
科学前沿
2012-01-13 08:52
科学中国人 2012年5期
科学前沿
Nature最新内容精选

银河中心一团气体云正向超质量黑洞运动
黑洞概念的科学提出是在一个世纪前,如何从观测上证明黑洞的真实存在是现代天体物理学中最具挑战性的课题之一。1974年2月发现的射手座射电源SgrA*被认为是位于银河系中心的一个超大质量黑洞的位点,质量约为400万个太阳质量,距太阳系约26,000光年。SgrA*所占区域的直径只有1.5亿公里,由此推断出的最小质量密度比任何目前已知的黑洞候选者的密度都要大了一万亿倍以上。SgrA*因距离最近,而被公认为是研究黑洞物理的最佳目标。最近,恒星轨道研究发现一个以每秒1,700公里的速度向SgrA*移动的物体。该物体低温和光谱特性表明这是一团正落入黑洞的质量为地球三倍的离子气体尘埃云。模型预测云团越接近黑洞,x射线辐射将变得越明亮。如果云团破裂并且气体进入黑洞,在几年内将出现巨大的辐射耀斑。本期封面是流体动力学模型预测2025年被破坏的气体云团与热吸积流的相互作用。

半胱氨酸甲基化修饰破坏NF-kB信号通路中泛素链信号的活性
通过三型分泌系统分泌效应分子进入真核细胞内,进而阻断或调节宿主关键信号转导通路是许多病原菌普遍采用的致病机制。寻找效应分子在宿主细胞中的靶蛋白并阐明其作用于靶蛋白及相关信号通路的生物化学机理对我们了解病原菌致病机理和建立有效防治手段有着重要的意义。同时,这也可能促进我们对真核细胞本身信号转导机制的进一步理解。
许多病原细菌在感染过程中都能有效地抑制宿主细胞中起抗感染作用的NF-kB炎症反应信号通路的激活。北京生命科学研究所邵峰课题组一直对病原菌如何拮抗宿主炎症信号通路的分子机制感兴趣。肠致病性大肠杆菌对NF-kB信号通路的抑制作用依赖于其三型分泌系统分泌的NleE效应蛋白。在这项研究中,研究人员首先通过一系列的细胞生物学实验发现NleE能直接作用于宿主细胞中NF-kB信号通路中的关键信号转导分子TAB2/3。在正常的由细菌感染所诱导激活的NF-kB信号转导过程中,TAB2/3的功能是通过感受来自上游Toll样受体复合物产生的泛素链信号,从而激活TAK1-TAB1-TAB2/3复合物中TAK1分子的激酶活性并实现对下游信号通路的激活。TAB2/3感受泛素链信号的功能是通过其C端保守的锌指结构域(NZF domain)直接结合泛素链而实现的。在进一步的生物化学实验分析中,研究发现NleE效应蛋白具有一种全新的甲基转移酶的活性,这种甲基转移酶能特异性地修饰TAB2/3锌指结构域中一个螯合锌离子的半胱氨酸。这种新颖的修饰作用导致NZF结构域中锌离子的离去和该结构域的构象变化。无论是在体外的重组实验还是肠致病性大肠杆菌感染的细胞中,被NleE效应蛋白甲基化修饰的TAB2/3都彻底丧失了结合泛素链的功能。这些实验有力地说明了NleE确实是通过甲基化修饰TBA2/3锌指结构域中的半胱氨酸而实现对宿主NF-kB信号通路的抑制作用的。
这项研究不仅揭示了一种新的病原菌效应蛋白阻断宿主炎症信号通路的机制,更为重要的是,这篇文章也首次报导了半胱氨酸甲基化作为一种新的翻译后修饰在调节信号转导中起关键作用。鉴于锌指结构域是一种最为广泛存在的结构域,同时正是由于锌离子的螯合作用才使得TAB2/3中的半胱氨酸能够被NleE所甲基化,这项研究发现也暗示真核细胞本身的蛋白也很有可能存在半胱氨酸甲基化这种翻译后修饰和调控机制。另外,最近在NF-kB信号通路的研究中存在一个到底是线性泛素链还是Lys63连接的泛素链哪个更为重要的争论,鉴于TAB2/3只感受Lys63连接的泛素链并且NleE对NF-kB通路中的其它泛素链结合蛋白没有修饰作用,这项工作从一个独特的角度说明了Lys63连接的的泛素链确实对NF-kB信号转导至关重要。
艾滋病毒侵袭机制:T细胞分化因子调控HIV-1Vif介导的宿主防御逃脱
HIV分为两种亚型,HIV-1和HIV-2,世界上大部分艾滋病患者都是由HIV-1感染的。人体都有一个抗HIV的天然防御系统,不过HIV病毒利用自身的蛋白(Vif)把这个屏障给破坏掉了,所以病毒可以复制。HIV-1Vif是由保守的vif基因编码的磷酸化蛋白, 它在产生感染性的病毒颗粒中起主要作用。Vif蛋白不能表达或其功能受到限制,会很大程度上减少病毒颗粒的产生。吉林大学白求恩第一医院于晓方教授领导的科研团队找到了与病毒蛋白相互作用的宿主体内的T细胞分化因子CBF-β,并阐述了其与HIV Vif相互作用。CBF-β因子在病毒侵袭和复制过程中有着重要的调控作用,可以说是宿主天然防御系统的总开关,艾滋病毒就是通过它突破了宿主天然防御系统完成复制的。在没有CBF-β存在的情况下,病毒无法抵抗宿主体内的天然抗病毒因子,从而无法完成复制。该项研究揭示了一个全新的艾滋病毒逃避机制,如何把这个逃避机制封闭住,使宿主体内的天然防御系统重新恢复作用,是今后抗病毒研究的一个新靶点。这一发现为新型的抗H I V抑制剂的发明提供了理论基础,同时为艾滋病鸡尾酒疗法添加药物新成员开辟了新的方向,对HIV的治疗具有重大意义。

