
紫花苜蓿耐盐性研究进展
2012-04-12 03:42:18张立全张凤英哈斯阿古拉
草业学报 2012年6期
张立全,张凤英,哈斯阿古拉
(内蒙古大学生命科学学院 内蒙古自治区牧草与特色作物生物技术重点实验室,内蒙古 呼和浩特010021)
*紫花苜蓿(Medicagosativa)是一种全球性栽培的豆科苜蓿属多年生草本植物,素有“牧草之王”和“饲料皇后”的美称[1]。然而,50~200 mmol/L NaCl胁迫就会显著降低紫花苜蓿的产量[2]。因此,选育耐盐紫花苜蓿品种既能提高盐碱地的利用,扩大紫花苜蓿种植面积,增加其产量,促进畜牧业的发展,又可以改良土壤,保持水土和保护生态环境。
近年来,国内外有关紫花苜蓿耐盐性的研究主要集中在紫花苜蓿耐盐品种的杂交选育、耐盐性生理、耐盐相关基因特性分析以及耐盐品种的基因工程选育。笔者围绕这几方面对紫花苜蓿的耐盐性研究进展进行综述,旨在为今后的研究工作提供参考。
1 耐盐紫花苜蓿杂交培育
20世纪,育种学家们在不完全了解作物表型和生理生化相互关系的情况下,通过传统的杂交育种方法获得了耐盐、高产、高品质和抗病的作物新品种。紫花苜蓿的耐盐育种在国外也有报道,如通过杂交培育获得的耐盐品 种 AZ-Germ SaltII[3]、AZ-90NDC-ST[4]、AZ-97MEC 和 AZ-97MEC-ST[5]、ZS-9491 和 ZS9592[6]以 及 Alfalafa[7]。Peel等[8]建立了温室中高效筛选耐盐紫花苜蓿的方法。我国的苜蓿育种工作起步较晚[9],但目前通过传统杂交育种已获得了9个耐盐性品种(表1)。
2 紫花苜蓿耐盐生理研究
2.1 耐盐生理研究
为了选育耐盐新品种,对紫花苜蓿耐盐性生理的研究十分必要,而且近年来已有许多相关研究报道。Torabi等[18]分析了不同气候条件下紫花苜蓿的耐盐性,发现不同品种紫花苜蓿因基因型不同而耐盐性不同,源于干旱荒漠地区的紫花苜蓿品种的耐盐性强于其他地区紫花苜蓿的耐盐性。Na2SO4和NaCl胁迫明显影响了紫花苜蓿的生长和离子吸收,相对耐盐性强的品系茎部Na+、Cl-和S2-的含量相对较低,且在以Na2SO4为主要盐分的土地上属于中度耐盐植物[19]。150 mmol/L NaCl长期胁迫紫花苜蓿,降低了叶片的大小,对老叶片引起的盐损伤比新叶片的严重,而且这种损伤与Na+/K+和清除H2O2酶的活性有关[20]。随着灌溉水中NaCl浓度(0~110 mmol/L)的升高,紫花苜蓿中Na和Cl的含量明显增高,而高浓度的Na和Cl将会影响苜蓿的品质,但在叶片中Ca和Mg的含量降低,在茎中K的含量增加,Zn和Fe在叶和茎中均降低,而这些微量金属离子含量的变化尚未影响到苜蓿的品质,仍适合动物的吸收[21]。盐胁迫既影响抗氧化酶的活性,又引起了脯氨酸的积累[22],盐胁迫条件下紫花苜蓿发芽期抗氧化酶活性与其耐盐性呈正相关[23],耐盐品种抗氧化酶活性高于敏盐品种[23,24],但紫花苜蓿苗期植株地上部Na+/K+与品种的耐盐性呈负相关[25]。
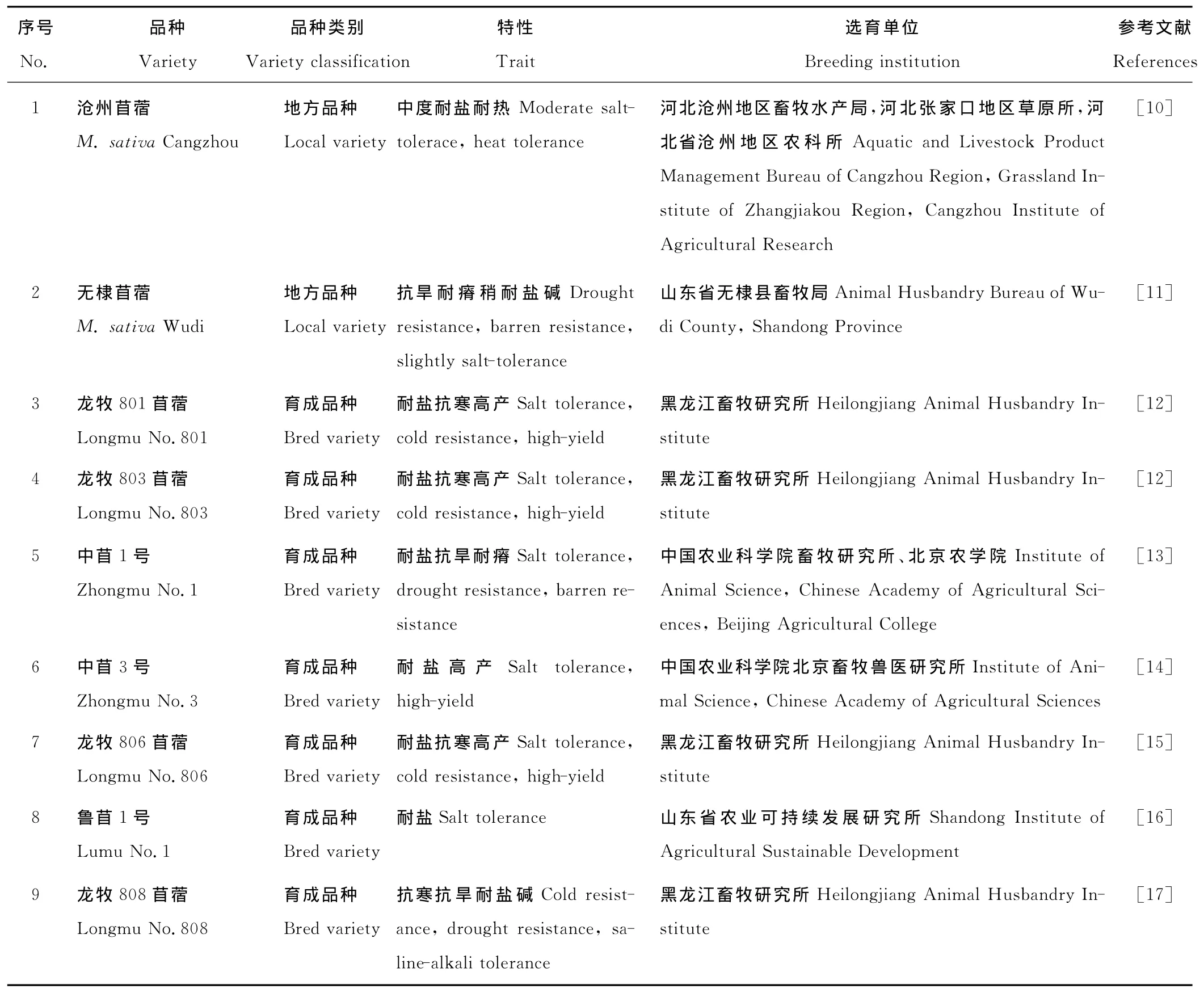
表1 耐盐紫花苜蓿品种Table 1 Cultivar of salt-tolerance alfalfa
低浓度的NaCl对紫花苜蓿种子的发芽和幼苗生长没有抑制作用或者具有促进作用,而外施ABA可以减弱高浓度NaCl对幼苗生长的抑制作用[26,27],用维生素C浸种处理可促进紫花苜蓿种子在盐胁迫下的萌发,增强根系和幼苗的生长[28],沙引发也可提高紫花苜蓿种子的活力和抗盐胁迫能力,促进盐逆境下种子的萌发和幼苗生长[29]。Wang等[30]研究发现H2S可能是通过与NO的相互作用,减少盐胁迫对植物氧化损伤,而缓解NaCl胁迫引起紫花苜蓿种子发芽和幼苗生长的抑制作用,增强紫花苜蓿对盐胁迫的应答耐性。近来,研究发现γ-射线的照射或臭氧协同胁迫时,也可提高紫花苜蓿的耐盐性。γ-射线照射下,紫花苜蓿中Na+积累降低[31];在臭氧协同胁迫时,盐土上紫花苜蓿相对产量增加42%[32]。
在自然界,盐胁迫和碱胁迫通常会同时发生,尤其在我国华北地区的土地盐分主要是中性盐(NaCl和Na2SO4)和碱性盐(Na2CO3和Na HCO3),而Na2CO3和Na HCO3含量约为总盐分的90%[33]。因此,了解混合盐碱对紫花苜蓿的影响亦十分必要。Peng等[34]用混合盐(NaCl、Na2SO4、Na HCO3和Na2CO3)和混合盐碱(盐浓度24~120 mmol/L,p H 7.03~10.32)处理紫花苜蓿,评析紫花苜蓿对混合盐碱适应的生理特性,发现高浓度的混合盐碱的胁迫损伤强于盐胁迫或碱胁迫。Gao等[35]分析了混合盐碱对紫花苜蓿种子发芽的影响,发现盐胁迫或碱胁迫引起紫花苜蓿胚根和胚芽长度的差异,而且混合盐碱胁迫还可以引起可溶性糖和Na+/K+的变化。张永峰等[36,37]分析了混合NaCl/Na2CO3胁迫对紫花苜蓿生理指标的影响,发现随着盐浓度的增大,可溶性糖、脯氨酸含量、抗氧化酶以及束缚水和自由水比率升高,而叶片持水力降低。
