
大岗山林区几种常见灌木生物量估算与分析1)
2012-07-02 00:07郭泉水牛树奎张志旭
东北林业大学学报 2012年9期
赵 蓓 郭泉水 牛树奎 孙 武 张志旭
(北京林业大学,北京,100083)(中国林业科学研究院森林生态环境与保护研究所)(北京林业大学)
生物量测定被广泛应用于森林生态系统生产力和群落动态特征等方面的研究[1],同时也被应用于分析比较森林资源价值、森林碳储量、群落演替、退化生态系统的恢复等方面[2-6]。因此,生物量的准确测定是深入研究许多生态和林业问题的基础[7-8]。灌木层是森林群落中重要的结构成分,其生物量的研究对于整个森林生态系统物质和能量的固定、消耗、积累和转化有着极为重要的意义[9-11]。目前,灌木生物量测定采用的主要方法为直接收割法,此方法效率较低、破坏性较大[12]。因此,研究简便易行的灌木生物量测定方法——生物量模型,即采用易测因子来估算灌木器官以及全株生物量,近年来受到众多学者的关注[13-17]。目前,研究中采用较多的易测因子,包括植株高度、基径,而实际观测表明,植株冠幅直径与生物量的关系也极为密切[18],冠幅直径和株高能够很好地反映灌木的形态特征,因此,以株高、基径和冠幅直径来拟合生物量模型可以建立更准确的生物量估测模型。
本研究对于更深入了解大岗山林下灌木层的生物学特性,以及进一步研究整个大岗山林区灌木生物量及其资源价值具有十分重要的意义。然而,以往对大岗山林下灌木层生物量的研究较少考虑树种间或者不同器官间最优生物量模型的差异[19],因此,本研究以大岗山年株林场3种典型森林类型(常绿阔叶林、人工杉木林、天然毛竹林)为研究对象,对林下灌木层常见的10种灌木构建单种各器官和全株生物量模型,并应用这些模型估算不同森林类型下这10种常见灌木的总生物量。研究结果将为准确估计大岗山林下灌木生物量提供科学依据,同时,为评价本地区林下常见资源植物价值和不同森林类型的生态服务功能提供参考。
1 研究区概况
