
两种喀斯特生境植物旱雨季生理指标特征
2012-07-16 07:37:58曹洪麟吴林芳
山东农业大学学报(自然科学版) 2012年1期
董 蕾,曹洪麟 ,吴林芳
(1.中国科学院华南植物园,广州 510650;2.中国科学院研究生院,北京 100039)
喀斯特地貌具有特殊的生境特征,表现为土壤干旱瘠薄富钙、土被不连续和生境异质性大等特征,这一系列生境特征决定喀斯特适生植物在生理上表现出耐干旱瘠薄的生理特征[1]。该生境旱雨季降水差异极大,4~9月降水量占全年总降水量的83%[2,3]。这种水分差异必然造成植物的生理指标差异。
目前喀斯特生理生态研究焦点集中于资源与环境胁迫的相互作用[4,5,6],对喀斯特适生植物季节性生理指标变化的研究鲜有报道。本文选取红背山麻杆(Alchornea trewioides Muell.)和桂花(Osmanthus fragrans Lour.)为研究对象。前者为广西木论喀斯特次生林、灌草丛的先锋种和优势种[7],后者有较高的园林、药用、日用化工价值[8]。为了解喀斯特生境植物旱雨季的生理变化,本研究对这两种植物在旱、雨季的饱和净光合速率(Pmax)、比叶面积(SLA)、叶片总氮量(Nmass-leaves)、茎总含氮量(Nmass-sterm)、叶绿素 a、b总量(Chla+b)及其比值(Chla/b)进行测定,以期为喀斯特地区植被恢复提供理论指导和数据支撑。
1 材料和方法
1.1 研究地点及材料
研究地点位于粤北阳山地区(2007年4月)。广东省阳山县溶岩分布广泛,岩溶地貌发育完全,属峰丛洼地-峰丛谷地型岩溶[9]。整体地貌为低山丘陵,土壤结构简单,为由白云岩、石灰岩风化形成的石灰土,土壤发育不全,土层较薄,深浅不一。研究对象:桂花(Osmanthus fragrans Lour.),木犀科常绿乔木;红背山麻杆(Alchornea trewioides Muell.),大戟科山麻杆属落叶小乔木。在调查区随机选取成熟植株每种各15棵,连同根部土壤带回广州华南植物园,栽入盆中种植。
1.2 实验方法
实验分为雨、旱两季进行,雨季为2008年3-5月(每天晚上5点浇水至盆底有水流出),旱季为2008年9~11月(转移至温室,每周浇水1次),每月10号、20号、30号各进行下述实验1次。
最大光合速率(Pmax)测定:每次试验在早八点进行,利用LI-6400便携式光合作用仪(LI-COR,美国)测定最大光合速率(PAR=2000mol photons s-1)。每棵植株选上、中、下不同位置成熟叶片各3片,每个树种各15株。
叶绿素提取:将测完Pmax的叶片摘下,迅速用面积为0.2743 cm2的打孔器打孔3~4片称重后放入5 mL试管,每管加入80%丙酮5 mL黑暗浸提60 h至叶片基本变白。利用UV-3802型紫外可见分光光度计(Unico,美国)测定提取液A663、A646处吸光值(80%丙酮为空白对照),根据Lichenthaler[10]对Arnon法的修正,其计算公式为:Chla(mg/g)=12.21A636-2.81A646;Chlb(mg/g)=20.13A646-5.03A663
比叶面积(SLA)测定:利用光电叶面积仪(WDY-500A)测定打孔叶片面积,烘干(105℃杀青30 min,80℃连续烘干至恒重)后分别测量干重。利用计算公式(SLA(cm2/g·dw)=叶面积/干重)计算每叶片的 SLA 指标[11]。
叶氮含量(Nmass-leaves)测定:烘干叶片利用LK-200A型中药粉碎机磨成粉末,过60目筛,准确称取0.5g。凯氏定氮法(Kjeldahl method)消煮,消煮所得溶液利用流动注射法测其含氮量(百分含量)。