紫花苜蓿与根瘤菌共生固氮的生理特性对改良土壤肥力,提高作物产量有着十分重要的作用和意义,但在盐渍地盐胁迫下豆科牧草共生固氮体系效能降低。有关紫花苜蓿耐盐性和固氮能力的相互作用亦有研究报道。Aydi等[38]比较分析了盐胁迫下紫花苜蓿的固氮能力和离子平衡,发现盐胁迫降低了紫花苜蓿的根瘤效率,但相对耐盐的品种叶片Na+含量最低,且根瘤量最大。Mhadhbi和Aouani[39]认为氮的固定减弱了盐胁迫引起的氧化胁迫,而Salah等[40]提出苜蓿属植物的固氮耐盐作用是与结节糖代谢有关的新观点。
环境营养成分会明显的影响植物对盐环境的适应,盐胁迫会抑制磷(phosphorus,P)营养的吸收,且在低浓度P时,这种相互影响更加严重。Rogers等[41]分析了NaCl和P共同作用对紫花苜蓿的耐盐性和P吸收的影响,发现低浓度盐胁迫时,添加P可以促进茎的生长,但低浓度P营养引起的生长抑制作用比盐胁迫更严重。
2.2 耐盐筛选时期及指标研究
对于紫花苜蓿耐盐性筛选和鉴定的时期,AI-Khatib和Collins[42]认为在早期进行耐盐筛选最为合适,因为紫花苜蓿在发芽期和苗期对盐分比较敏感,Li等[43]分别分析了盐(NaCl∶Na2SO4=1∶1,p H 7.01~7.05)和碱(Na HCO3∶Na2CO3=1∶1,p H 9.80~10.11)对紫花苜蓿的影响,发现紫花苜蓿在发芽期和幼苗早期对盐和碱的胁迫均很敏感,但碱胁迫引起的危害较盐胁迫严重。Johnson等[44]发现发芽期的混合选择能够提高紫花苜蓿的耐盐性。McCoy[45]则认为紫花苜蓿在苗期进行耐盐筛选最易进行,于卓等[46]发现紫花苜蓿苗期对盐分反应较发芽期更为敏感。
紫花苜蓿耐盐性鉴定的分析指标目前仍有不同的观点,李国良等[47]选用地上生物量作为耐盐性鉴定指标,对适宜我国北方种植的苜蓿品种进行耐盐性鉴定。刘春华和张文淑[48]分析了69个苜蓿品种的耐盐性,提出叶片细胞膜透性和脯氨酸积累量不能作为耐盐评价的指标。Wang和Han[22]认为抗氧化酶活性可以作为紫花苜蓿发芽期耐盐性鉴定的分析指标。任卫波等[49]提出了应用近红外指纹光谱(near infrared reflectance spectroscopy,NIRS)快速鉴别紫花苜蓿品种耐盐性的新方法,并通过对20个紫花苜蓿品种进行聚类分析,建立了紫花苜蓿品种耐盐性鉴定模型。李源等[50]提出运用标准差系数赋予权重法进行综合评价,不但考虑了不同指标的权重,还定量的鉴定出了每份材料的耐盐能力,比聚类分析的结果更具科学合理性。在此基础上,探讨了盐胁迫下紫花苜蓿的生理反应,认为可溶性糖含量、细胞质膜透性、丙二醛(malondialdehyde,MDA)含量、水分饱和亏缺以及叶水势等可直接作为耐盐评价的鉴定指标。姜健等[51]利用随机扩增多态性DNA(random amplified polymorphic DNA,RAPD)技术分析紫花苜蓿耐盐种质的遗传多样性,为紫花苜蓿耐盐种质核心种质库构建和耐盐新品种选育提供了理论依据。
3 紫花苜蓿耐盐基因工程研究
3.1 紫花苜蓿耐盐相关基因
近年来,许多参与调节离子平衡、渗透调节、抗氧化酶、转录因子等方面的盐诱导相关基因已从紫花苜蓿中获得克隆,而且这些基因与耐盐相关的功能特性也得到充分确认。
3.1.1 离子平衡相关基因 为了适应盐胁迫,植物进化获得了一系列的适应机制,其中之一是将Na+区隔在叶泡中,来减少胞质中过量Na+引起的损伤,同时将Na+作为有效的渗透调节剂来维持渗透平衡,从而增强植物的耐盐性和吸水能力。植物中Na+在叶泡中的区隔化是由Na+/H+反向转运体介导的。紫花苜蓿Na+/H+反向转运体基因Ms NH X1的表达受高浓度NaCl诱导,且MsN HX1具有转运Na+的功能,转Ms NH X1拟南芥(Arabidopsisthaliana)的耐盐性增强[52]。Na+/H+反向转运体介导Na+在叶泡中的区隔化是由液泡 H+-泵、H+-ATPase和 H+-焦磷酸酶(H+-pyrophosphatase,H+-PPase)产生 H+梯度驱动的。Sibole等[53]分析了木本苜蓿(Medicagoarborea)质膜H+-ATPase在应答盐胁迫时的作用,发现盐胁迫时木本苜蓿H+-ATPase活性增强,能够选择性的清除叶片中的Na+,维持叶片的生长,因此,H+-ATPase的活性增加可能与适应盐胁迫而清除Na+有关。
3.1.2 渗透调节相关基因 在逆境胁迫时,脯氨酸是被公认的渗透调节剂。脯氨酸合成途径中的关键酶△1-二氢毗咯-5-羧酸合成酶[Delta(1)-pyrroline-5-carboxylate synthetase]基因P5CS是目前研究比较多的渗透调节基因。Ginzberg等[54]从盐胁迫紫花苜蓿根部cDNA文库中分离获得了2个编码P5CS的c DNA,即MsP5CS-1和MsP5CS-2,二者的表达均受盐胁迫诱导。为了进一步了解盐胁迫对脯氨酸合成和降解的影响,Miller等[55]克隆了紫花苜蓿脯氨酸脱氢酶(proline dehydrogenase)基因MsPDH和△1-二氢毗咯-5-羧酸脱氢酶[Delta(1)-pyrroline-5-carboxylate dehydrogena-se]基因MsP5CDH,表达分析显示盐胁迫引起的脯氨酸积累和MsPDH基因的转录水平降低呈正相关,而盐胁迫没有影响基因MsP5CDH和MsP5CS的表达。Deutch和Winicov[56]从耐盐紫花苜蓿根特异性的cDNA文库中筛选获得了一个cDNA,被命名为MsPRP2,编码富含脯氨酸的细胞壁蛋白,其表达受盐胁迫的诱导。