研究区位于江西省分宜县境内中西部的大岗山年株林场,北纬 27°30'~ 27°50',东经 114°30'~114°40'。地貌形态主要为低山丘陵,区内最高峰海拔1 091.8 m。该区地处中亚热带,属于中亚热带季风湿润气候类型,年平均气温为16.8℃,≥10℃积温为5 355℃,年平均降水量为1 590.9 mm,全年平均日照时数为1 657.0 h,年均蒸发量为1 503.8 mm,年平均相对湿度为81%。土壤属黄壤系列的山地土壤,土质疏松,肥沃湿润,腐殖质层中等厚度,土体厚度一般在60~80 cm。研究区内的森林类型主要为常绿阔叶林、人工杉木林、天然毛竹林。主要树种有:丝栗栲(Castanopsis fargesii)、苦槠(Castanopsis sclerophylla)、檫木(Sassafras tsumu)、拟赤杨(Alniphyllum fortunei)、杉木(Cunninghamias lanceolata)、毛竹(Phyllostachys heterocycla)。
2 材料与方法
2.1 数据的获取
以大岗山年株林场3种森林类型常绿阔叶林、人工杉木林、天然毛竹林为研究对象,各森林类型设置3块不同密度的样地,在各样地的4个角和中心分别设置2 m×2 m的灌木样方,统计群落物种组成,分种测量每株灌木的冠幅直径(C)(冠幅为长短冠幅平均值)、株高(H)、基径(D)。根据样方内灌木调查结果,选择样地中频度较高的10个种(表1),每个种在样地附近选取具有代表性植株20~30株,测量株高、冠幅、基径。之后整株挖出,深度为根系分布所达范围,地上部分分为叶、枝、干,称鲜质量(精确到0.1 g),迅速去除根系上附着的泥土后称其鲜质量(精确到0.1 g),各部分分别取样装入信封称质量,并带回实验室在烘箱内105℃烘至质量不再变化,称干质量,计算干鲜质量比,通过干鲜质量比和野外调查的植株各部分的鲜质量,计算每株灌木各器官以及全株干质量。10种灌木的样本数及各调查参数的平均值及标准差见表1。

