
家蚕绵茧突变性状调查
2012-03-22 09:02:22刘春朱晓楠刘茹凤任静波石虎荀立杰夏庆友
蚕学通讯 2012年1期
刘春 朱晓楠 刘茹凤 任静波 石虎 荀立杰 夏庆友
(家蚕基因组生物学国家重点实验室西南大学生物技术学院,重庆 400716)
家蚕(Bombyx mori)具有吐丝结茧的行为,对正在变态发育的蚕蛹进行保护并防御外来侵害[1]。家蚕在驯养进化过程中出现大量的突变品种,其中在记录的超过400种,性状分布在家蚕发育的各个时期。从20世纪初,科学家们便开始利用突变品系对家蚕进行经典遗传学研究,取得成果颇丰,为家蚕研究打下坚实基础[2]。
随着家蚕基因组的破译,家蚕基因组学迅速发展,各种分子标记建立高密度分子连锁图谱、家蚕多样性指纹图谱以及以克隆为基础的物理图谱[3]相继出现。利用分子标记技术可对影响家蚕生长率、产量、茧丝质量以及抗病性的基因进行打靶,遗传上快速重组遗传物质建立新的品种,令蚕业经济保持可持续发展[3]。
近些年,家蚕定位克隆研究发展迅速,oq油蚕、伴性赤蚁、暗化型、绿茧等突变基因先后被报道定位。各种分子代谢机制也纷纷被解密。
丝腺为家蚕幼虫体内最大的器官,是合成和分泌绢丝的外分泌腺。蚕丝是由两根并行单纤维构成。一般茧丝中丝素占70%~80%,丝胶占20%~30%。
蚕茧主要由茧衣、茧层、蛹衣、蛹体和蜕皮构成。对蚕茧性状的描述,主要有茧形、茧色、全茧量、茧层率等。另外,茧层率因品种、时期、雌雄、化蛹程度、茧型大小、饲育方法环境等不同而呈现差异。此外,衡量蚕茧的经济性状指标还有:茧丝长和茧丝重、解舒丝长和解舒率、出丝率、茧丝纤度等[4]。
家蚕绵茧突变(Flossy,Fl,系统编号:02-220),为自然突变品系,因茧层浮松,茧外形宽大且不规则而得名,由Banno[5]等人于2001年报道。绵茧突变系单基因突变,遵循孟德尔经典遗传规律,其经典连锁位点是第7连锁群的32.1[1]。绵茧突变是家蚕少数有关茧丝形成和营茧行为相关的突变之一,对研究茧丝品质形成和家蚕营茧行为有着重要意义。
1 材料和方法
1.1 材料
夏芳、大造、绵茧品种蚕茧,由西南大学家蚕基因库提供。2009年春季于西南大学蚕学与生物系统研究所蚕房按常规方法饲养夏芳、大造、绵茧品种,上蔟采茧后随机抽样取茧若干做实验材料。
1.2 不同发育时期观察
绵茧和大造不同时期的形态观察比较:对所调查品种的蚁蚕,成虫、丝腺直接进行肉眼和显微镜形态观察。
1.3 茧层率的测量