Nolan等[57]从蒺藜苜蓿中克隆获得了胁迫激酶基因MtSK1,体外实验显示MtSK1的表达受盐胁迫的诱导。LEA(late embryogenesis abundant)蛋白是在植物胚胎发育晚期产生的,约占细胞总蛋白的4%,LEA基因的表达受渗透胁迫的调节。紫花苜蓿MsLEA3-1基因的表达受NaCl和ABA的诱导,超表达Ms LEA3-1的转基因烟草(Nicotianabenthamiana)的耐盐性明显增强[58],表明 Ms LEA3-1蛋白具有盐胁迫保护功能,因此Ms-LEA3-1基因具有耐盐基因工程候选基因的潜能。Jin等[59]以中苜1号为材料,利用消减杂交(suppression subtractive hybridization,SSH)技术构建了紫花苜蓿盐胁迫表达cDNA文库,获得了LEA蛋白基因。
3.1.3 转录因子相关基因 Winicov[60]首次从筛选获得的耐盐紫花苜蓿细胞系中克隆到了转录因子基因Alfin1,其编码锌指蛋白,可以与紫花苜蓿盐诱导表达基因MsPRP2启动子结合,调节MsPRP2的表达[61]。蒺藜苜蓿Krüppel类锌指蛋白转录因子Mt ZPT2-1能够提高酵母细胞的耐盐性,表达反向MtZPT2-1基因的紫花苜蓿失去具有固氮能力的结节[62]。Merchan等[63]发现Mt ZPT2-1基因主要在根和根部结节表达,且受盐胁迫的诱导。表达反向Mt ZPT2-1基因的紫花苜蓿恢复盐胁迫损伤的敏感性比未转基因紫花苜蓿更强,因此Mt ZPT2-1基因可能会被作为一个有效的分子标记,应用在紫花苜蓿耐盐性育种的耐盐性筛选中。紫花苜蓿Cab9基因是一个AP2蛋白家族转录因子基因[64]。Chao等[65]从紫花苜蓿c DNA文库中克隆获得了紫花苜蓿锌指蛋白基因MsZFN,其表达明显受NaCl的诱导。
3.1.4 抗氧化相关基因 Kang等[64]利用SSH技术构建了蒺藜苜蓿盐胁迫表达cDNA文库,获得了35个功能已知的基因表达序列标签(expressed sequence tags,ESTs),其中超氧化物歧化酶(superoxide dismutase,SOD)-1,CuZn-SOD和半胱氨酸蛋白酶基因受盐胁迫诱导表达。Rubio等[66]分析了在结节和叶片线粒体中表达 Mn-SOD、在叶绿体中表达MnSOD和FeSOD的转基因紫花苜蓿的SOD活性,发现紫花苜蓿的SOD在组织器官中的活性受转录后水平调控,且FeSOD具有重要的抗氧化功能。
血红素加氧酶(haem oxygenases,HO)催化血红素氧化降解,具有抗氧化保护的作用。紫花苜蓿HO1基因在成熟根结节大量表达,但其表达不受氧化剂H2O2、除草剂和硝酸钠的诱导,表明紫花苜蓿HO1不具有抗氧化保护的作用[67]。韩毅和沈文飚[68]发现HO的活性诱导剂高铁血红素对汞诱导紫花苜蓿根部组织的氧化损伤具有保护作用,而Fu等[69]的研究发现在氧化剂H2O2和硝酸钠处理时,紫花苜蓿HO2基因的表达被上调,说明HO2可能具有抗氧化保护的作用。
3.1.5 钙相关蛋白基因 植物钙依赖蛋白激酶(calcium-dependent protein kinases,CDPKs)参与各种信号途径,MtCPK3基因是蒺藜苜蓿根部CDPK异构体,其在盐胁迫早期出现表达[70]。紫花苜蓿钙相关蛋白(calmodulin-related protein)PPRG1具有传导逆境胁迫信号的功能,PPRG1的表达在盐、渗透、低温以及ABA的胁迫下快速上调[71]。
3.1.6 其他功能相关基因 Winicov和Button[72]分析了盐胁迫对紫花苜蓿光合作用相关基因表达的影响,结果显示光合作用相关叶绿体基因psbD、psaB、atp B、rbc L以及核基因pCab4、p Cabl、rbcS在耐盐紫花苜蓿中的表达增强。解旋酶在植物中参与适应盐和低温的胁迫,紫花苜蓿解旋酶1基因(M.sativahelicase 1,M H1)在根、茎和叶片中均有表达,且MH1的表达受NaCl、ABA和甘露醇的诱导。由35S启动子驱动在拟南芥中组成型表达M H1,转基因植株的耐盐性和抗旱性明显增强,抗氧化能力以及参与渗透调节的脯氨酸含量均升高,表明M H1基因通过增强清除活性氧(reactive oxygen species,ROS)和渗透调节的能力来增强耐盐和抗干旱的能力[73]。在真核生物模式植物烟草中表达紫花苜蓿线粒体热激蛋白(heat shock protein)基因Ms HSP23,转基因植株的耐盐性明显高于对照植株[74]。
3.2 耐盐紫花苜蓿基因工程培育
