海九龙湖、宁波梅墟、慈溪周巷、慈溪龙山、江北费市、宁海冠庄、北仑小港、余姚泗门、奉化江口等有设施蔬菜农业或蔬菜产量较大的农场,对各种蔬菜根系和周边土壤取样检测。
1.2 试验方法
1.2.1 病害分级标准
在各蔬菜基地采用对角线取样法,调查10点,每点取样2株,共20株,调查病情指数及发病率。
根结线虫危害的记载标准,病情指数采用切根百分数法[4]:
0级 无根瘤;
1级 根瘤总长度占总根长度的1%~24%;
2级 根瘤总长度占总根长度的25%~49%;
3级 根瘤总长度占总根长度的50%~74%;
4级 根瘤总长度占总根长度的75%~100%。

1.2.2 土壤病原线虫分离方法
土壤中线虫的分离采用以下两种方法。
改良漏斗法:取土样约250 g,用双层纱布包好,改良漏斗法分离线虫。25℃左右静置24 h后,接取线虫液约10 mL镜检。
淘洗过筛离心法[5]:称取土样100 g,倒入水盆中,加水搅匀,静置1 min。将水倒入一组网筛,即上层为60目,下层为400目,边倒边振荡分样筛,防止水充满下层的400目筛而从筛中溢出。然后,再在盆中加入水后混匀,静置1 min,倒入网筛中,如此重复3次。将400目的分样筛取下,用喷头把400目网筛中的线虫悬液中的泥浆冲洗干净,倒入烧杯中,静置。将静置烧杯中的上层水轻轻倒掉,只保留下层大约30 mL水、线虫和泥浆的混合物。将混合物轻轻摇匀,倒入离心管中,在天平上调平衡,把平衡后的离心管放入离心机中,第一次离心(离心机的转速2 000 r/min,离心时间为4 min)。倒掉第一次离心后离心管内的上层液,保留土层。在离心管中分别注入比重为1.18 g/L的蔗糖溶液约10 mL,在天平上调平衡后,摇匀,放入离心机中进行第二次离心(离心机的转速2 000 r/min,离心时间为1 min)。离心后,迅速取出离心管,把离心管内的上层液倒入500目筛中,用水把蔗糖液冲掉,以防线虫在蔗糖液中脱水变形。然后把线虫液转移到培养皿中,镜检。
1.2.3 植物病原线虫鉴定
形态学鉴定:用De Man公式法对会阴花纹、2龄幼虫、雄虫观察和测计,雌虫会阴花纹制作方法参见刘维志等[6]。