茧层率的测量方法参考(丝绸学院丝绸教研组.茧丝学实验指导),具体方法如下:随机抽取各品种蚕茧30粒,10粒一组,剥去茧衣,削茧,去蛹,标记。按常规方法称量全茧量、蛹重。将茧放入烘箱内烘至无水恒重(前后称量差不超过0.001g),称其无水干量(标准状态20℃RH60%~65%下称重)[6]。计算茧层率:

1.4 含胶率的测量
含胶率的测量方法参考(丝绸学院丝绸教研组.茧丝学实验指导),具体方法如下:取各品种蚕茧茧层5.00g,在浴比1∶100的中性皂溶液中煮沸进行脱胶,时间约1h,中间更换中性皂溶液一次。后期使用0.83%的碳酸钠溶液、微醋酸水溶液、单蒸水处理洗涤脱胶后的丝素。用胭脂红苦味酸法检验丝胶是否脱尽。最后于烘箱内将丝素烘至无水恒重(前后称量差不超过0.001g),称量[6]。做3次重复。

1.5 茧丝显微镜观察
茧丝显微镜观察的方法参考(丝绸学院丝绸教研组.茧丝学实验指导),具体方法如下:各取夏芳、大造、Fl蚕茧1粒,将蚕茧分为内层、中层、外层;每层剪取小块茧丝,置于载玻片上,使用2%NaOH溶液对茧丝进行脱胶处理,然后单蒸水洗净,做成装片,进行显微镜观察,放大倍数为4×10[6]。
1.6 茧丝丝胶蛋白SDS-PAGE
茧丝显微镜观察的方法参考荣海博硕士毕业论文,具体方法如下:各品种蚕茧,剪碎后,定量为0.10g,使用1×PBS溶解提取丝胶蛋白质,上样电泳[7]。
2 结果与分析
2.1 绵茧的形态观察
2.1.1 不同品种蚕茧形态观察
于常规条件下饲养夏芳、大造、Nd、绵茧蚕品种,将各品种采集的蚕茧分类放置于通风干燥环境下。蚕茧拍照结果如图1所示,图中可见绵茧茧衣与茧层之间界限不明显,蚕茧表现多层且浮松,形状不规则;大造和夏芳品种茧层紧实,大造茧形呈纺锤形,夏芳则呈椭圆形;蚕茧大小上,绵茧体型较大,接近于夏芳。绵茧蛹体型大小接近于大造蛹体。

图1 家蚕不同品种蚕茧比较
2.1.2 不同蚕品种丝腺观察比较
我们对在相同条件下饲养的大造、绵茧和夏芳品种,于5龄第三天时随机抽取三个品种蚕,并解剖取出丝腺进行观察比较,拍照结果如图2所示。图中比较发现,解剖大造、绵茧和夏芳5龄第三天幼虫,三者丝腺之间不存在明显差异,弯折基本一致,夏芳整个丝腺偏大,大造的丝腺偏小,而绵茧的居中。

图2 大造、绵茧和夏芳5龄第3天丝腺比较
2.2 不同品种蚕茧茧层率和含胶率调查
我们对夏芳、大造、绵茧品种的蚕茧的茧丝性状进行调查,主要对全茧量、茧层率以及蚕茧含胶率进行了调查,每个品种随机抽取蚕茧10粒,进行全茧量、茧层量、茧层率调查记录,共重复三次,结果如表1所示。