1985年,Vasil在国际草原学大会上第一次报告了利用遗传转化技术将特定基因导入牧草的可行性,为基因工程技术改良牧草,增加牧草对逆境的适应能力奠定了理论基础。1986年,Deak等[75]首次报道利用转基因技术获得抗性苜蓿植株。此后,利用基因工程技术提高苜蓿抗逆性已成为苜蓿育种的重要途径。目前,已有许多利用转耐盐相关基因来改良紫花苜蓿耐盐性的研究报道[76,77]。
盐胁迫时,在蒺藜苜蓿中表达绿藻(Anabaenavariabilis)黄素氧还蛋白基因flavodoxin,可引起转基因植株结节正调控固氮作用的氧化还原平衡酶活性变化,使得转基因植株的固氮能力与对照植株之间存在显著差异[78]。因此,flavodoxin基因具有改善豆科牧草耐盐和固氮特性的潜能。Verdoy等[77]在蒺藜苜蓿中表达P5CS基因,首次获得了通过增强耐盐性而提高固氮能力的转基因苜蓿。Suárez等[79]将酵母海藻糖-6-磷酸合成酶(trehalose-6-phosphate synthase,TPS)基因TPS1和海藻糖-6-磷酸磷酸酶(trehalose-6-phosphate phosphatase,TPP)基因TPS2融合,分别由35S和rd29A启动子驱动在紫花苜蓿中表达,转基因株系的耐盐性显著增强,这是首次将海藻糖代谢基因应用于紫花苜蓿基因工程中,且提高了紫花苜蓿对逆境胁迫的保护能力。
Li等[80]在紫花苜蓿中高效表达盐生植物苏打猪毛菜(Salsolasoda)Na+/H+反向转运体基因SsN HX1,转基因植株的耐盐性显著增强,可以在400 mmol/L NaCl胁迫条件下生长50 d。Jin等[81]在紫花苜蓿中诱导表达了Gm DREB1基因,Nothern杂交表明转基因植株的P5CS基因表达被上调,且转基因紫花苜蓿的耐盐性比对照植株显著提高。Bao等[76]将拟南芥H+-PPase基因AVP1导入紫花苜蓿新疆大叶苜蓿中,分析超表达AVP1的转基因植株的耐盐性,结果表明转AVP1可以明显地提高紫花苜蓿的耐盐性。Winicov和Bastola[82]将转录因子Aflin1导入紫花苜蓿,超表达Aflin1的愈伤组织可抵抗171 mmol/L NaCl胁迫作用,同时增强了MsPRP2的表达,且转基因紫花苜蓿的耐盐性增强[83]。刘艳芝等[84]将酵母HAL1基因转入紫花苜蓿龙牧803,获得了11株在含0.6%~1.0%NaCl的MS培养基上正常生长的转基因植株。王瑛等[85]将大麦(Hordeumvulgare)lea3基因转入紫花苜蓿中苜1号,与对照植株相比,转基因苜蓿耐盐性显著增强。化烨[86]将东北野生大豆(Glycinesoja)S-硫腺苷甲硫氨酸合成酶(S-Adenosy-L-Methion-nine,SAMS)基因GsSAMS导入肇东苜蓿,培育出耐盐碱的转基因苜蓿新株(品)系。王玉祥等[87,88]分析了表达费氏中华根瘤菌(Sinorhizobiumfredii)糖基/苷转移酶基因rst B保定苜蓿转基因幼苗的耐盐性,盐胁迫对幼苗的发芽率和根系影响的结果显示rst B基因可以显著提高紫花苜蓿的耐盐性,而且转基因植株的叶绿素和甜菜碱的含量呈增长趋势,而细胞膜相对透性呈下降趋势,表明转基因植株通过这些生理机制变化适应盐胁迫,减轻盐损伤,使其耐盐性优于对照植株。
山东省林业科学研究院课题组采用农杆菌介导技术将山菠菜(Atriplexhortensis)甜菜碱醛脱氢酶基因Ah-BADH导入紫花苜蓿受体材料中苜1号中,获得了PCR阳性植株。耐盐实验结果表明,T1代转基因植株可耐受0.8%和0.9%NaCl的胁迫,并正常生长,且BADH基因在T2代转基因植株中符合孟德尔分离法则,耐盐性明显高于未转基因受体植株,经过对T3代转基因植株耐盐性鉴定和遗传分离筛选,通过自交和杂交选育出遗传稳定、综合性状优良且耐盐碱强的转基因苜蓿新品种山苜2号[89-92]。Liu等[93]将Ah BADH导入紫花苜蓿Sanditi品种中,转基因植株的耐盐性明显增强。
张立全等[94,95]利用花粉管通道法将盐生植物红树总DNA导入紫花苜蓿阿尔岗金品种,获得12株耐盐性强的T0代植株。RAPD分析初步证实外源DNA已经整合到受体的基因组中,而且T0代植株耐盐能力提高可能与外源基因的导入有关。T1代幼苗经300 mmol/L NaCl胁迫后测定耐盐相关生理生化指标,结果表明T1代植株耐盐性明显增强,并获得了T1代耐盐株系,为进一步培育耐盐品种提供了新的种质资源。
4 结束语
紫花苜蓿作为重要的优良豆科牧草,对畜牧业的发展以及生态环境的改善起着重要作用,但中度耐盐的能力限制了紫花苜蓿的种植范围,因此选育耐盐性强的紫花苜蓿新品种是育种学家们急需解决的问题。育种学家们选用传统的杂交育种或基因工程技术进行了大量的新品种选育工作,虽然利用传统杂交技术已经获得了许多耐盐品种,但由于选育周期性长、费时耗力等缺点限制了传统杂交育种的发展和利用。随着分子生物学的发展,基因工程技术便成为全世界育种学家们热衷的育种方法[96,97]。目前,大量的研究报道利用基因工程技术已获得了耐盐性紫花苜蓿新品系(种),但除山苜2号品种外,其他的研究报道结果仍是基于人工模拟的环境——实验室或温室中的NaCl胁迫的研究特性,缺少相应的品种区域特性试验研究,而区域试验是评价新品种适应性最直接可靠的办法[98]。另外,由于植物的耐盐性是一个受多基因控制的数量性状[99,100],而目前的研究报道只涉及单基因的转化,这可能是限制基因工程耐盐育种发展的原因之一。再者,目前的研究报道均是在转基因品系(种)的生长早期——发芽期和(或)幼苗期进行的耐盐性评析,而其耐盐性是否可以维持并稳定遗传尚有待证实,且某个发育阶段的耐盐性尚不能代表植物的耐盐性[101]。
