
珍稀濒危树种银缕梅种子萌发特性研究
2012-07-30 10:01胡国伟卢毅军李贺鹏俞立鹏金水虎
浙江林业科技 2012年6期
胡国伟,卢毅军,李贺鹏,俞立鹏,金水虎*
(1. 浙江农林大学 林业与生物技术学院,浙江 临安 311300;2. 杭州植物园,浙江 杭州 310007;3. 浙江省林业科学研究院,浙江 杭州 310023;4. 浙江省安吉县龙王山自然保护区管理处,浙江 安吉 313301)
银缕梅(Parrotia subaequalis)为金缕梅科(Hamamelidaceae)银缕梅属落叶小乔木,是中国特有的孑遗植物,具有极高的科研和经济价值。其木材具有细密、坚硬、比重大等优良性质,可供珍贵机械部件优良用材,是一种有待开发的特种木材资源。另外,银缕梅树冠开张,主干旁逸斜出,花形奇特,可作为优良行道树种;其叶片在秋季变成红、黄等色,可作为珍贵的秋季观叶树种;此外,银缕梅生长极其缓慢,树姿苍老,是制作中小型盆景的理想材料。1935年9月沈隽教授在江苏宜兴芙蓉寺首次采集到该种植物果枝标本[1]。自20世纪90年代至今,在浙江省安吉、临安,江苏省宜兴,安徽省舒城、金寨、绩溪才相继被发现,但数量极其稀少。在1999年8月国务院批准的《国家重点保护野生植物名录(第一批)》中,银缕梅被列为一级保护植物,并被国际自然保护联盟(IUCN)列为极度濒危(Critically Endangered,CR)物种[2]。
银缕梅自然种群分布极为狭窄,呈间断岛屿状散布于天目山北段及大别山东南部,且种群内个体数量稀少。目前有关银缕梅的研究主要集中在系统发育地位、形态结构特征、种群分布、光合生理生态特性、濒危机理等方面[3~8],对银缕梅种子萌发特性研究较少。本文通过研究储藏方法、温度、赤霉素浓度、光照条件等不同处理方式对银缕梅种子萌发的影响,探讨提高银缕梅种子发芽率的途径,为其有效保护和开发利用提供科学的理论依据。
1 材料与方法
1.1 材料
2010 年 10 月,在浙江省安吉县龙王山(30° 23′ 54.7″ N, 119° 24′ 39.5″ E)采得种子。种子风干后,分别采用常规储藏、3 ~ 5℃冷藏和常温沙藏保存。
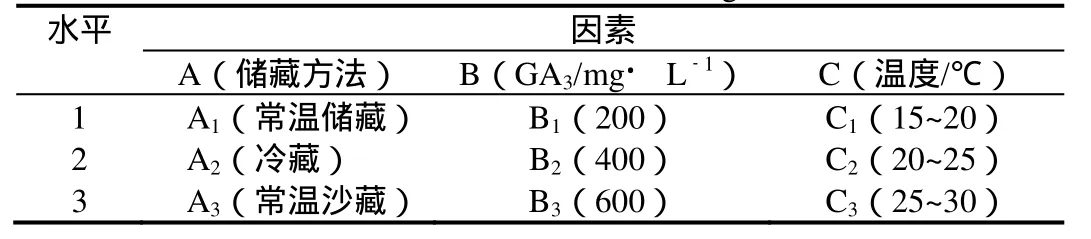
