
L-型钙电流在血管紧张素Ⅱ诱导新生大鼠肥大心肌细胞中的功能和分子水平改变
2012-11-09 06:58闫秋丽
湖北科技学院学报(医学版) 2012年5期
闫秋丽
(武汉大学基础医学院,湖北武汉430074)
在压力负荷和神经体液刺激作用下,心肌细胞产生适应性肥大生长,以暂时性改善心脏功能。但这种心脏适应性肥厚能够使心血管发病率和死亡率增高,这主要与其引起的电生理重塑和心律失常有关[1]。心脏的电生理重塑通常表现为复极化过程延迟而致动作电位时程(action potential duration,APD)延长等,其机制涉及到多种离子通道的改变和内外向电流的平衡,包括L型Ca2+电流这种内向离子流的变化。
在压力负荷导致的心肌肥厚中有多种神经体液因素参与,其中血管紧张素Ⅱ(AngiotensinⅡ,AngⅡ)是重要的体液因子。研究发现,血管紧张素Ⅱ能调节心肌细胞中多种离子电流或通道的功能,增强成年动物心肌细胞的L-型钙电流(L-type Ca2+current,ICa,L)[2]。而在人和鸡胚胎心肌细胞上,AngⅡ却是增强胚胎心肌的T-型钙电流,而减弱 L-型钙电流[3]。因此,AngⅡ对胚胎心肌 L-型钙电流的作用似乎与对成熟心肌的作用相反。而电压门控性钾电流在很大程度上决定了心肌细胞动作电位时程。钾通道的功能失调可导致室性心律失常,如:尖端扭转型室性心动过速(torsades de pointes)和长 Q-T 综合症引起的室性纤颤[4]。有报道称,AngⅡ也可抑制心肌细胞缓慢激活的延迟整 流 性 钾 电 流 (slowly-activating delayed-rectifier K+current,Ik,s),而增强快速激活的延迟整流性钾电流(rapidly-activating delayed-rectifier K+current,Ik,r)[5,6]。Ang Ⅱ调节各种跨膜离子流作用的存在,提示AngⅡ也可能促进了心肌电生理重构过程。事实上,AngⅡ被证实可增强培养心肌细胞缝隙连接蛋白connexin43的表达[7]。这种缝隙连接蛋白在心肌细胞间动作电位传导中起着关键作用。AngⅡ还可抑制心肌细胞Kv4.3通道蛋白的表达,减少心肌钾电流Ito;抑制心肌细胞Na+通道蛋白的表达,减少心肌Na+电流密度[8]。
为了探索AngⅡ诱使心肌细胞肥大时,心肌细胞L-型钙电流的功能特征和分子表达水平,本实验拟在培养的新生大鼠心肌细胞上,观察AngⅡ所诱导肥大心肌细胞中L型Ca2+通道的功能特征和分子表达水平的变化,以便为阐明病理情况下心肌电生理重构机制提供实验依据。
1 材料与方法
1.1 实验仪器
MilⅡ-Q纯水机:MilliPore公司制,普通4℃与-20℃冰箱。YS-1300-V 超净工作台。DGG-9249电热恒温干燥箱:压力蒸汽灭菌器:CO2恒温培养箱:日本SANYO公司。精密天平(Sartorius BSⅡ110S)。BeideSH-5型磁力搅拌器,pH测定计,电热恒温水浴箱,Olympus IX71倒置显微镜:日本OLYMPUS公司。膜片钳放大器:型号 Axopatch 700B,美国Axon公司。微电极拉制仪:Narishige P-97,日本。三维操纵器:Narishige,日本,紫外分光光度仪,低温高速台式离心机,PCR扩增仪,电泳槽和电泳仪,1008-0635(2012)05移液器等。
1.2 药品与试剂
DMEM培养基:Sigma公司。胎牛血清:GIBCO公司。胰蛋白酶:GIBCO公司。D-Hanks液:鼎国自产。溴脱氧尿苷(BRDU):Sigma公司。多聚赖氨酸:Sigma公司。RNeasy Mini Kit:德国Qiagen公司。RT-PCR试剂盒:日本 Toyobo公司。DNA Marker:美国Promega公司产品。
1.3 实验动物
将健康体质量250~300g的SD大鼠雌、雄各一只,合笼喂养。待雌鼠怀孕后,将其从笼中取出,待产。分娩后新生1~2d的健康SD大鼠仔鼠,用于实验。
1.4 实验方法
1.4.1 新生大鼠心肌细胞分离、培养