基因工程技术育种过程中选择标记基因的使用引起了人们对生物安全的顾虑,Ferradini等[102]分析了无筛选标记转化(marker-less transformation)和共转化(co-transformation)方法获得无选择标记基因(marker-free)紫花苜蓿的效率,发现这2种方法虽然能获得无选择标记基因的转基因紫花苜蓿,但转化效率非常低,在实际应用中仍需要进一步完善。因此,目前高效的紫花苜蓿转化中选择标记基因仍是必需的。但是为了避免源于细菌的选择标记基因在植物基因工程中带来抗生素抗性的危险,Ferradini等[103]克隆了源于紫花苜蓿的谷氨酸1-半醛转氨酶(glutamate 1-semialdehyde aminotransferase)基因MsGSA,并对MsGSA进行单一位点突变修饰,通过转化烟草和紫花苜蓿分析其选择效率,开发获得了可用于紫花苜蓿遗传转化的高效选择标记基因。
紫花苜蓿的耐盐性是个较复杂的问题,它是耐盐相关代谢途径以及其他多种代谢途径的协同综合作用的结果。目前,人们对紫花苜蓿的耐盐相关代谢途径仍不完全清楚,这正是限制获得耐盐性强的新品种的主要障碍。因此,全面研究并了解紫花苜蓿的耐盐相关代谢途径以及耐盐相关基因和其表达模式是紫花苜蓿耐盐分子育种的前提条件。要获得耐盐性强的品种,还需要全面地研究和了解紫花苜蓿的耐盐机制,相信在此基础上,必可培育出具有实际应用价值的耐盐新品种,为畜牧业的发展做出贡献。
[1] 曹宏,章会玲,盖琼辉,等.22个紫花苜蓿品种的引种试验和生产性能综合评价[J].草业学报,2011,20(6):219-229.
[2] Bekki A,Trinchant J C,Rigaud J.Nitrogen fixation(C2H2reduction)byMedicagonodules and bacteroids under sodium chloride stress[J].Physiologia Plantarum,1987,71:61-67.
[3] Dobrenz A K,Robinson D L,Smith S E,etal.Registration of AZ-germ salt-II nondormant alfalfa germplasm[J].Crop Science,1989,29:493.
[4] Johnson D W,Smith S E,Dobrenz A K.Registration of AZ-90NDC-ST nondormant alfalfa germplasm with improved forage yield in saline environments[J].Crop Science,1991,31:1098-1099.
[5] Al-Doss A A,Smith S E.Registration of AZ-97 MEC and AZ-97 MEC-ST very non-dormant alfalfa germplasm pools with increased shoot weight and differential response to saline irrigation[J].Crop Science,1998,38:568.
[6] Dobrenz A K.Salt-tolerant alfalfa[P].United States Patent:6005165,1999-10-21.
[7] Downes R W.New herbage CultivarsMedicagosativaCV.Alfalfa[J].Tropical Grasslands,1994,28:191-192.
[8] Peel M D,Waldron B L,Jensen K B,etal.Screening for salinity tolerance in alfalfa:A repeatable method[J].Crop Science,2004,44(6):2049-2053.
[9] 赵桂琴,慕平,张勃.紫花苜蓿基因工程研究进展[J].草业学报,2006,15(6):9-18.
[10] 张玉成,刘凤泉,刘肇清,等.沧州苜蓿的品种特性及利用[J].草与畜杂志,1992,3:5-7.
[11] 张令进,朱树森.无棣紫花苜蓿[J].农业知识,1997,9:23.
[12] 李红,罗新义,王殿魁.“龙牧801”和“龙牧803号”苜蓿新品种选育报告[J].黑龙江畜牧科技,1996,1:3-7.
[13] 杨青川,耿华珠,孙彦.耐盐苜蓿新品种中苜一号[J].作物品种资源,1999,2:26.