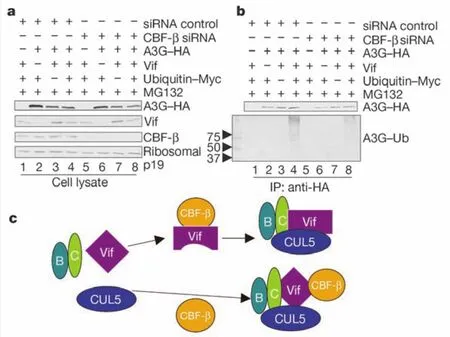
图1、V if诱导的A P O E C3G泛素化需要CBF-β因子。APOBEC3G能作为抑制分子阻止病毒到达其目的地。
狩猎-采集者间的社会网络和协作
坦桑尼亚北部的Hadza人与现代社会几乎是完全隔绝的,这个以狩猎和采集果物为生的流动部落人口只有750人。一个典型的Hadza群体由十多名男女及儿童组成。人类定居并饲养动物、耕作农田的经验只有约11,000年的历史,而在此之前的漫长岁月里,所有人类都像Hadza人一样以狩猎和采集为生。因此当人类学家需要考察早期人类的生活形态,特别是社会分工以及部落间的合作或竞争关系时,Hadza人为研究者提供了宝贵的材料。提出“祖母假说”的几位人类学家正是国际上研究Hadza人的权威。对Hadza人社会网络所做的一项新的研究工作(将他们彼此之间的关系及他们合作的倾向性进行量化)表明,现代化社会网络的主要特征,如“传递性”和“对同类之爱”,也见于Hadza人。而且,社会关系更可能在公共财产游戏中具有相似捐出水平的个体间产生,Hadza人部落的表现是:群体间合作的变化大,群体内合作的变化小。这些结果综合起来,提供了迄今最有力的证据,证明人类社会网络的这些关键特征反映了我们具有共同的祖先,并且可能是在人类历史中早期的某个时间点上形成的。本期封面图片:两名正在采摘浆果在采摘浆果的Hadza妇女。

Science最新内容精选

蜡烛烟灰作为透明强化超疏水材料涂层的模板
涂层是调整材料表面特性的重要一步。基于纳米和微米尺度上低能表面和粗糙度,研究人员已开发出了接触角大于150o、滚动角小于10o的超疏水涂层。然而,这些疏水表面仍然会被有机溶液浸润,如表面活性剂溶液、乙醇或烷烃,没有涂层能同时具有超疏水性和超疏油。我们设计了一种容易制作的、透明、可反弹油滴的超疏水涂层。以25nm厚的硅壳包裹蜡烛烟灰多空沉淀。600oC煅烧后,黑色涂层变得透明。硅化后,涂层具有超疏水性。在被沙子冲击破坏表层后,涂层依然可以保持超疏水性。这种价格低廉且容易规模化的技术可应用于触摸屏或眼镜等的防指纹涂层。本期封面:一只燃烧的蜡烛。