取出生后1~2 d的SD新生大鼠,无菌条件下开胸剪取心室肌部分,D-Hanks液清洗去除残血,将其剪碎,0.1%胰蛋白酶37℃反复消化直至获得足量的细胞。每次消化5min,弃去第一次消化的上清,收集以后消化的上清,转移到含有血清的DMEM培养基中以终止胰酶作用。300×g离心7min,弃上清取细胞沉淀加入含15%新生牛血清(NBS)的DMEM培养基制备成细胞悬液,在37℃,5%CO2培养箱中培养90 min,以差速贴壁法纯化心肌细胞,以2×105/瓶的细胞密度接种至25ml培养瓶继续培养72h开始实验。培养的前72 h加入0.1mmol/L 5'-溴脱氧尿苷以抑制非心肌细胞增殖。
1.4.2 心肌细胞肥大的诱导
心肌细胞接种72h后长成单层,可观察到同步搏动。换液时在培养基中加入AngⅡ至终浓度10-7M,之后每24h换液并加药一次,连续作用3d。设空白对照组细胞作相同换液处理,但不加AngⅡ。
1.4.3 心肌细胞总蛋白含量测定
心肌细胞肥大时,细胞内总蛋白含量会升高。因此,心肌细胞内总蛋白含量检测时常被作为鉴定心肌细胞是否肥大的指标之一。本实验参照南京建成生物工程公司考马斯亮蓝蛋白检测试剂盒说明测定心肌细胞蛋白质含量,根据各瓶细胞数,计算出每瓶细胞的总蛋白含量。蛋白含量(mg/ml)=(测定管 OD值-空白管OD值)/标准管OD值-空白管OD值/标准管浓度/稀释倍数。
1.4.4 电生理研究
细胞置于倒置显微镜上的恒温灌流槽中,温度控制在 22℃ ~24℃,先灌流标准台氏液(mmol/L):NaCl 140,NaOH 2.3,KCl 5.4,CaCl21.8,MgCl21,HEPES 10,Glucose 10,pH 7.4。记录L-Ca2+电流时细胞外液含(mmol/L):Choline-Cl 136,CsCl 5.6,CaCl22.0,MgCl21.0,NaH2PO40.33,HEPES 5,和葡萄糖10,用CsOH调节pH至7.4。记录L-Ca2+电流时电极内液成分(mmol/L):CsCl 20,Cs aspartate 110,HEPES 10,EGTA 10,MgCl21.0,Na2ATP 5,GTP 0.1,和磷酸肌酸 5,用CsOH调节pH至7.4。
记录电极由自动拉制仪制备,充灌电极内液后的电极电阻为1~3 MΩ。用电动显微操纵器将电极推向细胞,负压吸引形成高阻封接,负压吸破细胞膜,补偿电容电流和串联电阻(80% ~85%)以减少瞬时充放电电流和钳位误差,形成全细胞记录模式。信号经 Ag-AgCl电极引导,由 Axon 700B膜片钳放大器(Axon Instrument,USA)放大,经模/数转换系统(Digidata 1322A)采集数据,并进行数据分析。信号经截止频率为2 kHz的四阶贝塞尔低通滤波器,采样率为10 kHz,采样后数据存于计算机硬盘内,供测量及分析用。于细胞破膜后5 min开始记录ICa,L,使电极内液和细胞内液充分交换,减少了系统误差。为消除细胞间误差,电流值以电流密度(pA/pF)表示。以钳制电压为-50 mV,超级化-10 mV得到的细胞电容电流积分值,计算出细胞膜电容。细胞膜电容大小能反映细胞尺寸。
1.4.5 半定量RT-PCR
采用反转录-多聚酶链扩增反应法检测种L-型Ca2+通道(α1C亚单位)的mRNA表达量。
(1)总RNA的提取:用Trizol试剂一步法,提取总RNA的、接触组织的器具均经过0.5%DEPC水处理。①将培养皿中的培养液彻底弃干净。将1 ml Trizol直接加在培养皿中的细胞上。将细胞并Trizol一起移入Eppendorf离心管中,匀浆。②室温静置5 min,再加0.25 ml氯仿摇匀。静置5 min后,离心15 min(12000 g)。③水相移至新管,加0.7倍体积异丙醇,混匀。静置10 min后,离心10 min(12000 g),弃上清。④加75%乙醇1 ml(DEPC水新鲜配制),混匀,再离心5 min(7500 g)。⑤蒸馏水溶解RNA。⑥分光光度计测A260/A28O比值为1.7~2.0。⑦1%琼脂糖凝胶电泳观察185与285条带密度比值约等于2,确保RNA的纯度和完整性。