分子生物学鉴定:用单条2龄幼虫提取DNA[7]。将单条线虫放入dd H2O清洗,挑取单条放入200μL PCR管中(含8μL dd H2O和1μL 10×PCR Buffer(Mg2+free)),液氮中放置1 min,85℃加热2 min,向PCR管中加入1μL 1 mg/mL蛋白酶K,56℃加热15 min,95℃加热10 min,得到DNA提取液直接进行PCR扩增。
对mt DNA的COⅡ和l RNA基因间序列,采用1对引物进行扩增:上游引物为C2F3:GGTCAATGTTCAGAAATTTGTGG,下游引物为1108:TACCTTTGACCAATCACGCT[4]。
PCR扩增采用50μL反应体系:10×PCR Buffer(Mg2+free)5μL,25 mmol/L MgCl25μL,0.1 mmol/L d NTP 4μL,10μmol/L上游引物和下游引物各3μL,5 U/μL Taq酶0.6μL,DNA 提取液3μL,dd H2O补齐50μL。扩增程序:94℃4 min;94℃1 min,50℃ 1 min,72 ℃ 2 min,35个循环;最后72℃延伸10 min。
扩增产物在1%琼脂糖凝胶中电泳分离,如获得预期的PCR产物,则用Hin f I进行限制性酶切反应,酶切产物在2%琼脂糖凝胶中电泳分离,再用凝胶图像分析系统照相。
2 结果与分析
2.1 宁波病原线虫田间调查情况
通过2008年和2012年对宁波主要蔬菜基地的调查发现:江北费市、宁海冠庄、宁波梅墟和镇海九龙湖发现有南方根结线虫危害症状,其余基地未发现。镇海九龙湖和宁波梅墟两地的土壤中均分离到南方根结线虫的2龄幼虫;江北费市分离到咖啡短体线虫,除这两种病原线虫外,还分离到螺旋属(Helicotylenchus)、真滑刃属(Aphelenchus)、滑刃属(Aphelenchoides)、矮化属(Tylenchor hynchus)、盘旋属(Rotylenchus)、长尾属(Seinura)、垫刃属(Tylenchus)等其他非病原线虫。南方根结线虫的寄主有番茄、黄瓜、豇豆和白菜,咖啡短体线虫的寄主有萝卜和大豆。