图2(右)、多种形态的多空结构。(A)样品准备,将载玻片置于蜡烛火焰中直至沉积几微米厚的烟灰。(B)烟灰沉积物的扫描电镜图像。(C)高解析度扫描电镜图像显示,一个颗粒链由若干直径40±10nm的圆形碳株组成。(D)包裹硅壳后的扫描电镜图像。(E)600oC加热2小时移除碳核后,一个颗粒簇的高解析度扫描电镜图像。(F)煅烧后的颗粒簇的高解析度扫描电镜图像,硅涂层内的空洞,这些空洞煅烧前为碳核心,硅壳厚20±5nm。

图3(下)、超疏水表面。(A)2微升水滴,接触角165o±1o。(B)5微升十六烷滴,接触角156o±1o。(C)沉积在分形复合材料表面液滴的示意图。(D)十六烷滴在超疏水表面上弹跳的时间解析图。
信天翁分布和生活史的变化
本期封面是一只迁徙途中的年轻信天翁正快速掠过南半球大洋海面,信天翁借助于海风高效迁移。最近,南半球大洋上西风带的变得更强了,并向南极移动。长期的观察记录显示,海风变化使得信天翁能够更快的迁移并且其觅食范围也向南移动。因此,信天翁的觅食周期缩短,繁殖成功率提高,并且鸟体重也增加了1公斤,这对信天翁的繁殖和保护产生了积极的作用。如果南半球西风带的变化与预测气候情况一致,那么这些由气候变化产生的积极效果可能都是暂时的。本项研究还强调觅食的重要性在于其为环境变化和繁殖过程间的关键联系。

褐海藻直接生产生物燃料
本期封面是智力奇洛埃岛沿岸1米深海中巨大的褐藻(Macrocystispyrifera)。海藻通过生物转化生产生物燃料和日用化学品的主要障碍是缺乏能够代谢褐藻多糖的微生物。最近,研究人员从灿烂弧菌(Vibriosplendidus)中发现了编码海藻多糖运输和代谢酶的基因片段。通过基因工程将该基因构建成细胞外海藻聚糖降解系统,构建出一个可以同时降解、摄取和代谢海藻糖的微生物平台,进而构健成乙醇合成系统。该平台可以固态工艺直接将褐藻转化为生物乙醇,每吨干褐藻可以产0.281吨乙醇,相当于褐藻中所含糖的最大理论产量的80%。

神经系统外朊病毒更易在不同物种间传递
本期封面是一张朊病毒的错色透射电镜图像(约150nm长,10nm厚)。朊病毒是一种传染性病原体,能引发动物和人类的致死性脑部疾病。主要由一种PrPSc蛋白组成,PrPSc蛋白是宿主错误折叠的PrPC蛋白。人们一直认为存在一种PrPC与PrPSc之间限制性立体相互作用使朊病毒具有物种特异性,并限制朊病毒在物种间传播,包括人类。最新研究表明,虽然跨物种朊病毒传染可能在大脑中没有任何可见的致病迹象,但是淋巴组织却极易受到了感染的损害。最近,研究人员比较了绵羊和人类PrP转基因小鼠大脑和淋巴组织复制其它物种朊病毒的能力。淋巴组织总是比大脑更容易容纳朊病毒颗粒,造成慢性消耗性疾病和牛海绵状脑病。朊病毒会聚集在脾脏内,如果出现损伤,病毒向脑转移的障碍被突破,宿主很有可能会患病。因此,朊病毒的跨物种传播可能具有明显的组织依赖性。可能更多的动物和人类可能携带这种疾病。

图4、仓鼠Sc237朊病毒向携带绵羊PrP基因tg338小鼠(白色)和仓鼠PrP基因tg7小鼠(灰色)转移的示意图。

猜你喜欢
中国生物化学与分子生物学报(2022年4期)2022-09-07
生物化学与生物物理进展(2022年8期)2022-08-20
湖北农业科学(2022年11期)2022-07-18
心血管病学进展(2021年8期)2021-09-13
湖北农业科学(2020年24期)2021-01-21
实用肿瘤学杂志(2020年4期)2020-12-08
当代水产(2020年3期)2020-06-15
小星星·阅读100分(高年级)(2015年11期)2015-11-28
大自然探索(2015年11期)2015-09-10
医学研究杂志(2015年12期)2015-06-10

- 科学中国人的其它文章
- 高考该怎样考?
- 最新科协动态
- 首都科学讲堂内容介绍