茎氮含量(Nmass-sterm)测定:每月末取每个物种的一棵植株,分上、中、下三段测量。测定方法与叶氮含量测定相同。
1.3 结果处理
数据利用SPSS 12.0软件进行分析,方差分析采用one-way ANOVA法。
2 结果与分析
2.1 比叶面积(SLA)及最大光合速率(Pmax)季节性变化
比叶面积(SLA)可以有效反映植物获取资源的能力[12]。两种植物的比叶面积在旱季均呈下降趋势(图1A)。其次,桂花的比叶面积在旱、雨季均显著低于红背山麻杆(P<0.05)。这说明红背山麻杆在受试季节的获取资源能力较桂花强。最大光合速率(Maximum photosynthesis rate,Pmax)是决定植物光合特征的关键性参数之一[13,14]。在研究期间,桂花的最大光合速率(Pmax)(雨季:8.5 ±1.5 μmol m-2·s-1;旱季:3.8 ±0.4 μmol m-2·s-1)均低于红背山麻杆(雨季:10 ±1.8 μmol m-2·s-1; 旱季:10.8 ±1.2 μmol m-2·s-1);且相比雨季,桂花旱季的最大光合速率显著降低(P<0.1),而红背山麻杆保持不变甚至略有升高(图1B)(P>0.05)。这可能与植物生活史特性有关。红背山麻杆在旱季凋落叶片,进入避旱期,所以它需要制造和储存更多的光合产物用以度过旱季。
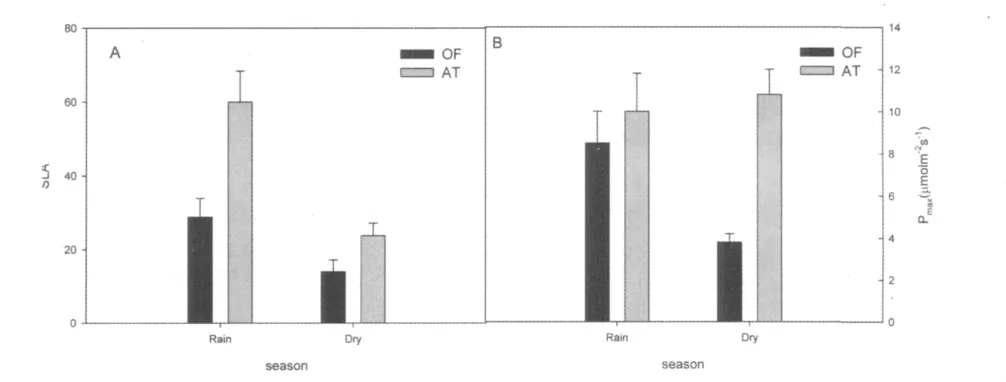
图1 两种植物旱雨季SLA(A)及最大光合速率(Pmax)(B)变化OF:桂花;AT:红背山麻杆Fig.1 SLA(A)and maximum leaf photosynthetic rate(Pmax)(B)of two species in two seasons.Corner marker:OF:Osmanthus fragrans Lour.;AT:Alchornea trewioides Muell
2.2 季节性叶绿素总量(Chla+b)及其Chla、b比值(Chla/b)变化
叶绿素含量受生境影响会发生显著变化[15]。叶绿素a、b含量随季节不同会发生变化,且叶绿素a比叶绿素b对干旱胁迫更加敏感[16],导致叶绿素a/b(Chla/b)值也随之变化。雨季两种植物的Chla/b并无显著性差异,旱季该指标均降低,且桂花(2.41±0.57)显著低于红背山麻杆(3.21±0.32)(图2A)(P<0.1)。叶绿素是高等植物同化器官的重要组成部分,决定植物光合能力[17]。叶绿素含量因生境差异而发生显著变化[15]。桂花雨季叶绿素含量高于旱季,红背山麻杆则相反。两者相比,雨季桂花叶绿素含量(3.11 ±0.31)显著高于红背山麻杆(1.34 ±0.41)(图2B)(P <0.1)。结合饱和光合速率(图1B)可以看出桂花虽然雨季叶绿素含量显著高于红背山麻杆(P<0.1),但其光合能力仍相当(P>0.05),即桂花的叶绿素光合效率低于红背山麻杆。另外,红背山麻杆旱季叶绿素含量的上升可能是其光合速率(图1B)上升的原因。

图2 两种植物雨旱季Chla/b(A)及Chla+b(B)变化 OF:桂花;AT:红背山麻杆Fig.2 the content of chlorophyll(A)and the ratio of Chla and Chlb(B)of two species in two seasons Corner marker:OF:Osmanthus fragrans Lour.;AT:Alchornea trewioides Muell.
