
白垩纪−古近纪古生物地理区的变化与印度和亚洲的碰撞
2012-09-20 00:46:46-Q,-B
地球学报 2012年1期
白垩纪−古近纪古生物地理区的变化与印度和亚洲的碰撞
E-mail: wanxq@cugb.edu.cn.
Geologic studies of the Late Cretaceous-Paleogene stratigraphic record of southern Tibet reveal distinct episodes of collision and post-collisional compression. The composition of sediments exposed along the Indus-Yarlung Zangbo suture, tectonic deformation, paleomagnetism, magmatic activity of the Gangdese Arc, high pressure metamorphism, paleobiogeography and terrestrial biota changes have been variously used to diagnose the timing of Indian-Eurasian continental plates collision.Among them, paleobiogeography and marine facies cessation are visible scenarios to trace the continental collision. This study recognized that dramatic changes of paleobiogeography happened during early Paleogene.
It is firmly established that the rise of the Tibetan plateau is the result of convergence of different terranes. During the middle Cretaceous, the Lhasa block had attached to the Eurasian continent. The Bangong Lake–Nujiang River suture closed at the end of Jurassic. By that time the Lhasa block was at the south margin of the Eurasian continent, thus the shallow water faunas like Orbitolinids and bivalves could migrate along the east and west margin of the Eurasian continent. Paleonbiogeographically, the Lhasa Block belongs to the north temperate realm. The Indian Plate started to move away from Africa by early Cretaceous and drifted to the north. It was isolated from Eurasian continent to the north and from Africa to the west by Neo-Tethys Ocean that was a barrier for migration of shallow water faunas. No Orbitolinids have been found from Indian Plate where faunas are shown not to be closely related to contemporary shallow water biota of the Eurasian continent.
In the late Cretaceous, the distribution of Orbitolinids in Tibet is limited in the Lhasa Block and some parts of the Qiangtang Basin. Its distribution extends to Myanmar in the east and Ladakh to the west. Along the coast of Tethys, it migrated to Japan, Malaysia,Indonesia, Kashmir, Afghanistan, Mid-East, West and South Europe, North Africa and Caribbean regions. It never migrated into the Indian Plate where a mixed planktonic and smaller benthic foraminiferal fauna appeared during that time. Orbitolinids was found in the Tanganyika region in east Africa, but it did not migrate eastward into the Indian Plate either. Like Orbitolinids, a bivalve fauna ofIsodomella–Caestocorbulafound in the Lhasa Block belongs to the Japan–East Asia sub-province, and no affinity has been found from the Indian continent. Whatley and Bajpai(2000) described a Masstrichtian ostracod fauna ofLimnocythere falsocarinata-Frambocythere tumiensis-Gomphocythere gomphiomatos. The fauna is shown to be an endemic Indian fauna without links to the specific level with China. The bio-province differences still existed in latest Cretaceous, when the benthic larger foraminiferal faunas are distinct at both sides of the Yarlung Tsangpo suture. TheOrbitoidesfauna is dominated in north margin of the Indian Plate, andLepidorbitoidesfauna in the Lhasa Block.
Toward Paleogene, faunas at both sides of the Yarlung Tsangpo suture tend to be identical. The PaleoceneRotalia-Lockhatia-Smoutina, late PaleoceneMiscellanea- Daviesinaand EoceneNummulites-Discocyclinalarger foraminiferal faunas occurred at both sides showing affinity of one biotic province. The Paleocene charophyte assemblage from the Deccan area of India also shows an affinity to that of South China.
Paleobiogeographic data from southern Tibet indicates that the initial contact of the Indian and Eurasian continental margins was in the early Paleocene,which allows the migration of shallow water faunas.However, the biotic migration and affinity provide only a minimum age to collision since marine conditions may persist on continental crust for some time after collision. Wan (1987) reported that the cessation of marine deposition in the north margin of Indian Plate was Late Eocene (early Priabonian), which is about 25 Ma after the initial contact as defined above.More uppermost marine fossil records were reported,such as the Paleocene dinoflagellates (Li et al, 2005)and radiolarian (Liu et al, 2002) in the Gyangze area,the Paleocene planktonic foraminifera from Zanda(Wan et al., 2005) and the early Eocene radiolaria in the Saga area (Li et al., 2007) among others.The gradual cessation of marine sedimentation in these residual basins is the result of continuous plate collision, but does not identify the initial stage of the plate contact. Our data indicate that India contacted Asia in early Paleocene (65-60Ma) and the collision did not finish until late Eocene (early Priabonian,~35Ma).