表1 取样灌木的基本参数
2.2 模型建立方法
用 SPSS15.0[20]软件做回归分析,选用变量:H(植株高度/m);D(植株基径/cm);C(植株冠幅直径/m);A(植冠面积/m2,A=πC2/4);V(植株体积/m3,V=AH);DH(基径与株高乘积/m2);D2H(基径平方乘以株高/m3);W(生物量/kg,分为叶、枝、干、根和全株)。对灌木单株各器官及全株生物量进行分析,拟合生物量模型,选取常用函数[21]:
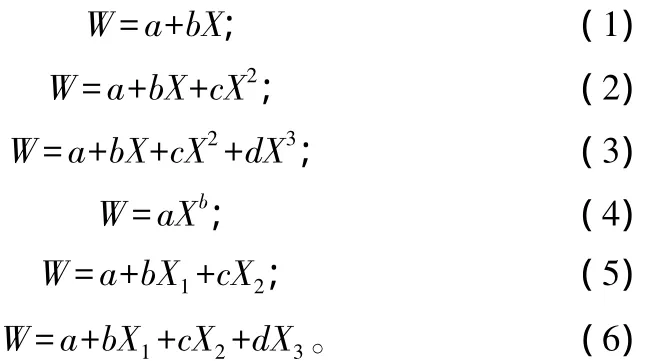
式中:a为常数项;b、c、d分别为系数;X 为 D、H、C、A、V、DH、D2H 中的任意 1 个;式(5)中的 X1、X2为D、H、C、A、V、DH、D2H 中的任意 2 个;式(6)中的X1、X2、X3为 D、H、C、A、V、DH、D2H 中的任意 3 个。对拟合的回归方程均进行t检验,用判定系数(R2)和估计的标准差(SE,E)作为选取最优模型的评价指标[22]。选取相关性最密切、拟合度最好的模型,即R2最大,SE,E最小的模型作为生物量估算的最优模型。将拟合得到的灌木单株器官生物量模型用于计算大岗山常绿阔叶林、人工杉木林、天然毛竹林下这10种灌木的生物量。
3 结果与分析
3.1 单株器官生物量模型
3.1.1 叶生物量模型
各灌木种的叶生物量模型回归分析结果表明,叶生物量与各预测变量之间相关性较高,与各预测变量组成的线性和非线性模型的拟合效果都较好,R2均在0.79以上。各灌木种的叶生物量最优模型形式和采用的自变量不同,草珊瑚、杜茎山、黄瑞木、丝栗栲、油茶和朱砂根叶生物量模型以W叶=a+bV+cV2+dV3相关性最高;矩形叶鼠刺和毛冬青叶生物量模型以W叶=a+bA+cA2+dA3相关性最高;柃木叶生物量模型以W叶=a+bV+cV2相关性最高;檵木叶生物量模型以W叶=a+b(D2H)+c(D2H)2相关性最高。具体模型参数及显著性检验结果见表2。