表1 正交实验设计的因素与水平Table 1 Factors and levels for orhogonal test
1.2 研究方法
1.2.1 种子形态学测定 种子大小测定:取10粒饱满种子,用游标卡尺测量种子的长与宽,重复5次,计算平均值。种子千粒重测定:随机取100粒饱满种子,用电子天平称量,重复5次,计算千粒质量。
1.2.2 种子萌发实验 将不同储藏方式的供试种子采用L9(34)的正交实验。因素与水平见表1。该实验共设9个处理,每一处理3次重复。每个处理随机选 50粒种子,置于细沙基质的培养皿内,在每天光照14 h,湿度75%的相应温度设置的培养箱内进行种子萌发实验。每天观察记录种子的萌发情况,直到连续7 d内不再有种子萌发为止。
参照《国际种子检验规程》,以胚根突出种皮的长度为种子长度的一半视为发芽,统计发芽率与发芽势。

1.2.3 不同光照条件下的种子萌发实验 在25℃条件下,设置持续光照、14 h光照+10 h黑暗的周期性光照和全黑暗3组处理,进行种子发芽实验。每一处理随机选取50粒冷藏保存的种子,每个处理3次重复。每天观察记录种子的萌发情况,直到连续7 d内不再有种子萌发为止。
1.2.4 数据处理 对于上述属于二项分布的实验数据中百分率(p)大于70%或小于30%,根据方差分析的基本要求,数据采用了反正弦数据转换[arcsinp1/2][9]。采用SPSS17.0对转换后数据进行方差分析及多重比较。
2 结果与分析
2.1 种子形态
银缕梅种子光滑、褐色,种皮坚硬,并有一定厚度。银缕梅种子呈长纺锤型,千粒重为46.35 g,平均长径为8.08 mm,平均短径为3.79 mm,种粒大小和质量差异较小(表2)。

表2 银缕梅种子的物理性状Table 2 Physical properties of P. subaequalis seeds
2.2 不同处理方式对银缕梅种子发芽的影响
对9个正交处理组合的种子发芽率进行极差分析,结果见表 3。种子储藏方式、赤霉素浓度、温度的发芽率极差R分别为19.49、3.76、17.32,即种子储藏方式(A)>温度(C)>赤霉素浓度(B),表明种子储藏方式是影响银缕梅种子萌发的首要因素,其次为温度,赤霉素浓度对种子发芽率的影响较小。各因素的 k1、k2、k3数值表明:种子萌发的最佳组合为 A3B3C2。