2.3 季节性茎氮含量(Nmass-sterm)及叶氮含量(Nmass-leaves)变化
氮是植物必需元素,植物的茎具有储藏和繁殖的功能[18]。图3A中桂花(0.811±0.070)和红背山麻杆(1.083 ±0.10)旱季茎氮含量均高于雨季(桂花:0.673 ±0.087; 红背山麻杆:1.008 ±0.070),但桂花的增幅较红背山麻杆显著(P<0.1),即说明桂花转移的氮量较多。
叶氮含量直接影响植物体内叶绿素、可溶蛋白及酶,从而调节光合作用[19]。单位面积氮含量高的叶片具有较高光合能力[20,21,22]。叶片氮含量的季节性变化与SLA变化相似。两种植物叶氮含量旱季(桂花:1.2 ±0.5; 红背山麻杆:1.82 ±0.41)均低于雨季(桂花:1.6 ±0.35; 红背山麻杆:2.1 ±0.2),且红背山麻杆在两个季节都高于桂花(图3B)。
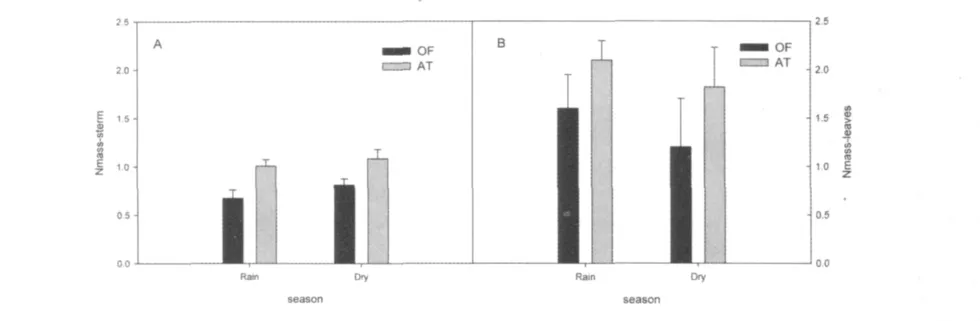
图3 两种植物雨旱季茎氮含量(A)及叶氮含量(B)变化OF:桂花;AT:红背山麻杆Fig.3 the content of nitrogen in sterm(A)and content of nitrogen in leaves(B)of two species in two seasons Corner marker:OF:Osmanthus fragrans Lour.;AT:Alchornea trewioides Muell.
3 讨论
最大光合速率(Maximum photosynthesis rate)能够显示植物对气候变化的响应[23,24,25,26]。在旱季桂花叶绿素含量降低,光合能力下降,叶氮含量降低,叶片中营养转移至茎(图1,2,3)。由于是常绿植物,桂花只能选择忍耐策略应对干旱,这可能是其转移营养物质,降低消耗度过旱季的方法。
红背山麻杆为落叶植物,其进入旱季后将有6个月无法进行光合,故其将叶氮一部分用来制造叶绿素从而提高光合能力,另一部分转移至茎进行储存(图1,2,3)。SLA及叶氮含量高表征具有较高相对潜在生长率[27]。从图1可以看出,红背山麻杆雨季有较高SLA和叶氮含量,这说明它在雨季潜在生长率较高,这可能是因为红背山麻杆在每年雨季重新萌发新叶,且仅有6个月时间进行营养制造,需要利用6个月制造的营养物质维持一年的生长。因此在喀斯特特殊生境下,红背山麻杆比常绿植物桂花更具适应能力。这与木论喀斯特森林调查结果:落叶植物比常绿植物比重大、红背山麻杆为次生林、灌草丛的先锋种、优势种的结果相符[7]。
[1]黄威廉,屠玉麟.贵州植被[M].贵阳:贵州人民出版社,1988.1-20
[2]何腾兵.贵州喀斯特山区水土流失状况及生态农业建设途径探讨[J].水土保持学报,2000,14(5):28-34
[3]彭建,杨明德.贵州花江喀斯特峡谷水土流失状态分析[J].山地学报,2001,19(6):511-515
[4]何跃军,钟章城,刘济明,等.石灰岩退化生态系统不同恢复阶段土壤酶活性研究[J].应用生态学报,2005,16(6):1077-1081
[5]谢宗强,江明喜.三峡地区石灰岩灌丛植被特征及其合理利用[J].植物学通报,2005,12:85-89
[6]朱守谦.茂兰喀斯特森林小生境特征研究[M].贵阳:贵州科技出版社,2003.38-48
[7]何加林,曹洪麟,张燕婷,等.广西木论喀斯特森林土壤养分水平与植被及地形的关系[J].热带亚热带植物学报,2009,17(5):502-509
[8]鄢洪星,黄义林,周传涛,等.桂花的经济价值及其繁育技术[J].现代农业科学,2009,1:62-63
[9]曹晓娟 ,董 颖.广东阳山国家地质公园内地质遗迹特征[J].中国地质灾害与防治学报,2010,21(2):125-128
[10]Lichtenthaler H K.Chlorophylls and carotenoids:pigments of photosynthetic biomembrane[J].Methods Enzymol,1987,148:350 -382
[11]李轩然,刘琪璟,蔡 哲,等.千烟洲针叶林的比叶面积及叶面积指数[J].植物生态学报,2007,31(1):93-101
[12]Lambers H,Poorter H.Inherent variation in growth rate between higher plants:a search for physiological causes and ecological consequences[J].Advances in Ecological Research,1992,23:188 -242
[13]Ruimy A,Jarvis P G,Baldocchi D D,et al.CO2fluxes over plant canopies and solar radiation:a review[J].Advance in Ecological Research,1995,26:1-68
[14]Schulze E D,Caldwell M M .Ecophysiology of photosynthesi.Berlin,Heidelberg[M],Germany:Springer-verlag,1994