3.1.2 枝生物量模型
灌木枝生物量模型拟合结果显示,除黄瑞木枝生物量最优模型自变量为D外,其他灌木种的枝生物量最优模型采用的自变量多为DH或者V,最优模型形式多为二次或者三次方程。草珊瑚、杜茎山、丝栗栲、油茶和朱砂根以方程W枝=a+bV+cV2+dV3拟合效果最好;檵木和矩形叶鼠刺以 W枝=a+b(DH)+c(DH)2+d(DH)3拟合效果最佳;黄瑞木以W枝=a+bD+cD2+dD3拟合效果最优;柃木枝生物量最优模型以W枝=a+b(DH)+c(DH)2拟合效果最好;毛冬青以W枝=a+bV+cV2拟合效果最佳。经t检验均呈极显著相关。具体模型参数及显著性检验结果见表3。
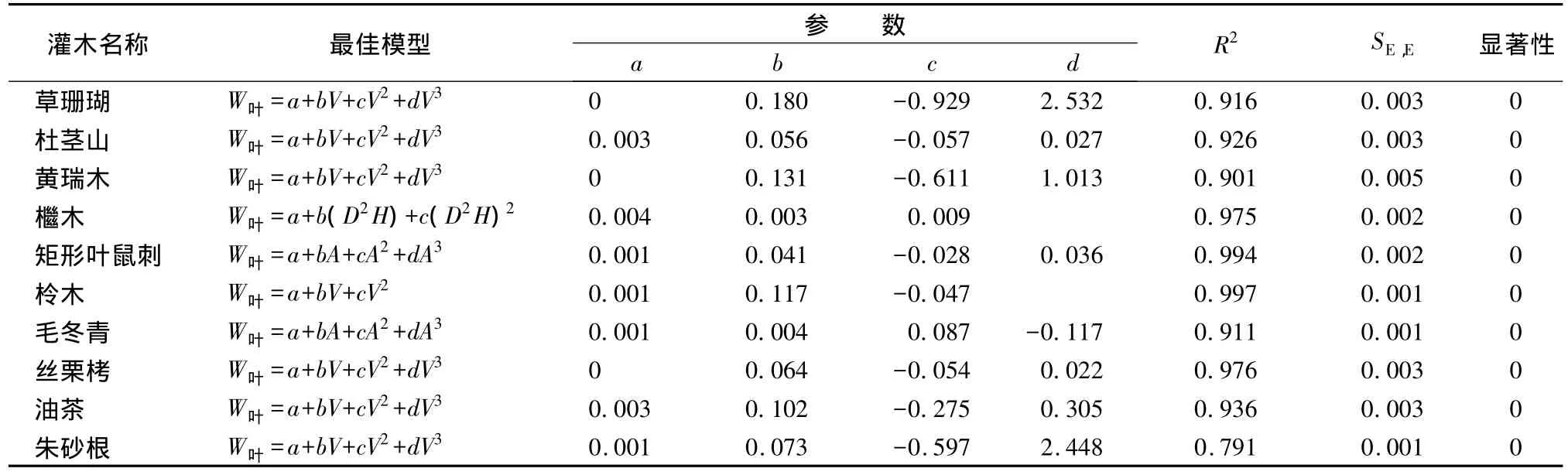
表2 叶生物量最优模型
3.1.3 干生物量模型
灌木干生物量模型拟合结果显示,除杜茎山、毛冬青外各灌木种的干生物量最优模型采用的自变量均为DH,草珊瑚、檵木、矩形叶鼠刺、柃木、丝栗栲、油茶和朱砂根干生物量最优模型为W干=a+b(DH)+c(DH)2;杜茎山和毛冬青干生物量最优模型为W干=a+b(D2H)+c(D2H)2+d(D2H)3;黄瑞木干生物量最优模型为W干=a(DH)b。经t检验均呈极显著相关。具体模型参数及显著性检验结果见表4。
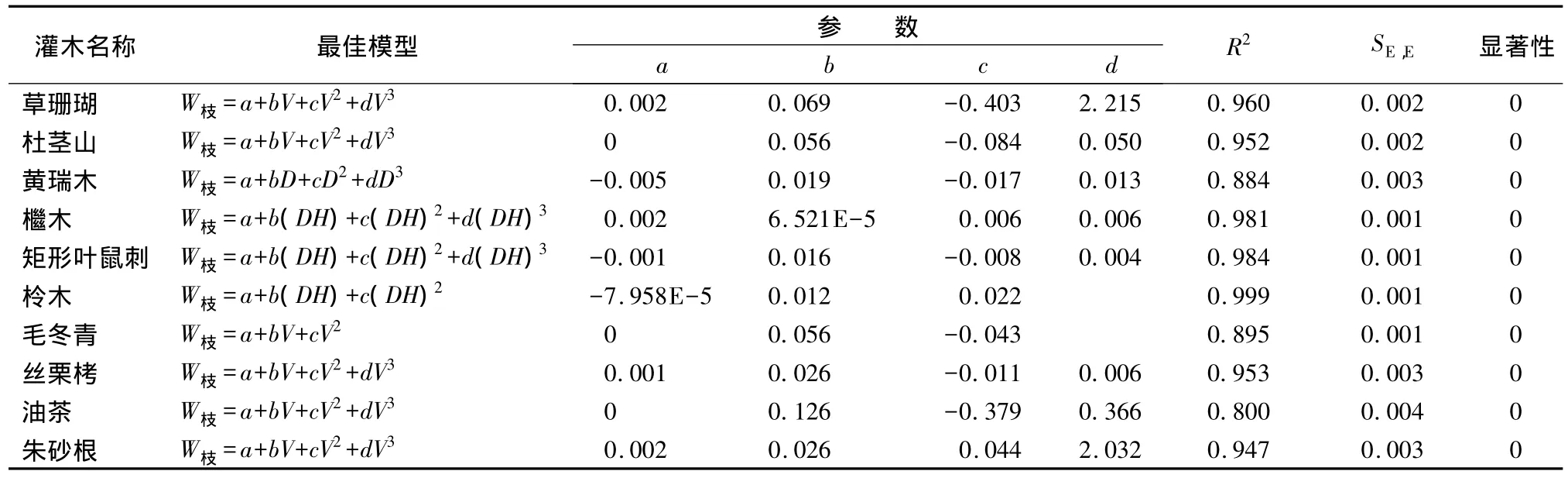
表3 枝生物量最优模型
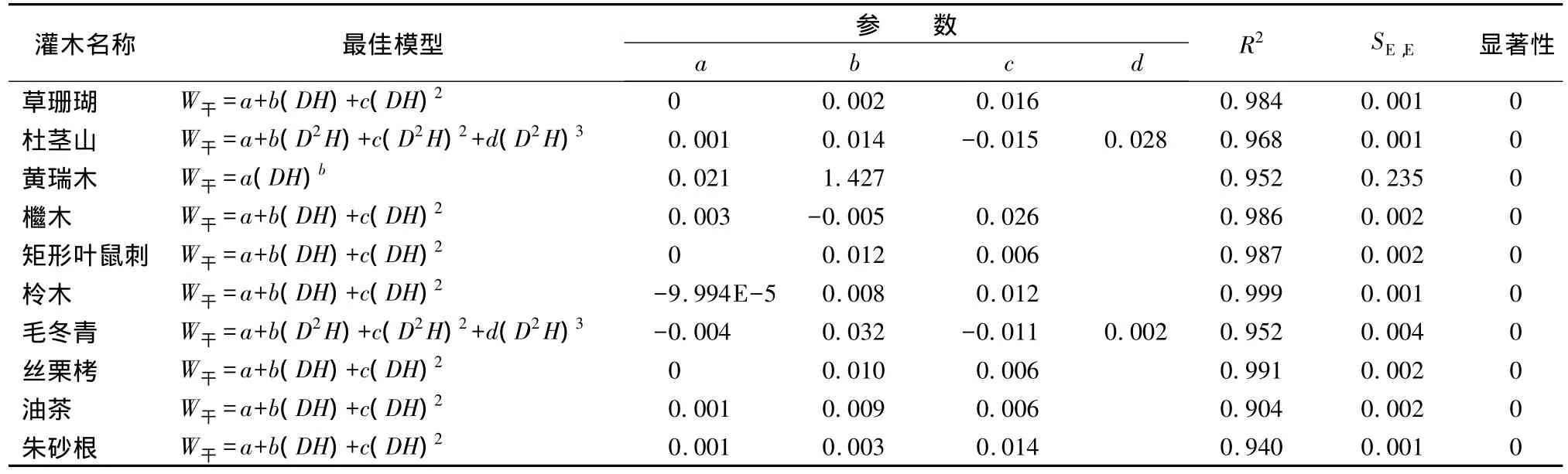
表4 干生物量最优模型
3.1.4 根生物量模型
各灌木种的根生物量模型回归分析结果表明,各模型均具有较高的R2值和较小的SE,E值,R2均在0.76以上。各灌木种根最优模型形式和采用的自变量不同,最优模型形式多为二次或者三次方程,杜茎山、黄瑞木模型形式为幂函数。经t检验均呈极显著相关。具体模型参数及显著性检验结果见表5。
3.2 全株生物量模型