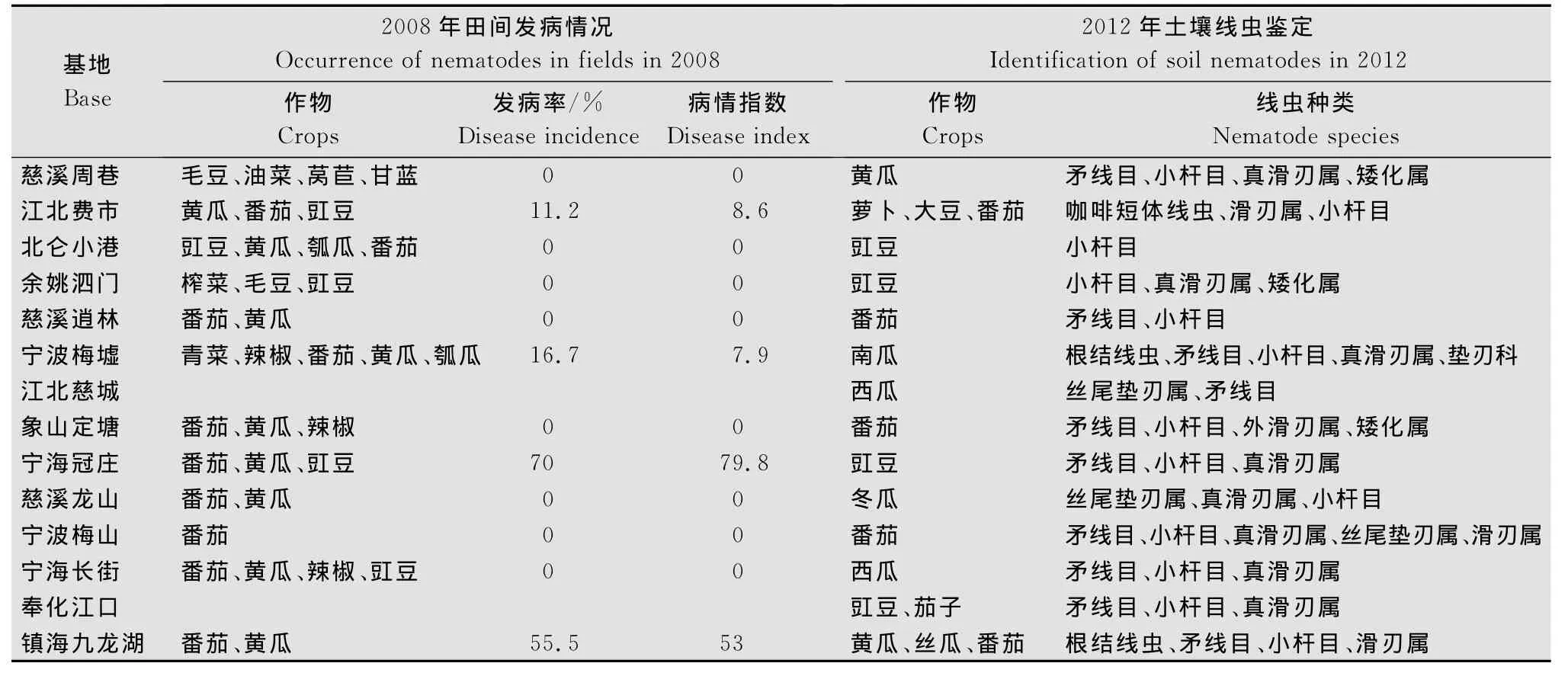
表1 宁波主要蔬菜基地植物病原线虫的发生情况Table 1 The occurrence of plant pathogenic nematodes on main vegetable bases in Ningbo
2.2 南方根结线虫的鉴定
根据获得的雌虫会阴花纹、雄虫和2龄幼虫的测计值,结合分子生物学分析,所有发现的根结线虫均为南方根结线虫。
2龄幼虫:体长390(360~470)μm,口针长11.0(10.1~11.8)μm,半月体在排泄孔前或与其相邻。尾较细,长47(43~50)μm,末端略钝圆,透明尾长6~14μm。背食道腺开口距口针基球1.3~2.5μm(图1)。
雌虫:虫体呈梨形,后部不隆起。口针长15~16μm,基部球圆。会阴花纹背弓相对较高,由平滑到波浪形的线纹组成,侧线不明显。有些会阴花纹发生变异,背弓变得圆而扁平。背食道腺开口距口针基球2~4μm(图1)。
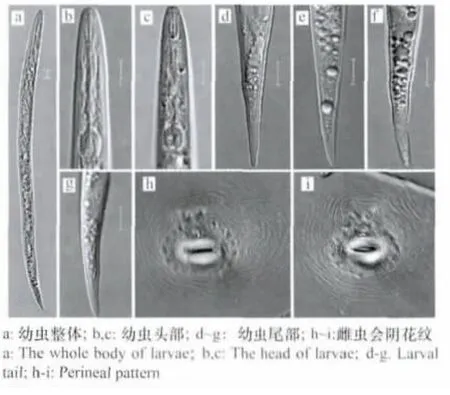
图1 南方根结线虫2龄幼虫和雌虫会阴花纹(标尺=10μm)Fig.1 Second instar larvae of Meloidogyne incognita and perineal patter n of female wor m(Scale=10μm)
雄虫:温热杀死后虫体向腹面略弯。体前端较细,后部较粗,尾部弯曲约90°,侧区有4条侧线。唇区无明显缢缩,头冠与头区分界明显,头冠比头区稍窄,头区有2~3条不明显的环纹,头架骨化明显。口针锥体前端细,后端逐渐加粗,口针杆部圆柱形,上下几乎等宽。口针基球为球形或扁球形。背食道腺开口距口针基球2.9(2.0~3.7)μm。
应用COⅡ和ⅠRNA基因间的C2F3和1108这对引物,扩增获得的PCR产物片段大小为1 700 bp,该产物经Hin f I限制性内切酶酶切,获得2条酶切产物,其片段大小为1 300 bp和400 bp(图2),这与文献报道一致[8-9]。