(2)RT(逆转录):取逆转录cDNA产物2 μl,加L-型Ca2+通道α1C亚单位上下游引物(序列:Forward:5'-CCTCAGTGCAGACACATTTGC-3',Reverse:5'-CCGTTACAGCAG CACCCCCACA-3'。由上海生物工程公司合成。扩增产物尺寸为253 bp)各2 μl及内参 GAPDH上下游引物(序列:Forward:5'-ACGGCAAATTCAACGGCACAGTCA-3',Reverse:5'-CTGGGGGCATCGGCAGAAGG-3'。由上海生物工程公司合成。扩增产物尺寸为251 bp)各 2 μl,2 × PCR Master Mix(Fermentas)12.5 μl,灭菌水 2.5 μl,总体积为 20 μl。扩增反应条件为:94 ℃,30 s,58 ℃,40 s;72℃,1 min;30 个循环;完成全部循环后72℃ 10 min以达到充分延伸。
取RT-PCR产物6 μl进行2%琼脂糖凝胶平板电泳,将电泳条带图像摄入凝胶成像分析系统,保存为tiff格式图片,供分析。
1.5 数据分析
采用Origin 7.5 software(Microcal Software Inc.,USA)和 Clampfit 10.0 software(Axon Instruments Inc.,USA)软件分析电生理数据。电流强度的测定,以电流激活过程中刚出现时数值与电流达最大激活时数值之间的差值为准。电流密度大小即为电流强度与膜电容的比值(pA/pF)。
所有数据均以mean±S.E.M.表示。采用t检验和方差分析。P<0.05被认为具有统计学差异。
2 结果
2.1 各实验组心肌细胞总蛋白含量和膜电容值的比较
从表1可以看出,与正常对照组相比,AngⅡ组心肌细胞的总蛋白含量明显增高(P<0.01);应用AngⅡ1型受体阻断剂Losartan明显使心肌细胞的总蛋白含量恢复至正常水平。
与心肌细胞总蛋白含量改变一致,AngⅡ组心肌细胞也要高于正常对照组(P<0.01);Losartan同样能使心肌细胞的膜电容值恢复正常。膜电容值的升高反映了心肌细胞尺寸的扩大。
这些结果表明,AngⅡ诱导心肌细胞肥大厚模型建立成功了。

表1 各实验组心肌细胞总蛋白含量和膜电容值
2.2 AngⅡ诱导的肥大心肌细胞中L-型Ca2+电流密度的改变
在形成全细胞封接状态后,将细胞钳制在-50 mV,给予指令电压从-40 mV~+80 mV,波宽为300 ms的刺激,可记录到一系列的ICa,L。当加入硝苯地平(3 μmol/L),再给予上述刺激后,可见该电流被迅速抑制,说明该电流为 L-型Ca2+电流。图1是实验中记录的典型L-型Ca2+电流曲线。实验连续记录 ICa,L15 min,未见明显的“rundowm”现象。
如图2A所见,AngⅡ组心肌细胞的 L-型Ca2+电流密度在-20~+40 mV范围内明显高于正常组。其中,0 mV处 AngⅡ组 L-型Ca2+电流密度(6.13 pA/pF)比正常组(4.03 pA/pF)增高52.2%(P<0.01)。但L-型Ca2+电流激活电位和翻转电位并没有改变。Losartan可以使大鼠心肌细胞L-型Ca2+电流密度恢复至正常组水平。
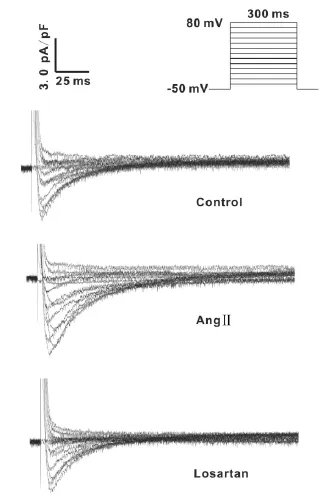
图1 记录的典型L-型Ca2+电流曲线
2.3 AngⅡ诱导的肥大心肌细胞中L-型Ca2+通道动力学参数变化
稳态激活特征:记录ICa,L稳态激活曲线时刺激方案与记录I-V曲线的方案相同。稳态激活曲线是将I-V曲线转换成膜电导后得到,用Boltzmann 方程:G/Gmax=1/{1+exp[(V-Va)/k]}进行拟合。其中,G是膜电压为 V时的膜电导,Gmax为最大膜电导,Va为半数激活电压,k为斜率因子,G=I/(V-Erev),I为对应膜电压V时的峰电流,Erev为通过I-V曲线测得的反转电位。
如同图2B所显示,正常组、AngⅡ组、Losartan组三组的ICa,L激活曲线特征没有明显不同。三组的半数激活电压Va分别为(-7.86±1.03)mV、(-8.15±1.21)mV、(-8.06±1.18)mV,P>0.05。