将各灌木种单株总生物量与各预测变量进行回归分析,建立全株生物量模型。经筛选,草珊瑚、杜茎山、矩形叶鼠刺、丝栗栲和油茶全株生物量以W全株=a+bV+cV2+dV3为最优模型;黄瑞木和毛冬青全株生物量以 W全株=a+b(D2H)+c(D2H)2+d(D2H)3为最优模型;檵木和柃木全株生物量以W全株=a+b(D2H)+c(D2H)2为最优模型;朱砂根全株生物量以W全株=a+bV+cV2为最优模型。经t检验均呈极显著相关。具体模型参数及显著性检验结果见表6。
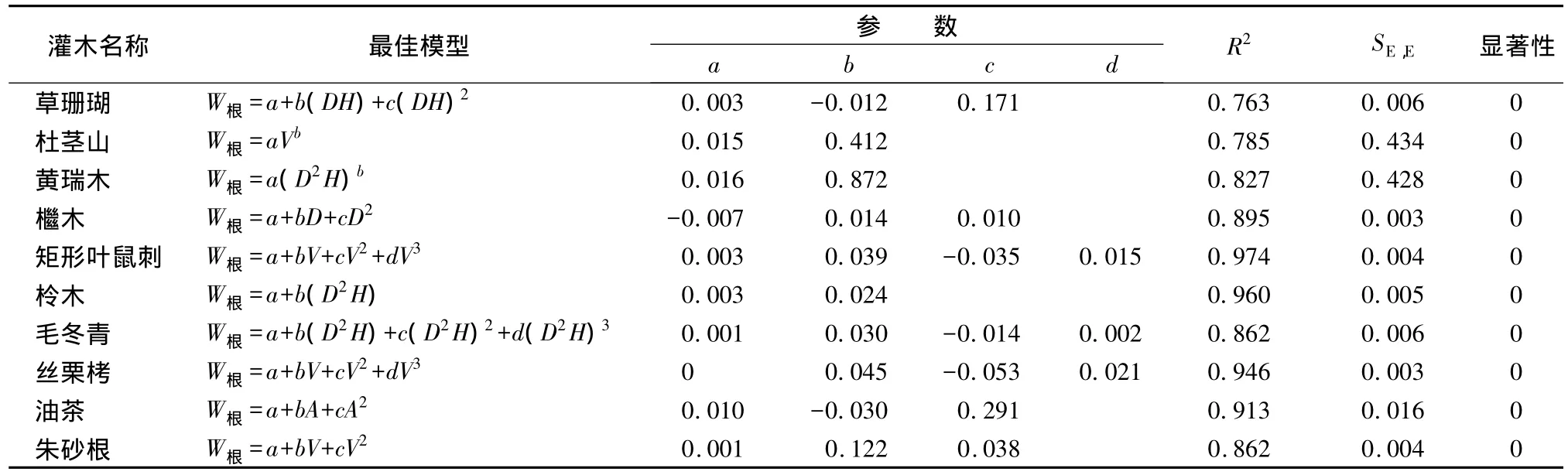
表5 根生物量最优模型

表6 全株生物量最优模型
3.3 单株器官生物量模型的应用
将拟合得到的各灌木种的器官生物量模型用于3种林型下各灌木树种生物量的估算。各森林类型调查情况详见表7。

表7 不同森林类型物种及其密度分布
根据计算各灌木种在毛竹林下的生物量情况可知,各灌木种在毛竹林下的生物量由大到小依次为:杜茎山、矩形叶鼠刺、丝栗栲、黄瑞木、柃木、毛冬青、檵木、油茶、朱砂根、草珊瑚。各灌木种在毛竹林下的生物量情况详见表8。
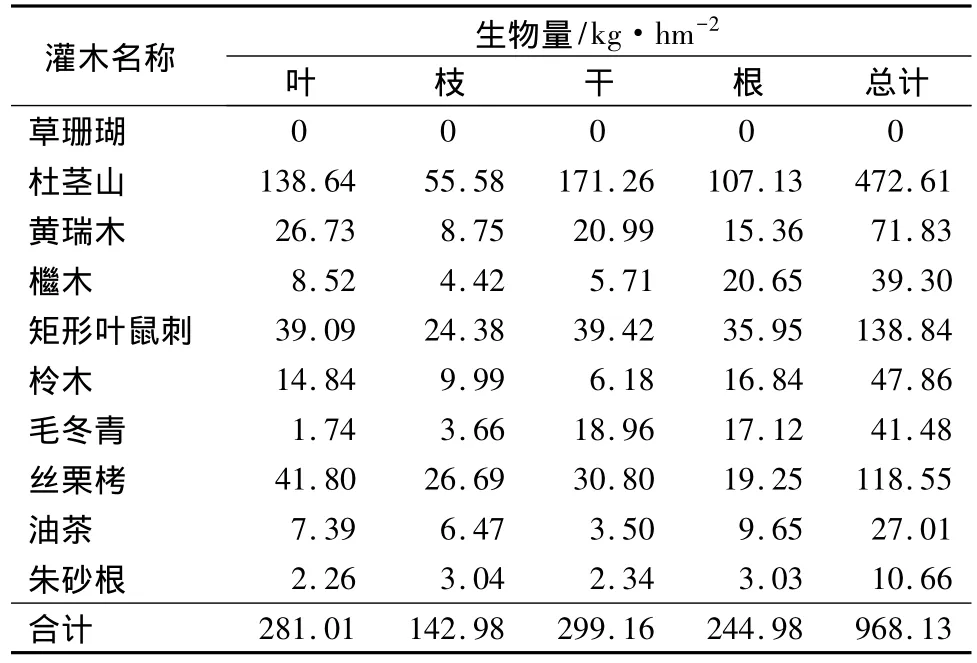
表8 各灌木种在毛竹林下生物量情况
根据计算各灌木种在人工杉木林下的生物量情况可知,各灌木种在人工杉木林下的生物量由大到小依次为:杜茎山、柃木、檵木、丝栗栲、矩形叶鼠刺、黄瑞木、油茶、朱砂根、毛冬表、草珊瑚。各灌木种在人工杉木林下的生物量情况详见表9。
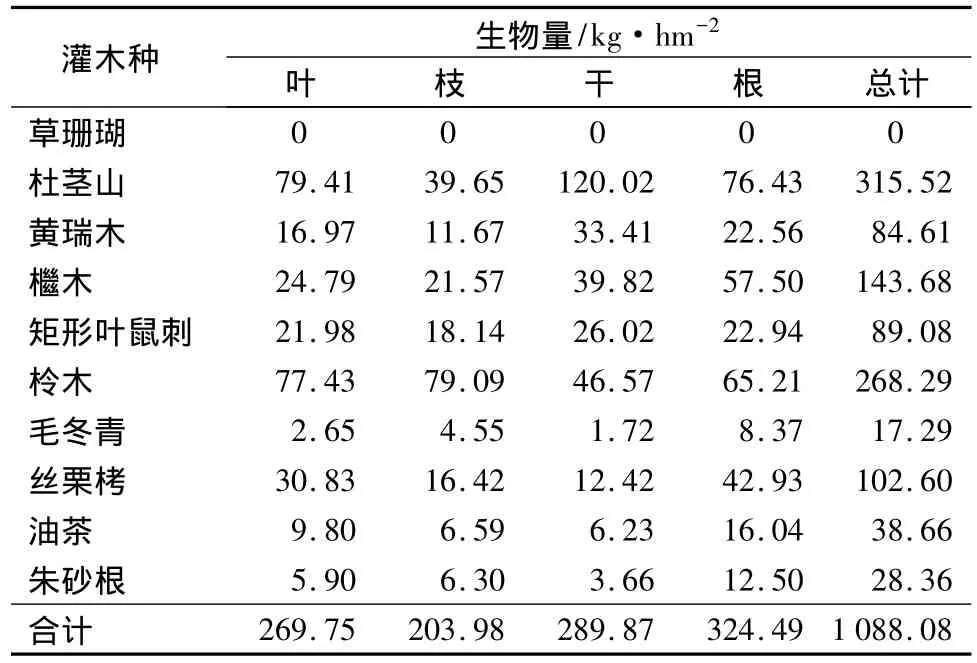
表9 各灌木种在人工杉木林下生物量情况
根据计算各灌木种在常绿阔叶林下的生物量情况可知,各灌木种在常绿阔叶林下的生物量由大到小依次为:杜茎山、黄瑞木、草珊瑚、柃木、檵木、矩形叶鼠刺、油茶、毛冬表、朱砂根、丝栗栲。各灌木种在常绿阔叶林下的生物量情况详见表10。

表10 各灌木种在常绿阔叶林下生物量情况