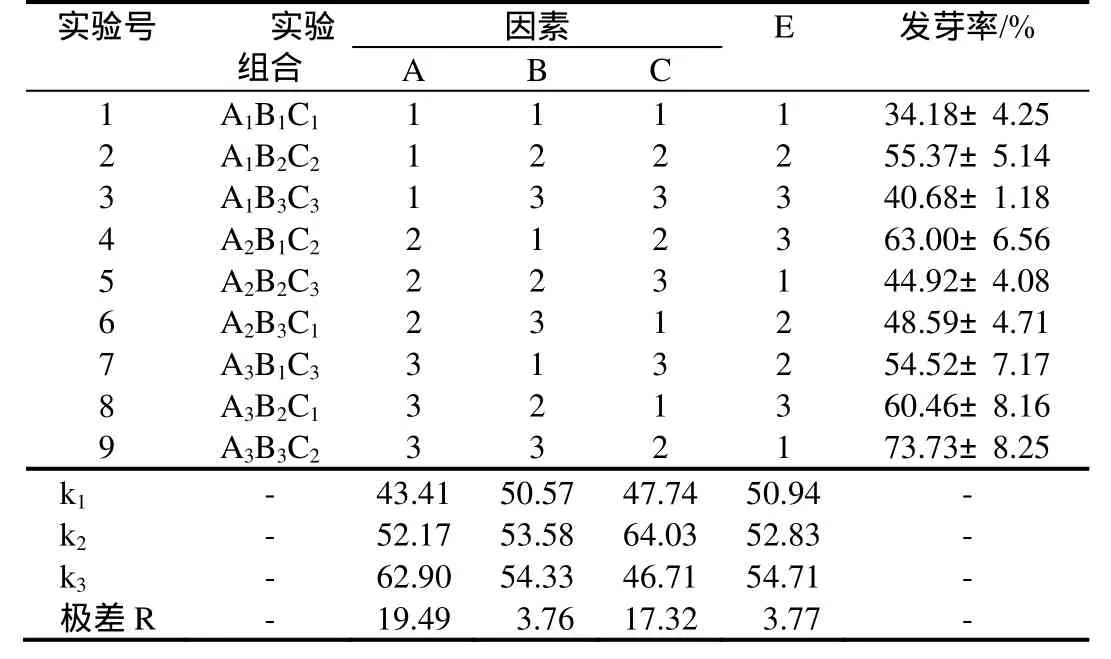
表3 银缕梅种子萌发正交实验结果Table 3 The results of orthogonal test of seed germination of P. subaequalis
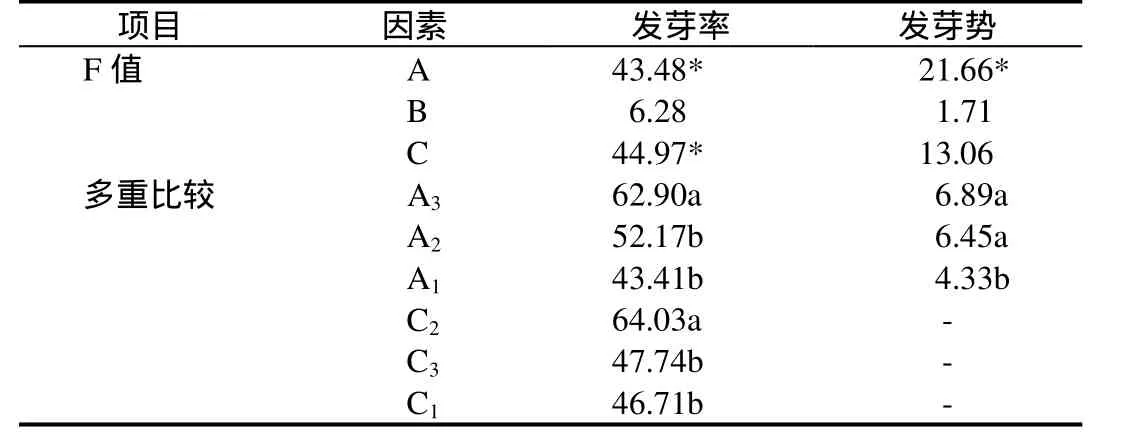
表4 各实验因素的方差分析及多重比较Table 4 ANOVA and multiple comparisons on experimental factors
各因素的方差分析及多重比较结果显示(表4):各储藏方式间种子发芽率与发芽势的F值分别为 43.48、21.66,差异显著;各赤霉素浓度间其F值分别为6.28、1.71,差异不显著;各温度间发芽率 F值 44.97,差异显著,但发芽势的 F值为13.06,差异不显著。实验结果表明,种子储藏方式和温度均对银缕梅种子发芽率有显著影响,在储藏方式中冷藏和常温沙藏对发芽势影响显著。
2.3 光照对银缕梅种子发芽的影响
在全光照、半光照半黑暗、全黑暗三种不同光照条件下,银缕梅种子14 d内萌发较平缓,而到21 d时表现出明显发芽高峰期,累积发芽率分别为37.50%、25.01%、8.33%。在这之后全光照条件下只有少量种子陆续萌发,最终发芽率仅为47.98%;半光照半黑暗条件下种子在28 d后萌发减少趋于稳定,发芽率达60.48%;而全黑暗条件下种子陆续萌发一直持续到 49 d,发芽率为52.08%(图1)。
由表5可看出,不同光照条件下种子发芽率及发芽势均无显著性差异。因此可推断半光照半黑暗条件下银缕梅种子发芽率最高,适度光照可缩短种子发芽时间,提高发芽势,利于种子发芽整齐。
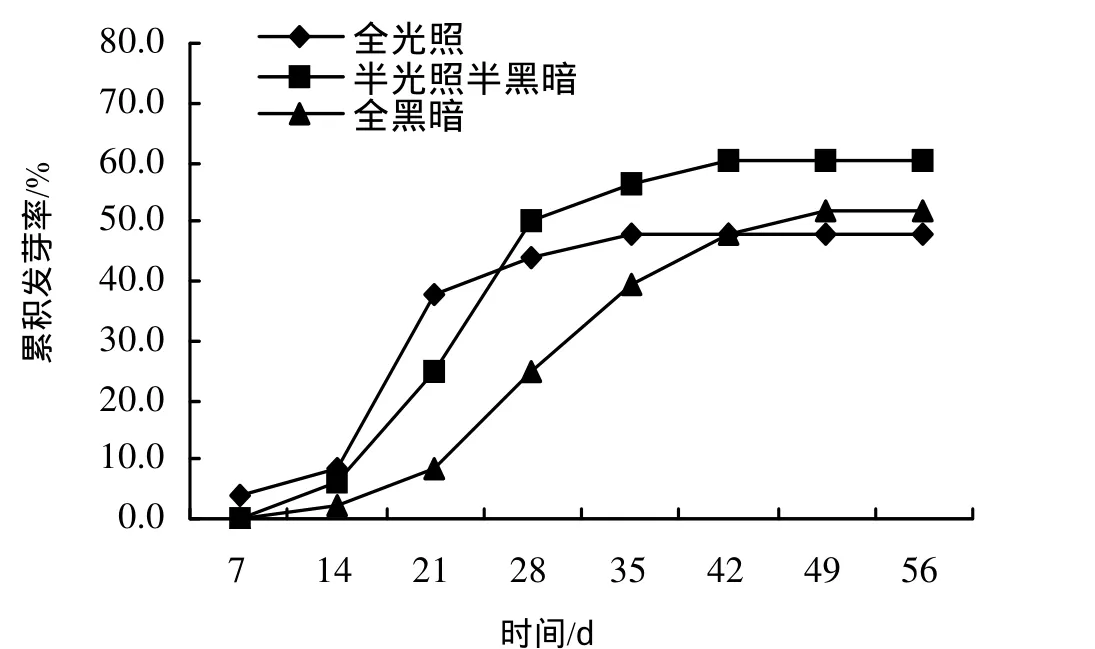
图1 不同光照条件下银缕梅种子发芽率Figure 1 Germination rates of P. subaequalis seed in different lighting conditions
3 讨论