稳态失活特征:记录ICa,L稳态失活曲线应用双脉冲刺激,钳制电压为 -50 mV,条件脉冲从-60 mV开始,阶跃10 mV,时间3000 ms;随后测试脉冲为10 mV,时间300 ms。稳态失活曲线是用 Boltzmann 方程:I/Imax=1/{1+exp[(V-Vi)/k]}进行拟合。其中,I为测试脉冲引出的峰电流,Imax为测试脉冲引出的最大峰电流,V为条件脉冲电压,Vi为半数失活电压,k为斜率因子。
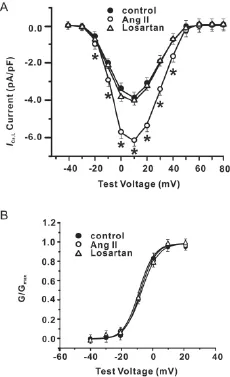
图2 三组细胞L-型Ca2+电流的I-V曲线和激活曲线,*P<0.01 vs control
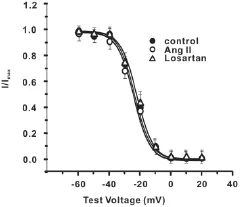
图3 三组心肌细胞L-型Ca2+电流的稳态失活曲线

图4 三组心肌细胞L-型Ca2+电流的复活曲线

图5 三组心肌细胞L-型Ca2+通道α1C亚单位mRNA的表达水平。
结果显示,正常组、AngⅡ组、Losartan组三组的失活曲线特征相近(图3)。三组的半数失活电压Vi分别为(-24.86±3.65)mV、(-25.55±4.01)mV、( -24.31±3.87)mV,P >0.05。
失活后复活特征:记录ICa,L稳态失活后恢复曲线采用双脉冲刺激,钳制电压为 -50 mV,给予+10 mV、时间为200 ms(P1)和200 ms(P2)的两个方波刺激,两刺激间隔时间为从50 ms开始,每次递增50 ms,直至750 ms。分别记录P1和P2对应的峰电流。计算复活电流指数(P2/P1)。以复活电流指数对两刺激间隔时程作复活曲线。并用单指数方程:P2/P1=A1 exp(-t/τ)+A0 进行拟合。求出复活时间常数τ。结果显示,正常组、AngⅡ组、Losartan组三组的失活后恢复曲线特征相近(图4)。三组的复活时间常数分别为(115±13)ms、(120 ±25)ms、(118 ±24)ms,P >0.05。
2.4 AngⅡ诱导的肥大心肌细胞中L-型Ca2+通道α1C亚单位mRNA表达的改变
为了确认AngⅡ诱导的肥大心肌细胞中L-型Ca2+通道分子水平上的改变,我们采用RT-PCR检测了心肌细胞中L-型Ca2+通道α1C亚单位的mRNA表达量。图3A是RT-PCR产物的电泳条带。如图5A所显示,AngⅡ组心肌细胞中α1C亚单位的mRNA表达量明显高于正常组。为了进一步的实时定量检测α1C亚单位的mRNA表达量,我们又做了Real-time PCR(实时定量 PCR),定量检测了α1C亚单位的表达量。实时定量PCR检测结果进一步证实,AngⅡ组心肌细胞中α1C亚单位的mRNA表达明显增高,高于正常组56.0%(图5B)。
3 讨论
3.1 AngⅡ诱导的肥大心肌细胞中L-型Ca2+通道的分子和功能水平改变
尽管目前已有一些实验证实,AngⅡ可以调节心肌细胞 ICa,L电流[2,3]。但是,这些实验并没有探讨AngⅡ对L-型Ca2+通道表达的影响。当前研究的主要新发现是,AngⅡ在引起新生大鼠心肌肥大的同时,增加心肌细胞 ICa,L电流和 L-型Ca2+通道α1C亚单位mRNA表达。
L-型Ca2+通道主要是由α亚单位和β亚单位构成[10]。单独α亚单位表达即可形成功能性L-型Ca2+通道孔,是主要功能亚单位;β亚单位是调节性亚单位,它与α亚单位共表达,可以调节L-型Ca2+通道的功能特征。目前研究发现,在心室肌等工作细胞中表达的主要是α1C亚单位。在窦房结、房室结中还有少量α1D亚单位的表达。心脏组织中并无其他α亚单位类型。心脏的β亚单位主要是β2亚单位。基于上述发现,本实验只检测了培养新生鼠心室肌细胞的L-型Ca2+通道α1C亚单位的表达。
本实验研究中,AngⅡ对新生大鼠心肌细胞ICa,L电流的影响是与其调节L-型Ca2+通道 α1C 亚单位表达的作用相互一致。AngⅡ所诱导的ICa,L电流密度增加可能与其促进L-型Ca2+通道α1C亚单位表达增加有关。另外,本研究也发现,AngⅡ虽然增加了心肌细胞ICa,L电流密度,但对L-型Ca2+通道的激活、失活、复活等通道门控动力学特征并无影响。因而,AngⅡ可能只是增加α1C亚单位的表达量,而对心肌细胞L-型Ca2+通道的其它亚单位构成类型并没有影响。