由表8、表9、表10可知,10种灌木中杜茎山在毛竹林、人工杉木林和常绿阔叶林中的生物量都很高,且杜茎山在常绿阔叶林中的生物量最大;草珊瑚在毛竹林、人工杉木林中没有分布,在常绿阔叶林中生物量为143.63 kg·hm-2;黄瑞木在常绿阔叶林中生物量最大;檵木在人工杉木林中生物量最大;矩形叶鼠刺在毛竹林中生物量最大,在常绿阔叶林中最小;柃木在人工杉木林中生物量最大,毛竹林中最小;毛冬青在毛竹林中生物量最大;丝栗栲在毛竹林中生物量最大,在常绿阔叶林中最小;油茶在常绿阔叶林中生物量最大,在毛竹林中最小;朱砂根在毛竹林中生物量最小,在人工杉木林和常绿阔叶林中基本相等。
由表8、表9、表10可知,调查的10种灌木在常绿阔叶林中总生物量最大,人工杉木林中次之,毛竹林中最小,依次为 1 284.38、1 088.08、968.13 kg·hm-2。各灌木不同器官总生物量比较发现,常绿阔叶林中叶的生物量占总生物量的比例最大,为389.52 kg·hm-2,其次是根、干,分别为 360.80,344.39 kg·hm-2,枝的总生物量最小,为 189.66 kg·hm-2;毛竹林中干的生物量占总生物量的比例最大,为299.16 kg·hm-2,其次是叶、根,分别为 281.01、244.98 kg·hm-2,枝的生物量最小,为 142.98 kg·hm-2;人工杉木林中根的生物量占总生物量的比例最大,为324.49 kg·hm-2,其次是干、叶,分别为 289.87、269.75 kg·hm-2,枝的生物量最小,为 203.98 kg·hm-2。
4 结论与讨论
通过回归分析得出10种灌木各器官和全株生物量的最优模型。以植冠面积(A)、植冠体积(V)、基径与株高乘积(DH)、基径平方与株高乘积(D2H)等为自变量来估算生物量,克服用直接收获量法研究生物量的困难和麻烦,同时可以减少对野生植物资源的破坏,且具有较高的实用价值。同时,为研究大岗山林区林下灌木层的生态功能和准确测定林下灌木生物量提供科学依据,也为评价大岗山林区林下常见资源植物价值、评价大岗山不同森林类型生态服务功能价值提供关于生物量的基础数据。