温度作为外界条件对种子萌发起着至关重要的作用。适宜的温度可以打破种子休眠,增强种子酶活性,促进物质和能量的转化,表现出高发芽势和发芽率[10~11]。实验结果表明,温度对银缕梅种子萌发影响最大,在20 ~ 25℃发芽率最高,过高过低都会影响种子发芽率。因此建议在生产或实验中将银缕梅种子置于20 ~ 25℃萌发。
种子的休眠类型多样,大体可归于两类:一种是生理后休眠,包括胚未分化、胚生理后熟和抑制物质的作用。另一种是外致的强迫性休眠,包括光控制的暗休眠和种皮不透水性的机械阻碍作用[12]。研究表明,沙藏有利于种胚发育,可打破种子休眠,银缕梅种子以常温沙藏(120 d)发芽率最高,说明银缕梅种子存在一定休眠特性,这与张莹等在银缕梅引种栽培及种子营养成分分析结论相一致[13]。通常情况下,低温沙藏更有利于种子萌发,赖江山等[14]、史晓华等[15]认为低温层积可以显著提高种子发芽率和发芽势。银缕梅种子最适沙藏温度与时间有待进一步探讨。
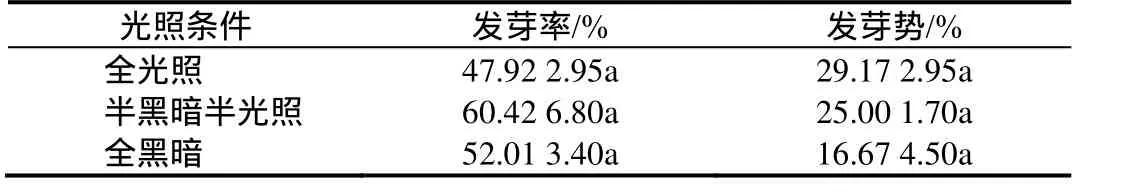
表5 不同光照条件对银缕梅种子发芽的影响Table 5 Influence to the germination of the seeds under the different light conditions
赤霉素溶液浸种处理可解除种子休眠,促进种子萌发[16~17]。实验结果显示赤霉素提高了银缕梅种子发芽率,但影响不显著,发芽率随赤霉素浓度增大而提高。银缕梅种皮坚硬,硬实率高,吸水困难,可能浸种时间较短使赤霉素对其影响不显著。机械或酸碱处理银缕梅种皮后浸种,增加赤霉素浸种时间等方法可能有助于其种子萌发。
光照对于银缕梅种子萌发是有利的,但不是主要条件。实验表明,银缕梅种子在全黑暗与有光照条件下的发芽率并没有显著差异,但光照可以缩短种子发芽时间,提高发芽势,利于种子发芽整齐。
笔者将常温储藏两年的银缕梅种子做了萌发实验,发芽率只有12.33%,推断其种子寿命较短。自然条件下,其种子有效保存会更短,一般只能在当年内萌发;银缕梅结实大小年现象明显,其种子长椭圆形、中粒、较坚硬,不易于水流、动物等传播;在龙王山保护区内采种银缕梅大树周围仅有少许幼苗,不利于种群的更替,可能是其致濒原因之一,这有待进一步研究。
[1]刘兴剑,郝日明. 小叶银缕梅[J]. 植物杂志,1999(4):6.