在心肌细胞中,AngⅡ的作用受体至少有两种,即血管紧张素Ⅱ1型受体(angiotensin receptor type 1,AT1R)和血管紧张素Ⅱ2型受体(angiotensin receptor type 2,AT2R)[11]。大部分 Ang Ⅱ的作用是由AT1R介导的[12]。同样,本实验研究发现,AngⅡ增加新生鼠心室肌细胞ICa,L电流密度和L-型Ca2+通道α1C亚单位表达的作用也是由AT1R介导的。应用AT1R拮抗剂losartan能抑制的这些作用。
3.2 AngⅡ增加 ICa,L电流密度和 L-型 Ca2+通道表达的可能意义
Ang Ⅱ对 ICa,L电流密度和 L-型Ca2+通道表达的增强,可能增加心肌细胞动作电位时期跨细胞膜内流的 Ca2+量[13],而增加细胞内 Ca2+负荷。这种Ca2+负荷加重(或称Ca2+超载)可以导致触发活动发生,引起心律失常[14]。这也许是血管紧张素转换酶抑制剂(angiotensin converting enzyme inhibitor,ACEI)和血管紧张素受体拮抗剂(angiotensin receptor antagonist,ARB)能够对室性心律失常具有一定防治作用的原因[15,16]。
[1]Koren MJ,Devereux RB,Casale PN,et al.Relation of left ventricular mass and geometry to morbidity and mortality in uncomplicated essential hypertension[J].Ann Intern Med,1991,1(14):345
[2]Vila Petroff MG,Aiello EA,Palomeque J,et al.Subcellular mechanisms of the positive inotropic effect of angiotensin Ⅱ in cat myocardium[J].Journal of Physiology,2000,529(1):189
[3]Bkaily G,Sculptoreanu A,Wang SM,et al.AngiotensinⅡ-induced increase of T-type Ca2+current and decrease of L-type Ca2+current in heart cells[J].Peptides,2005,26:1410
[4]Tristani-Firouzi M,Chen J,Mitcheson JS,et al.Molecular biology of K+channels and their role in cardiac arrhythmias[J].Am J Med,2001,110:50
[5]Daleau P,Turgeon J.Angiotensin Ⅱ modulates the delayed rectifier potassium current of guinea pig ventricular myocytes[J].Pflugers Arch,1994,427:553
[6]Yang XJ,Jiang WP.Angiotensin Ⅱ receptor regulates ionic currents in guinea pig ventricular myocytes[J].J Cardiovasc Pharmacol Ther,1998,3:1
[7]Dodge SM,Beardslee MA,BJ Darrow,et al.Effects of angiotensinⅡon expression of the gap junction channel protein connexin43 in neonatal rat ventricular myocytes[J].J Am Coll Cardiol,1998,32:800