调查结果显示,夏芳、绵茧、大造茧层率分别为:23.476%、15.551%、12.919%,品种间茧层率为下降趋势;含胶率调查中,三个品种的含胶率分别为:27.182%、29.608%、34.767%,品种间含胶率为上升趋势。绵茧蛹体重与大造蛹体基本一致。综合看来,三个品种茧层率和含胶率之间呈现负相关关系,我们将结果整理,如图3所示。图中可看出,三个品种间,随着茧层率的下降,对应含胶率则呈上升趋势。这里值得注意的是,在绵茧茧层量比大造品种高的情况下,两个品种的蛹体重基本一致。
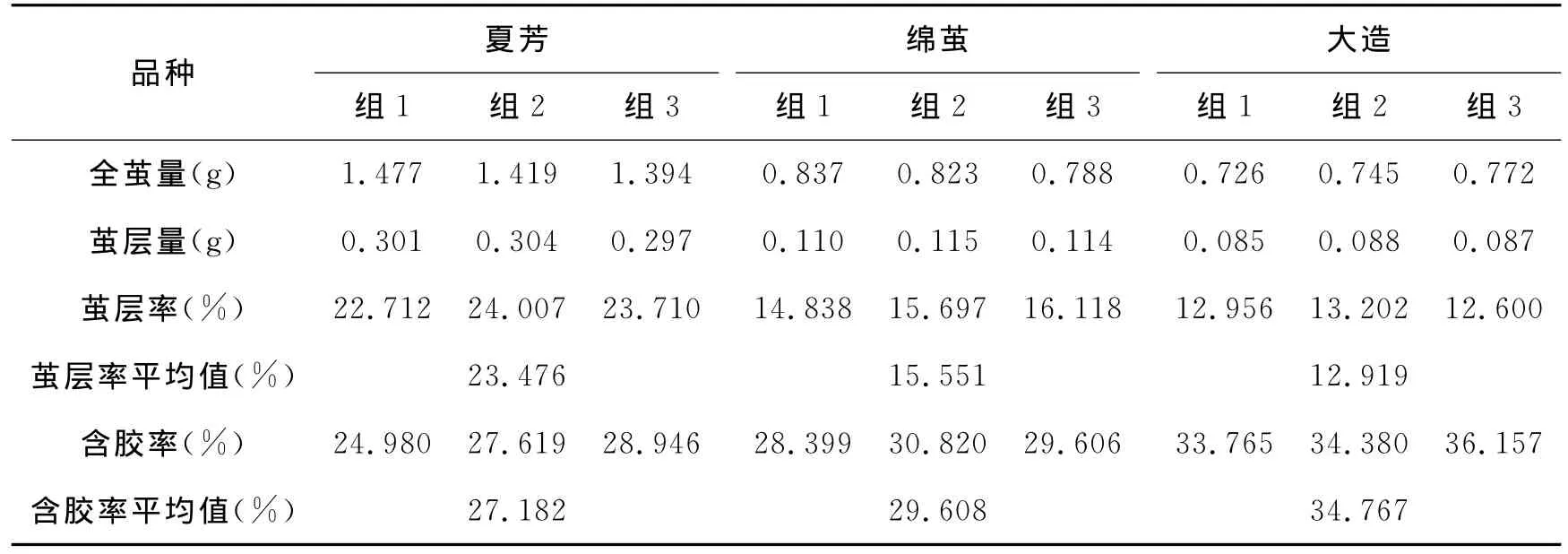
表1 不同品种蚕茧茧丝性状调查
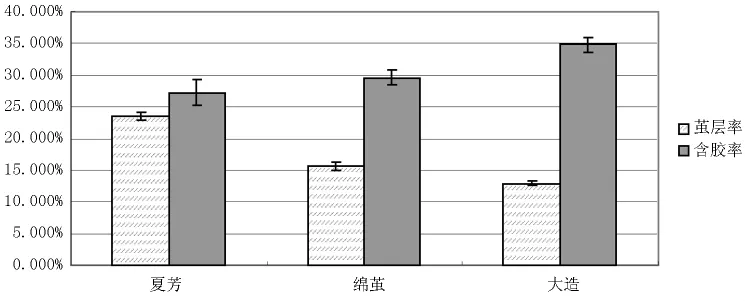
图3 不同品种蚕茧性状调查
2.3 茧丝显微镜观察
比较夏芳、大造、绵茧的内中外层茧层,各品种的茧层用2%NaOH溶液脱胶后,显微镜拍照如图4所示。结果显示,同品种不同茧层之间未发现明显差异;绵茧同其他品种相比,不同品种相同茧层之间没有明显差异。由于茧丝脱胶的原因,因而不排除含丝胶的茧丝存在差异。
2.4 不同品种蚕茧丝胶蛋白电泳
为了比较夏芳、大造、绵茧茧层丝胶蛋白的成分差异,将1×PBS提取的丝胶蛋白进行蛋白质电泳,结果如5所示。图中可以看出,绵茧丝胶蛋白和夏芳、大造相比,成分上没有明显差异。图中可看出,在25KD左右处,各个品种都有一条较为明显的蛋白带,该蛋白是丝素蛋白P25或fib-L蛋白或其它蛋白还有待进一步鉴定。
3 讨论
绵茧突变体在研究和生产上具有很大应用价值,其不仅具有揭露茧丝形成机理的潜力,而且绵茧更适合被应用于蚕丝被的生产。遗传上,绵茧属于单基因突变,遵循经典遗传定律,表型上,绵茧茧层浮松,多分层。通过对绵茧茧层率、含胶率和丝胶蛋白成分分析,希望能找出其茧层浮松的原因,为茧丝形成机理和绵茧定位克隆研究提供线索。
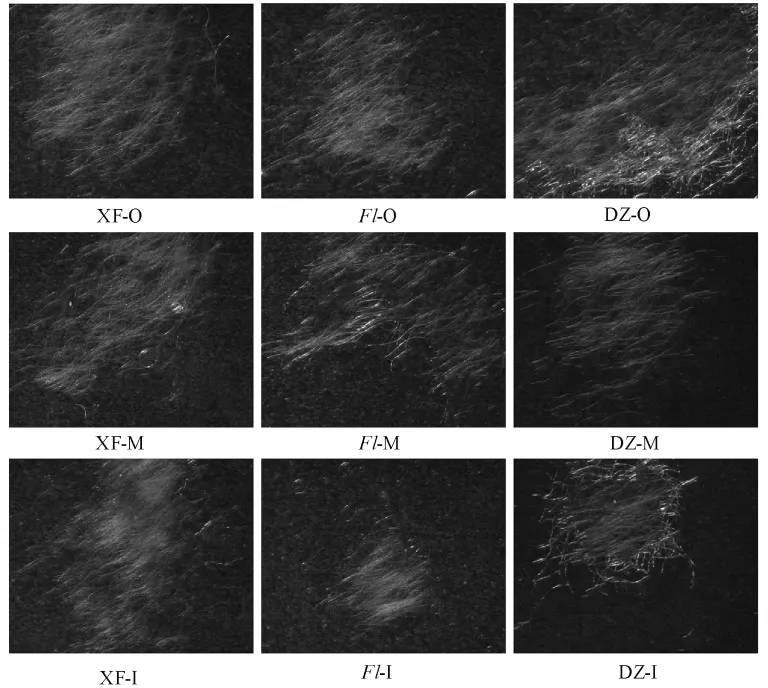
图4 不同品种外中内茧层的茧丝比较

图5 不同品种蚕茧丝胶蛋白电泳
绵茧的形成机制一直是个谜题,绵茧突变体在发育、体形、生理上与野生型相比均没有明显差异,致使茧层浮松和多层的原因一直未知。日本方面也对绵茧突变体幼虫的吐丝行为进行了观察研究,同样未发现明显的差异。本文从茧层率、含胶率和蛋白质水平寻找绵茧与野生型个体的不同,但从茧层率、含胶率和蛋白质电泳结果来看,与正常个体相比,绵茧并未表现出明显的差异。这也意味着绵茧形成的原因仍是一个未知的谜题。而就目前的实验结果看来:其一,绵茧的丝胶蛋白的含量也许是个突破口。而丝胶是茧丝之间胶着的介质,绵茧形成和丝胶蛋白有着十分密切的关系;其二,在绵茧茧层量比大造品种高的情况下,绵茧蛹体重却与大造蛹体基本一致。由于不知绵茧与大造熟蚕体重,故而无法比较得出结果,是否存在绵茧体内含水量较高的可能性需要进一步实验验证。因而,下一步实验将会围绕寻找绵茧丝胶蛋白差异和调查绵茧吐丝过程中脱水量展开。