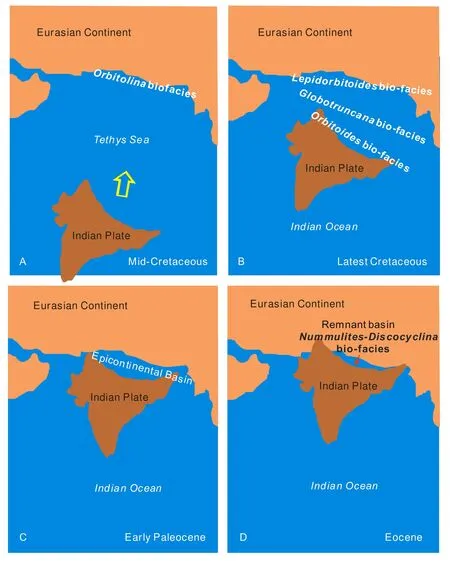
Fig. 1 Sketch map showing the paleobiogeographical change and India-Asian collision.A-Indian plate drifted north rapidly. Orbitolina fauna in mid-Cretaceous did not migrate over the Tethys sea south to India; B-Indian plate was near to Eurasian continent. A semi-pelagic sea still existed between two continents, it was the barrier for the migration of shallow water faunas; C-Initial contact of the Indian and Eurasian continental margins. An epicontonental basin occurred between them. Faunas became identical since Paleocene; D-Only remnant seas yielding same fauna remained between two continents. Marine environment ceased after late Eocene
LI G, WAN X, LIU W, LIANG D, HYESU Y. 2005. The discovery of Paleogene marine stratum along the southern side of Yarlung-Zangbo suture zone and its implications in tectonics.Sicences in China Ser.D Earth Sciences, 48(5): 647-661.
LI Y, WANG C, HU X, BAK M, WANG J, CHEN L. 2007. Characteristics of early Eocene radiolarian assemblages of the Saga area, southern Tibet and their constraint on the closure history of the Tethys. Chinese Science Bullutin, 52(15): 2108-2114.
LIU J, JONATHAN C. 2002. Upper Paleocene radiolarians from the Yamdrok mélange, south Xizang (Tibet), China. Micropaleontology, 48 (suppl.1): 145-154.
WAN X, LI J, ZHANG S, WU L. 2005. Late Cretaceous-Paleocene planktonic foraminifera from Zanda, western Tibet and their chronostratigraphic implication. Acta Micropaleontologica Sinica, 22(1): 10-18.
WHATLEY R C, BAJPAI S. 2000. A new fauna of Late Cretaceous non-marine ostracoda from the Deccan Intertrappean Beds of Lakshmipur, Kachchh (Kutch) District,Gujarat, Western India.Riv. Esp. de Micropleont., 32(2): 385-409.
Paleobiogeographical Change and India-Asian Collision during Cretaceous-Paleogene Period
WAN Xiao-qiao, LI Guo-biao, JIANG Tian
State Key Laboratory of Biogeology and Environmental Geology, China University of Geosciences, Beijing100083,China
India-Asian collision; Orbitolinids; Bivalve; Paleobiogeography
10.3975/cagsb.2012.s1.31
book=0,ebook=175
猜你喜欢
中国自动识别技术(2023年6期)2024-01-12 08:13:24
厦门科技(2021年4期)2021-11-05 06:50:44
小哥白尼(神奇星球)(2021年3期)2021-07-22 03:18:08
故事作文·高年级(2021年2期)2021-02-04 07:31:26
科学大众(2020年12期)2020-08-13 03:22:40
红领巾·探索(2018年12期)2018-01-26 12:34:14
足球周刊(2016年14期)2016-11-02 11:42:02
科学启蒙(2016年4期)2016-03-23 12:00:49
Coco薇(2015年12期)2015-12-10 03:11:51
生物进化(2014年3期)2014-04-16 04:36:43