[14] 2006年度全国草品种审定委员会审定通过的草品种名录(一)[J].草业科学,2007,24(5):99.
[15] 李红.高产、高蛋白、高抗性龙牧806号苜蓿[J].牧草与饲料,2007,1(3):64.
[16] 贾春林,杨秋玲,吴波,等.鲁苜1号紫花苜蓿选育及栽培技术[J].山东农业科学,2008,5:100-103.
[17] 李红,杨曌,黄新育,等.龙牧808紫花苜蓿新品种选育报告[J].中国草地学报,2011,33(3):12-17.
[18] Torabi M,Halim R A,Sinniah U R,etal.Influence of salinity on the germination of Iranian alfalfa ecotypes[J].African Jouranl of Agricultural Reserch,2011,6(19):4624-4630.
[19] Rogers M E,Grieve C M,Shannon M C.The response of lucerne(MedicagosativaL.)to sodium sulphate and chloride salinity[J].Plant and Soil,1998,202:271-280.
[20] Boughanm N,Michonneau P,Daghfous D,etal.Adaptation ofMedicagosativacv.Gabes to long-term NaCl stress[J].Journal of Plant Nutrition Soil Science,2005,168:262-268.
[21] Guerrero-Rodríguez J D,Revell D K,Bellotti W D.Mineral composition of lucerne(Medicagosativa)and white melilot(Melilotusalbus)is affected by NaCl salinity of the irrigation water[J].Animal Feed Science and Technology,2011,170:97-104.
[22] Wang X S,Han J G.Changes of proline content,activity,and active isoforms of antioxidative enzymes in two Alfalfa cultivars under salt stress[J].Agricultural Sciences in China,2009,8(4):431-440.
[23] Wang W B,Kim Y H,Lee H S,etal.Analysis of antioxidant enzyme activity during germination of alfalfa under salt and drought stresses[J].Plant Physiology and Biochemistry,2009,47:570-577.
[24] Babakhani B,Khavari-Nejad R A,Sajedi R H,etal.Biochemical responses of Alfalfa(MedicagosativaL.)cultivars subjected to NaCl salinity stress[J].African Journal of Biotechnology,2011,10(55):11433-11441.
[25] Bao A K,Guo Z G,Zhang H F,etal.A procedure for assessing the salt tolerance of lucerne(MedicagosativaL.)cultivar seedlings by combining agronomic and physiological indicators[J].New Zealand Journal of Agricultural Research,2009,52(4):435-442.
[26] 景艳霞,袁庆华.NaCl胁迫对苜蓿幼苗生长及不同器官中盐离子分布的影响[J].草业学报,2011,20(2):134-139.
[27] Wu C H,Wang Q Z,Xie B,etal.Effects of drought and salt stress on seed germination of three leguminous species[J].African Journal of Biotechnology,2011,10(78):17954-17961.
[28] 申玉华,徐振军,李文辉,等.维生素C浸种对盐胁迫下紫花苜蓿种子发芽特性的影响[J].种子,2009,28(7):42-44.
[29] 解秀娟,胡晋.沙引发对紫花苜蓿种子盐逆境下发芽及幼苗生理生化变化的影响[J].种子,2003,4:5-6.
[30] Wang Y Q,Li L,Cui W T,etal.Hydrogen sulfide enhances alfalfa(Medicagosativa)tolerance against salinity during seed germination by nitric oxide pathway[J].Plant and Soil,2011,351:107-119.
[31] Rejili M,Telahigue D,Lachiheb B,etal.Impact of gamma radiation and salinity on growth and K+/Na+balance in two populations ofMedicagosativa(L.)cultivar Gabes[J].Progress in Natural Science,2008,18:1095-1105.
[32] Maggio A,Chiaranda F Q,Cefariello R,etal.Responses to ozone pollution of alfalfa exposed to increasing salinity levels[J].Environmental Pollution,2009,157:1445-1452.
[33] 葛莹,李建东.盐生植被在土壤积盐—脱盐过程中作用的初探[J].草业学报,1990,1(1):70-76.
[34] Peng Y L,Gao Z W,Gao Y,etal.Eco-physiological characteristics of Alfalfa seedlings in response to various mixed salt-alkaline stresses[J].Journal of Integrative Plant Biology,2008,50(1):29-39.
[35] Gao Z W,Zhu H,Gao J C,etal.Germination responses of Alfalfa(MedicagosativaL.)seeds to various salt-alkaline mixed stress[J].African Jouranl of Agricultural Reserch,2011,6(16):3793-3803.
[36] 张永峰,梁正伟,隋丽,等.盐碱胁迫对苗期紫花苜蓿生理特性的影响[J].草业学报,2009,18(4):230-235.
[37] 张永峰,殷波.混合盐碱胁迫对苗期紫花苜蓿抗氧化酶活性及丙二醛含量的影响[J].草业学报,2009,18(1):46-50.
[38] Aydi S,Sassi S,Abdelly C.Growth,nitrogen fixation and ion distribution inMedicagotruncatulasubjected to salt stress[J].Plant and Soil,2008,32:59-67.
[39] Mhadhbi H,Aouani M E.Growth and nitrogen-fixing performances ofMedicagotruncatula-Sinorhizobiummelilotisymbioses under salt(NaCl)stress:Micro-and macro-symbiont contribution into symbiosis tolerance[J].Biosaline Agriculture and High Salinity Tolerance,2008,1:91-98.
[40] Salah I B,Slatni T,Albacete A,etal.Salt tolerance of nitrogen fixation inMedicagociliarisis related to nodule sucrose metabolism performance rather than antioxidant system[J].Symbiosis,2010,51:187-195.
[41] Rogers M E,Grieve C M,Shannon M C.Plant growth and ion relations in lucerne(MedicagosativaL.)in response to the combined effects of NaCl and P[J].Plant and Soil,2003,253:187-194.
[42] AI-Khatib-M M,Collins J C.Between and with in culture variability in salt tolerance in Lucerne[J].Genetic Resources and Crop Evoluation,1994,41(3):159-164.
[43] Li R L,Shi C,Fukuda K,etal.Effects of salt and alkali stresses on germination,growth,photosynthesis and ion accumulation in alfalfa(MedicagosativaL.)[J].Soil Science and Plant Nutrition,2010,56:725-733.
[44] Johnson D W,Smith S E,Dobrenz A K.Genetic and phenotypic relationship in response to NaCl at different developmental stages in alfalfa[J].Theoretical and Applied Genetics,1992,83:833-838.