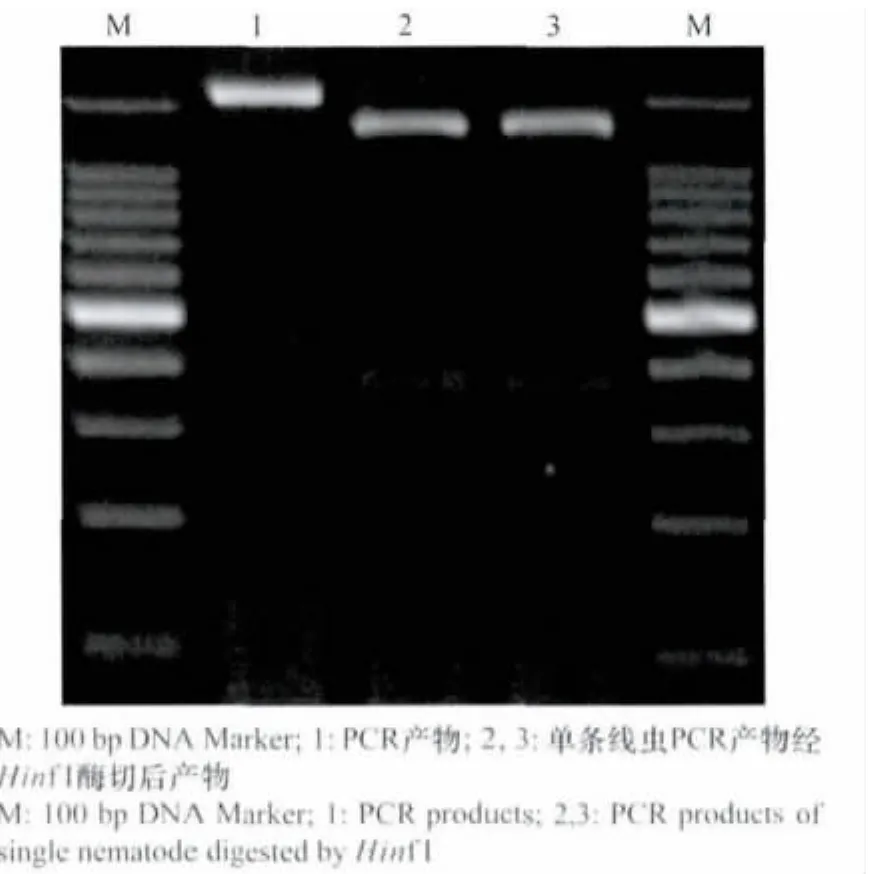
图2 C2F3和1108引物扩增产物的Hin f I酶切结果Fig.2 PCR products using pri mers C2F3 and 1108 digested by Hin f I enzy me
2.3 咖啡短体线虫的鉴定
在对宁波地区采集的土样调查时,还从费市的萝卜和大豆根围土壤中发现了一种短体属线虫,经形态学和分子生物学方法,鉴定为咖啡短体线虫。该线虫为植物根部内寄生线虫,对农作物危害较大,短体属非中国种,是我国进境植物检疫对象,该线虫未见在宁波发生的报道。
该线虫唇环2条,极个别的唇一侧为3条,唇区略缢缩;口针基球圆形;受精囊长卵形或近圆形,内充满精子;阴道较直;后阴子宫囊长17~50μm,有细胞分化;尾通常亚圆柱形,无纹,钝圆,平截或有凹痕,符合咖啡短体线虫主要鉴定特征(图3)。
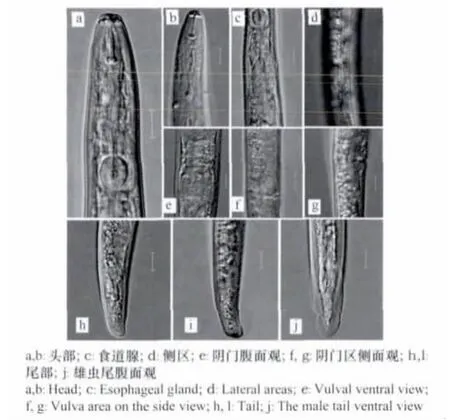
图3 咖啡短体线虫形态特征Fig.3 Mor phological characteristics of Pr atylenchus cof f eae
在获得单条线虫的DNA粗提液后,对核糖体28S基因的D2和D3区,采用1对引物进行扩增:上游引物为D2A:ACAAGTACCGTGAGGGAAAGTTG,下游引物为D3B:TCGGAAGGAACCAGCTACTA[10]。其他同上。PCR扩增产物送南京金思瑞公司直接测序,测序结果用 MEGA 4.0软件自带的Clustal W进行比对、拼接和人工校正,然后用邻接法构建系统发育树(图4),采自萝卜的序列号为KC786880;采自大豆的序列号为KC786881。
28S DNA序列进化关系分析表明,采自宁波大豆和萝卜上的线虫与日本和印尼的2种咖啡短体线虫聚类成一个分支,进一步确认该线虫为咖啡短体线虫。

图4 部分短体属线虫28S DNA序列构建的系统发育树Fig.4 Phylogenetic trees based on 28Sr RNA sequences of Pr atylenchus spp.