[8]Shang LL,Sanyal S,Pfahnl AE,et al.NF-kappaB-dependent transcriptional regulation of the cardiac scn5a sodium channel by angiotensin Ⅱ[J].Am J Physiol Cell Physiol,2008;294(1):C372
[9]Livak IG,Schmittgen TD.Analysis of relative gene expression data using real-time quantitative PCR and the 2[-delta delta C(T)]method[J].Methods,2001,25(4):402
[10]张建成,黄从新,邓玉莲.L-型钙通道与心房快速激动诱发的心房电重构[J].中国心脏起搏与心电生理杂志,2003,17(1):68
[11]Suzuki J,Matsubara H,Urakami M,et al.Rat angiotensinⅡ(type 1A)receptor mRNA regulation and subtype expression in the myocardial growth and hypertrophy[J].Circ Res,1993,73:439
[12]Baker KM,Booz GW,Dostal DE.Cardiac actions of angiotensin Ⅱ:role of an intracardiac renin-angiotensin system[J].Annu Rev Physiol,1992,54:227
[13]Kass RS,Tsien RW.Fluctuations in membrane current driven by intracellular calcium in cardiac Purkinje fibers[J].Biophys J,1982,38:259
[14]Song LS,Guia A,Muth JN,et al.Ca2+signaling in cardiac myocytes overexpressing the alpha(1)subunit of L-type Ca2+channel[J].Circ Res,2002,90:174
[15]Anvari A,Türel Z,Schmidt A,et al.Angiotensin-converting enzyme and angiotensinⅡreceptor 1 polymorphism in coronary disease and malignant ventricular arrhythmias[J].Cardiovasc Res,1999,43:879
[16]Hiura N,Wakatuki T,Yamamoto T,et al.Effects of angiotensinⅡtype I receptor antagonist(candesartan)in preventing fatal ventricular arrhythmias in dogs during acute myocardial ischemia and reperfusion[J].J Cardiovasc Pharmacol,2001,38:729
猜你喜欢
世界科学技术-中医药现代化(2022年2期)2022-05-25
世界科学技术-中医药现代化(2021年7期)2021-11-04
电镀与环保(2016年3期)2017-01-20
电镀与环保(2016年3期)2017-01-20
电镀与环保(2016年2期)2017-01-20
海南医学(2016年8期)2016-06-08
中国病理生理杂志(2015年8期)2015-12-21
河南科技(2015年2期)2015-02-27
电镀与环保(2014年3期)2014-09-18
微创医学(2012年6期)2012-01-13