[15]Ferus P,Arkosiova M.Variability of chlorophyll content under fluctuating environment[J].Acta fytotechnica et zootechnica,2001,4:123 -125
[16]林植芳,李双顺,林桂珠.水稻叶片的衰老与超氧物歧化酶活性及脂质过氧化作用的关系[J].植物学报,1984,26(6):605-615
[17]Uvalle Sauceda JI,H Gonzalez Rodriguez,RG Ramirez Lozano,et al.Seasonal trends of chlorophylls a and b and carotenoids in native trees and shrubs of Northeastern Mexico[J].Boil Sci,2008,8:258 -267
[18]李扬汉.植物学[M].上海:科学技术出版社,1984:112-113
[19]赵 平,孙谷畴,彭少麟.植物氮素营养的生理生态学研究[J].生态科学,1998,17(2),37-42
[20]Evans J R.Photosynthesis and nitrogen relationships in leaves of C3 plants[J].Ocologia,1989,78:9 -19
[21]Field C and Mooney H A.Leaf age and seasonal effects on light water and nitrogen efficiency in a California shrub[J].Oecologia,1983,56:348-355
[22]Field C A,Mooney H A.The photosynthesis-nitrogen relationship in wild plants.In T J Givnish ed.on the Economy of Plant Form and Function.Cambridge:Cambrige University Press,1986.25-55
[23]Gardiner E S,Krauss K W.Photosynthetic light response of flooded cherry barks oak(Quercus pagoda)seedlings grown in two light regimes[J].Tree Physiol,2001,21(15):1103 -1111
[24]Herrick J D,Thomas R B.Effects of CO2 enrichment on the photosynthetic light response of sun and shade leaves of canopy sweetgum trees(Liquidambar styraciflua)in a forest ecosystem[J].Tree Physiol,1999,19(12):779 -786
[25]Koike T,Lei T T,Maximov T C,et al.Comparison of the photosynthetic capacity of Siberian and Japanese birch seedlings grown in elecated CO2 and temperature[J].Tree Physiol,1996,16(3):381 - 385
[26]Lewis J D,Mckane R B,Tingey D T,et al.Vertical gradients in photosynthetic light response within an old-growth Douglas-fir and western hemlovk canopy[J].Tree Physiol,2000,20(7):447 - 456
[27]PB Reich,D.S.Ellsworth and MB Walters.Leaf structure(specific leaf area)modulated photosynthesis- nitrogen relations:evidence from within and across species and functional groups[J].Functional ecology,1998,12:948-958
猜你喜欢
儿童时代·快乐苗苗(2022年9期)2022-10-31 09:57:52
湖南农业大学学报(自然科学版)(2021年3期)2021-07-02 01:48:06
小读者(2021年2期)2021-03-29 05:03:12
阅读(低年级)(2020年8期)2020-11-06 06:23:19
新疆农业科学(2020年1期)2020-02-14 03:22:46
乡村地理(2019年2期)2019-11-16 08:49:32
————水溶蚀岩石的奇观
家教世界(2019年4期)2019-02-26 13:44:20
读友·少年文学(清雅版)(2018年10期)2019-01-29 06:57:58
小学生优秀作文(低年级)(2018年10期)2018-10-13 01:56:46
青苹果·教育研究版(2016年7期)2016-12-12 05:19:52