3 结论
赵雷等2007-2009年期间,对浙江省不同地区蔬菜上病原线虫样本进行形态学鉴定,鉴定出南方根结线虫和爪哇根结线虫。姚红燕等在2009年对宁波地区番茄上根结线虫进行形态学鉴定,确定为南方根结线虫[11],两者均未进行分子生物学鉴定。本研究通过形态学和分子生物学鉴定,确认宁波主要蔬菜基地存在南方根结线虫和咖啡短体线虫两种病原线虫,这对下一步开展抗病育种等针对性的防治措施具有重要价值。
对宁波主要蔬菜基地的调查发现,南方根结线虫和咖啡短体线虫的分布均比较集中,如咖啡短体线虫只出现在江北费市采样区,寄主为萝卜和大豆。南方根结线虫分布在江北费市、宁海冠庄、宁波梅墟和镇海九龙湖4个采样区,寄主主要有番茄、黄瓜、豇豆和白菜。南方根结线虫在不同寄主上的危害特性基本相同,以侧根和须根最易受害,根系受害后形成大小和形状不同的瘤状根结,根结之上一般可长出细弱新根,再度感染形成根结。地上部初期中午萎蔫,早晚恢复,严重者全株枯死;叶片小、叶色变浅、变黄、似缺素症;落花落果,果实小而畸形。咖啡短体线虫又称根腐线虫,主要危害根系,受害根系出现浅褐色病斑,伴随次生病原物,造成根系根腐。
根结线虫和咖啡短体线虫的防治目前均采用综合防治措施,比较有效的农业防治措施是水旱轮作和采用抗病品种,药剂防治方面目前生产中比较好的防治药剂是阿维菌素和淡紫拟青霉等生物制剂。
[1] 刘维志.植物病原线虫学[M].北京:中国农业出版社,2000.
[2] 杨宁,段玉玺,陈立杰,等.植物寄生线虫生物防治中存在的问题及解决途径[J].植物保护,2006,32(4):4-9.
[3] 赵鸿,彭德良,朱建兰.根结线虫的研究现状[J].植物保护,2003,29(6):6-9.
[4] 肖炎农,王明祖,付艳平,等.蔬菜根结线虫病情分级方法比较[J].华中农业大学学报,2000,19(4):336-338.
[5] 陈立杰,王旭,段玉玺,等.蔗糖离心法分离土壤线虫的最佳条件筛选[J].沈阳农业大学学报,2007,38(6):849-851.
[6] 刘维志.植物线虫学研究技术[M].沈阳:辽宁科学技术出版社,1995.
[7] 王江岭,张建成,顾建锋.单条线虫DNA提取方法[J].植物检疫,2011,25(2):32-35.
[8] Orui Y.Identification of Japanese species of the genus Meloidogyne (Nematoda:Meloidogynidae)by PCR-RFLP analysis[M].Applied Ento mology and Zoology,1998,33:43-51.
[9] Powers T O,Harris T S.A poly merase chain reaction method for identification of five major Meloidogyne species[J].Jour nal of Nematology,1993,25:1-6.
[10]De Ley P,Felix M A,Frisse L M,et al.Molecular and morphological characterisation of t wo reproductively isolated species with mirror-i mage anatomy (Nematoda:Cephalobidae)[J].Nematology,1999,1(6):591-612.
[11]姚红燕,周金波,谌江华,等.宁波地区番茄根结线虫的发生及防治[J].浙江农业科学,2009(2):375-376.
猜你喜欢
今日农业(2022年14期)2022-09-15
当代水产(2021年8期)2021-11-04
昆明医科大学学报(2021年4期)2021-07-23
科技视界(2020年26期)2020-09-24
科技视界(2020年17期)2020-07-30
中国森林病虫(2019年5期)2019-10-11
猪业科学(2018年4期)2018-05-19
中成药(2018年2期)2018-05-09
化学教学(2018年1期)2018-02-28
浙江大学学报(农业与生命科学版)(2017年1期)2017-04-17