[45] McCoy T J.Tissue culture evaluation of NaCl tolerance inMedicagospecies[J].Plant Cell Reports,1987,8(3):31-34.
[46] 于卓,孙祥,张文忠,等.苜蓿品种间种子萌发及苗期耐盐性差异的研究[J].干旱区资源与环境,1993,7(2):106-111.
[47] 李国良,刘香萍,迟文峰,等.紫花苜蓿耐盐性生理的初步研究[J].现代畜牧兽医,2007,3:26-27.
[48] 刘春华,张文淑.六十九个苜蓿品种耐盐性及其二个耐盐生理指标的研究[J].草业科学,1993,10(6):16-22.
[49] 任卫波,韩建国,张蕴薇,等.红外光谱紫花苜蓿品种耐盐性鉴别方法研究[J].光谱学与光谱分析,2009,29(2):386-388.
[50] 李源,刘贵波,高洪文,等.紫花苜蓿种质耐盐性综合评价及盐胁迫下的生理反应[J].草业学报,2010,19(4):79-86.
[51] 姜健,杨宝灵,夏彤,等.紫花苜蓿耐盐种质资源的遗传多样性分析[J].草业学报,2011,20(5):119-125.
[52] 安宝燕,罗琰,李加瑞,等.紫花苜蓿Na+/H+逆向转运蛋白基因在拟南芥中表达提高转基因植株的耐盐性[J].作物学报,2008,34(4):557-564.
[53] Sibole J V,Cabot C,Michalke W,etal.Relationship between expression of the PM H+-ATPase,growth and ion partitioning in the leaves of salt-treatedMedicagospecies[J].Planta,2005,221:557-566.
[54] Ginzberg I,Stein H,Kapulnik Y,etal.Isolation and characterization of two different cDNAs of Delta(1)-pyrroline-5-carboxylate synthase in alfalfa,transcriptionally induced upon salt stress[J].Plant Molecular Biology,1998,38:755-764.
[55] Miller G,Stein H,Honig A,etal.Responsive modes ofMedicagosativaproline dehydrogenase genes during salt stress and recovery dictate free proline accumulation[J].Planta,2005,222(1):70-79.
[56] Deutch C E,Winicov I.Post-transcriptional regulation of a salt-inducible alfalfa gene encoding a putative chimeric proline-rich cell wall protein[J].Plant Molecular Biology,1995,27:411-418.
[57] Nolan K E,Saeed N A,Rose R J.The stress kinase geneMtSK1 inMedicagotruncatulawith particular reference to somatic embryogenesis[J].Plant Cell Reports,2006,25:711-722.
[58] Bai Y Q,Yang Q C,Kang J M,etal.Isolation and functional characterization of aMedicagosativaL.gene,MsLEA3-1[J].Molecular Biology Reports,2012,39:2883-2892.
[59] Jin H C,Sun Y,Yang Q C,etal.Screening of genes induced by salt stress from Alfalfa[J].Molecular Biology Reports,2010,37:745-753.
[60] Winicov I.cDNA encoding putative zinc finger motifs from salt-tolerant alfalfa(MedicagosativaL.)cells[J].Plant Physiology,1993,102:681-682.
[61] Bastola D R,Pethe V V,Winicov I.Alfin1,a novel zinc-finger protein in alfalfa roots that binds to promoter elements in the salt-inducibleMsPRP2 gene[J].Plant Molecular Biology,1998,38:1123-1135.
[62] Frugier F,Poirier S,Satiat-Jeunematre B,etal.A Krüppel-like zinc finger protein is involved in nitrogen-fixing root nodule organogenesis[J].Genes Development,2000,14:475-482.
[63] Merchan F,Breda C,Hormaeche J,etal.A Krüppel-like transcription factor gene is involved in salt stress responses inMedicagospp.[J].Plant and Soil,2003,257:1-9.
[64] Kang J M,Xie W W,Sun Y,etal.Identification of genes induced by salt stress fromMedicagotruncatulaL.seedlings[J].African Journal of Biotechnology,2010,45(9):7589-7594.
[65] Chao Y H,Kang J M,Sun Y,etal.Molecular cloning and characterization of a novel gene encoding zinc finger protein fromMedicagosativaL.[J].Molecular Biology Reports,2009,36:2315-2321.
[66] Rubio M C,Ramos J,Webb K J,etal.Expression studies of superoxide dismutases in nodules and leaves of transgenic alfalfa reveal abundance of iron-containing isozymes,posttranslational regulation,and compensation of isozyme activities[J].Molecular Plant-Microbe Interactions,2001,14(10):1178-1188.
[67] Baudouin E,Frendo P,Le G M,etal.AMedicagosativahaem oxygenase gene is preferentially expressed in root nodules[J].Journal of Experimental Botany,2004,55:43-47.
[68] 韩毅,沈文飚.血红素加氧酶/一氧化碳信号系统对汞、镉诱导的紫花苜蓿根部氧化胁迫的调节[D].南京:南京农业大学,2008.
[69] Fu G Q,Jin Q J,Lin Y T,etal.Cloning and characterization of a heme oxygenase-2 gene from alfalfa(MedicagosativaL.)[J].Applied Biochemistry Biotechnology,2011,165:1253-1263.
[70] Gargantini P R,Gonzalez-Rizzo S,Chinchilla D,etal.A CDPK isoform participates in the regulation of nodule number inMedicagotruncatula[J].Plant Journal,2006,48(6):843-856.
[71] Borsics T,Lados M.cDNA cloning of a mechanical/abiotic stress-inducible calmodulin-related gene from dodder-infected alfalfa[J].Plant Cell and Environment,2001,(24):649-656.
[72] Winicov I,Button J D.Accumulation of photosynthesis gene transcripts in response to sodium chloride by salt-tolerant alfalfa cells[J].Planta,1991,183:478-483.
[73] Luo Y,Liu Y B,Dong Y X,etal.Expression of a putative alfalfa helicase increases tolerance toabiotic stress inArabidopsisby enhancing the capacities for ROS scavenging and osmotic adjustment[J].Journal of Plant Physiology,2009,166:385-394.
[74] Lee K W,Cha J Y,Kim K H,etal.Overexpression of alfalfa mitochondrialHSP23 in prokaryotic and eukaryotic model systems confers enhanced tolerance to salinity and arsenic stress[J].Biotechnology Letters,2012,34(1):167-174.