在目前的家蚕养殖生产中,家蚕上蔟时多湿的环境会造成绵茧发生概率增大。蚕茧性状是由多基因控制,而绵茧突变体属单基因显性突变,即由于单基因突变致使绵茧持续稳定发生。因而笔者推断,突变体有可能是自身原因造成蚕茧内环境形成一个多湿的环境,从而造成绵茧形成,这个问题有待于进一步的实验验证。
[1]向忠怀.蚕丝生物学[M].北京:中国林业出版社,2005.
[2]向仲怀.家蚕遗传育种学[M].北京:农业出版社,1994.
[3]Nagaraju,J.,Goldsmith,M.R.,Silkworm genomics-progress and prospects[J].Current Science,2002,83(4):451-425.
[4]黄国瑞.茧丝学[M].北京:农业出版社,1994.
[5]Banno Y,Hujii H,Doira H.Inheritance of one cocoon shape mutant named“Floossy cocoon”of the silkworm,Bombyx mori[J].Journal of Insect Biotechnology and Sericology,2001,70:189.
[6]丝绸学院丝绸教研组.茧丝学实验指导[M].重庆,2001.9
[7]荣海博.家蚕丝腺特异表达的bHLH转录因子的克隆及蛋白表达[D].重庆:西南大学,2009.
猜你喜欢
四川蚕业(2022年1期)2022-06-06 02:03:56
辽宁丝绸(2021年1期)2021-05-12 06:19:52
四川蚕业(2021年2期)2021-03-09 03:15:34
农学学报(2020年6期)2020-06-29 12:09:00
四川蚕业(2017年2期)2017-07-18 10:58:21
中国纤检(2016年8期)2016-10-22 11:39:48
吉林大学学报(医学版)(2015年4期)2015-12-17 07:48:09
丝绸(2014年12期)2014-02-28 14:56:18
丝绸(2014年12期)2014-02-28 14:56:17
丝绸(2014年12期)2014-02-28 14:56:16