灌木生物量研究表明,灌木各器官和全株生物量与DH、D2H和V呈显著相关,在最优模型中,灌木器官及全株最优模型方程形式多为二次或者三次方程,各灌木生物量最佳估算模型均有较高的R2值。全株生物量最优模型采用的自变量多为V;叶生物量最优模型采用的自变量多为V或A;枝、干生物量最优模型采用的自变量多为DH或者V,这与陈瑕林[8]等研究结果有所不同,其枝、干生物量模型所选自变量为D2H;根生物量最优模型采用的自变量多为D2H或V,这与以往研究存在不同程度的一致性[21-24]。部分研究显示灌木器官生物量模型拟合效果优于全株生物量模型[25],本研究结果表明,灌木器官模型不总是优于全株模型。灌木各器官和全株生物量最优模型估测精度存在差异,通过比较判定系数R2可以看出,单一物种干生物量最优模型估测精度最高,对大部分灌木而言,干、枝、叶生物量模型的估测精度高于根生物量模型。将单一物种器官生物量模型应用于大岗山林区常绿阔叶林、人工杉木林、天然毛竹林下这10种灌木的总生物量的计算,生物量从大到小依次为常绿阔叶林、人工杉木林、天然毛竹林,并且不同森林类型下各物种生物量情况存在较大差异。
本研究回归分析所得生物量模型均建立在大岗山林区林下灌木调查的基础上,对于灌木种植株高度、地径调查范围以外或研究区域以外的应用需通过进一步验证。
[1]卢振龙,龚孝生.灌木生物量测定的研究进展[J].林业调查规划,2009,34(4):37-40,45.
[2]Alamgir M,Al-Amin M.Allometric models to estimate biomass organic carbon stock in forest vegetation[J].Journal of Forestry Research,2008,19(2):101-106,171.