[2]ZhengYi W, Raven P H. Flora of China(Vol. 9)[M]. Beijing: Science Press, 2004.
[3]Yue C L, Jin S H, Chang J,et al. Response of photosynthesis in Shaniodendron subaequale to soil water status[J]. Ann Bot Fen, 2006,43(5):389-393.
[4]刘兴剑,汤诗杰,姚淦,等. 银缕梅开花过程与花形态观察[J]. 江苏农业科学,2008(6):165-166.
[5]颜超,王中生,安树青,等. 濒危植物银缕梅(Parrotia subaequalis)不同径级个体的光合能力差异与更新限制[J]. 生态学报,2008,28(9):4153-4161.
[6]朱汤军,岳春雷,金水虎. 银缕梅和伴生植物光合生理生态特性比较[J]. 浙江林学院学报,2008,25(2):176-180.
[7]姚志刚,王中生,颜超,等. 濒危植物银缕梅幼苗对不同光强的光合响应[J]. 南京林业大学学报(自然科学版),2010,34(3):83-88.
[8]李建华,A LinnBogle,Anita S Klein,等. 金缕梅科银缕梅属与帕罗堤属的亲缘关系——核糖体DNA ITS序列证据[J]. 植物分类学报,1997,35(6):481-493.
[9]盖钧镒. 试验统计方法[M]. 北京: 中国农业出版社, 1999.
[10]Maraghni M, Gorai M, Neffati M. Seed germination at different temperatures and water stress levels, and seedling emergence from different depths of Ziziphus lotus[J]. South Afr J Bot,2010,76(3):453-459.
[11]Kumar B, Verma S K, Singh H. Effect of temperature on seed germination parameters in Kalmegh (Andrographis paniculata)[J]. Ind Crops Prod,2011, 34(1):1241-1244.
[12]管康林. 种子生理生态学[M]. 北京: 中国农业出版社, 2009.
[13]张莹,李思锋,黎斌,等. 银缕梅引种栽培及种子营养成分分析[J]. 西北林学院学报,2011,26(4):148-151.
[14]赖江山,李庆梅,谢宗强. 濒危植物秦岭冷杉种子萌发特性的研究[J]. 植物生态学报,2003,27(5):661-666.
[15]史晓华,黎念林,金玲,等. 秤锤树种子休眠与萌发的初步研究[J]. 浙江林学院学报,1999,16(3):12-17.
[16]何志,唐宇丹,石雷,等. 猬实种子休眠特性研究[J]. 园艺学报,2008,35(10):1505-1510.
[17]何中声,刘金福,洪伟,等. 不同处理对格氏栲种子发芽的影响[J]. 北京林业大学学报,2012,34(2):66-70.
猜你喜欢
农业工程技术(2021年12期)2021-12-23
江西农业学报(2021年2期)2021-12-08
现代园艺(2020年19期)2020-10-02
河北林业科技(2020年3期)2020-03-23
种子(2019年7期)2019-08-13
种子(2019年6期)2019-07-13
种子(2019年4期)2019-05-28
现代农村科技(2019年4期)2019-01-06
现代农业科技(2016年8期)2016-10-20
浙江大学学报(农业与生命科学版)(2015年4期)2015-12-15