[75] Deak M,Kiss G B,Korkz C,etal.Transformation ofMedicagoby agrobacterum mediated gene transfer[J].Plant Cell Reports,1986,5:97-100.
[76] Bao A K,Wang S M,Wu G Q,etal.Overexpression of theArabidopsisH+-PPase enhanced resistance to salt and drought stress in transgenic alfalfa(MedicagosativaL.)[J].Plant Science,2009,176:232-240.
[77] Verdoy D,Coba P T,Redondo F J,etal.TransgenicMedicagotruncatulaplants that accumulate proline display nitrogenfixing activity with enhanced tolerance to osmotic stress[J].Plant Cell Environment,2006,29:1913-1923.
[78] Coba de la Pea T C,Francisco J R,Esteban M,etal.Nitrogen fixation persists under conditions of salt stress in transgenicMedicagotruncatulaplants expressing a cyanobacterial flavodoxin[J].Plant Biotechnology Journal,2010,8:954-965.
[79] Suárez R,Calderón C,Iturriaga G.Enhanced tolerance to multiple abiotic stresses in transgenic alfalfa accumulating trehalose[J].Crop Sciences,2009,49(5):1791-1799.
[80] Li W F,Wang D L,Jin T C,etal.The vacuolar Na+/H+antiporter geneSsN H X1 from the halophyteSalsolasodaconfers salt tolerance in transgenic Alfalfa(MedicagosativaL.)[J].Plant Molecular Biology Reporter,2011,29:278-290.
[81] Jin T C,Chang Q,Li W F,etal.Stress-inducible expression ofGm DREB1 conferred salt tolerance in transgenic alfalfa[J].Plant Cell,Tissue and Organ Culture,2010,100:219-227.
[82] Winicov I,Bastola D R.Transgenic overexpression of the transcription factorAlfin1 enhances expression of the endogenousMsPRP2 gene in Alfalfa and improves salinity tolerance of the plants[J].Plant Physiology,1999,120:473-480.
[83] Winicov I.Alfinl transcription factor overexpression enhances plant root growth under normal and saline conditions and improves salt tolerance in alfalfa[J].Planta,2000,210(3):416-422.
[84] 刘艳芝,韦正乙,邢少辰,等.HAL1基因转化苜蓿再生植株及其耐盐性[J].吉林农业科学,2008,33(6):21-24.
[85] 王瑛,朱宝成,孙毅,等.外源lea3基因转化紫花苜蓿的研究[J].核农学报,2007,21(3):249-252.
[86] 化烨.GsSAMS基因对苜蓿的遗传转化及转基因新株系的培育[D].哈尔滨:东北农业大学,2009.
[87] 王玉祥,王涛,张博.转rstB基因苜蓿耐盐性初评[J].草地学报,2008,16(5):539-541.
[88] 王玉祥,张博,王涛.盐胁迫对苜蓿叶绿素、甜菜碱含量和细胞膜透性的影响[J].草地科学,2009,26(3):53-55.
[89] 梁慧敏,夏阳,孙仲序,等.根癌农杆菌介导苜蓿遗传转化体系的建立[J].农业生物技术学报,2005,13:152-156.
[90] 燕丽萍,夏阳,梁慧敏,等.转BADH基因苜蓿T1代遗传稳定性和抗盐性研究[J].草业学报,2009,18(6):65-71.
[91] 燕丽萍,夏阳,毛秀红,等.转BADH基因紫花苜蓿山苜2号品种的抗盐性鉴定及系统选育[J].植物学报,2011,46(3):293-301.
[92] Yan L P,Liu C L,Liang H M,etal.Physiological responses to salt stress of T2alfalfa progenies carrying a transgene for betaine aldehyde dehydrogenase[J].Plant Cell,Tissue and Organ Culture,2012,108:191-199.
[93] Liu Z H,Zhang H M,Li G L,etal.Enhancement of salt tolerance in alfalfa transformed with the gene encoding for betaine aldehyde dehydrogenase[J].Euphytica,2011,178:363-372.
[94] 张立全,牛一丁,郝金凤,等.通过花粉管通道法导入红树总DNA获得耐盐紫花苜蓿T0代植株及其RAPD验证[J].草业学报,2011,20(3):292-297.
[95] 张立全,敖登花,师文贵,等.转红树总DNA紫花苜蓿T1代耐盐株系的生理生化特性分析[J].草业学报,2012,21(2):149-155.
[96] Ashraf M,Akram N A.Improving salinity tolerance of plants through conventional breeding and genetic engineering:An analytical comparison[J].Biotechnology Advances,2009,27:744-752.
[97] Chinnusamy V,Jagendorf A,Zhu J K.Understanding and improving salt tolerance in plants[J].Crop Science,2005,45:437-448.
[98] 马金星,张吉宇,单丽燕,等.中国草品种审定登记工作进展[J].草业学报,2011,20(1):206-213.
[99] Zhu J K.Salt and drought stress signal transduction in plants[J].Annual Review of Plant Biology,2002,53:247-273.
[100] 王舟,刘建秀.DREB/CBF类转录因子研究进展及其在草坪草和牧草抗逆基因工程中的应用[J].草业学报,2011,20(1):222-236.
[101] 耿华珠,李聪,李茂森.苜蓿耐盐性鉴定初报[J].中国草地,1990,2:69-72.
[102] Ferradini N,Nicolia A,Capomaccio S,etal.Assessment of simple marker-free genetic transformation techniques in alfalfa[J].Plant Cell Reporter,2011,30:1991-2000.
[103] Ferradini N,Nicolia A,Capomaccio S,etal.A point mutation in theMedicagosativaGSA gene provides a novel,efficient,selectable marker for plant genetic engineering[J].Journal of Biotechnology,2011,156:147-152.
猜你喜欢
青海草业(2022年1期)2022-04-27 06:02:48
今日农业(2021年21期)2021-11-26 05:07:00
河南农业科学(2020年2期)2020-03-11 10:44:04
草业科学(2017年1期)2017-03-14 05:38:41
上海农业学报(2016年2期)2016-10-27 00:49:35
浙江农业学报(2016年7期)2016-06-15 20:30:08
化工进展(2015年6期)2015-11-13 00:27:26
草业科学(2015年4期)2015-02-16 06:21:32
草业科学(2015年7期)2015-02-15 09:18:32
食品工业科技(2014年15期)2014-03-11 18:17:31