[3]黄怀雄,赵红艳.长株潭地区森林固碳释氧功能价值评价[J].林业调查规划,2010,35(2):136-138.
[4]国家林业局森林资源管理司.第七次全国森林资源清查及森林资源状况[J].林业资源管理,2010(1):1-8.
[5]谢咏红,彭珍宝,旷建军,等.基于能值理论的南岳衡山野生植物价值估算[J].现代农业科技,2010(4):11-13.
[6]Navar J,Mendez E,Najera A,et al.Biomass equations for shrub species of Tamaulipan thornscrub of North-eastern Mexico[J].Journal of Arid Environments,2004,59:657-674.
[7]曾慧卿,刘琪璟,冯宗炜,等.红壤丘陵区林下灌木生物量估算模型的建立及其应用[J].应用生态学报,2007,18(10):2185-2190.
[8]陈遐林,马钦彦,康峰峰,等.山西太岳山典型灌木林生物量及生产力研究[J].林业科学研究,2002,15(3):304-309.
[9]郑绍伟,唐敏,邹俊辉,等.灌木群落及生物量研究综述[J].成都大学学报:自然科学版,2007,26(3):189-192.
[10]范小莉.长白山地区云冷杉林和近原始林林下灌草生物量预测模型的研究[D].北京:北京林业大学,2011.
[11]程堂仁,马钦彦,冯仲科,等.甘肃小陇山森林生物量研究[J].北京林业大学学报,2007,29(1):31-36.
[12]曾慧卿,刘琪璟,马泽清,等.千烟洲灌木生物量模型研究[J].浙江林业科技,2006,26(1):13-17.
[13]Patona D,Nunez J,Bao D,et al.Forage biomass of 22 shrub species from Monfrage Natural Park(SW Spain)assessed by loglog regressin models[J].Journal of Ard Environments,2002,52(2):223-231.
[14]林伟,李俊生,郑博福,等.井冈山自然保护区12种常见灌木生物量的估测模型[J].武汉植物学研究,2010,28(6):725-729.
[15]王蕾,张宏,哈斯,等.基于冠幅直径和植株高度的灌木地上生物量估测方法研究[J].北京师范大学学报:自然科学版,2004,40(5):700-704.
[16]何列艳,亢新刚,范小莉,等.长白山区林下主要灌木生物量估算与分析[J].南京林业大学学报:自然科学版,2011,35(5):45-50.
[17]李燕,张建国,段爱国,等.杉木人工林生物量估算模型的选择[J].应用生态学报,2010,21(12):3036-3046.
[18]唐守正,张会儒,胥辉.相容性生物量模型的建立及其估计方法研究[J].林业科学,2000,36(增1):19-27.
[19]惠刚盈,罗云伍,张校林.江西大岗山丘陵区杉木人工林生产力的研究[J].林业科学,1989,25(6):564-569.
[20]刘大海,李宁,晁阳.SPSS15.0统计分析从入门到精通[M].北京:清华大学出版社,2008.
[21]魏文俊.江西省暨大岗山林区森林碳密度与碳储量的研究[D].呼和浩特:内蒙古农业大学,2007.
[22]段爱国.杉木人工林生物量变化规律的研究[J].林业科学研究,2005,18(2):125-132.
[23]张家城.江西分宜地区杉木人工林不同代数间生产力与生物量构成的比较研究[J].林业科学研究,2001,14(2):160-167.
[24]Sah P J,Ross S M,Koptur S,et al.Estimating aboveground biomass of broadleaved woody plants in the understory of florida keys pine forests[J].Forest Ecology and Management,2004,203:319-329.
[25]李晓娜,国庆喜,王兴昌,等.东北天然次生林下木树种生物量的相对生长[J].林业科学,2010,46(8):22-32.
猜你喜欢
天津农林科技(2022年2期)2022-04-19
农民致富之友(2020年8期)2020-05-11
乡村科技(2018年8期)2018-06-27
乡村科技(2018年8期)2018-02-13
现代园艺(2017年23期)2018-01-18
现代园艺(2017年23期)2018-01-18
诗潮(2017年2期)2017-03-16
湖南林业科技(2017年1期)2017-02-06
湖南林业科技(2017年1期)2017-02-06
广西林业科学(2016年2期)2016-03-20
