
银杏抗寒优良无性系选育
2012-01-05 09:03郁万文曹福亮汪贵斌
中南林业科技大学学报 2012年12期
郁万文,曹福亮,汪贵斌
(南京林业大学 森林资源与环境学院, 江苏 南京 210037)
银杏抗寒优良无性系选育
郁万文,曹福亮,汪贵斌
(南京林业大学 森林资源与环境学院, 江苏 南京 210037)
采用-20℃低温处理方法, 分析了39个银杏无性系枝条的LT50,可溶性糖含量、可溶性蛋白含量,SOD活性等指标的变化。运用多元统计主成分分析,初步判断受试材料抗寒性,由强到弱排序为: 16#、38#、17#、40#、41#、36#、45#、15#、53#、44#、19#、42#、57#、10#、35#、18#、50#、55#、32#、26#、12#、30#、56#、9#、13#、34#、12#、37#、14#、33#、54#、20#、28#、11#、31#、27#、29#、39#。其中抗寒性较高的银杏无性系有16#、41#、36#、45#、19#、17#、38#、40#,表现为较好的保水能力、抗氧化能力和渗透调节能力,从而表现出较强的抗寒力,具有在高纬度或高海拔地区推广试验的潜力。
银杏;无性系;冰冻胁迫;抗寒性;选育
选用抗寒性较强的树种、种源、品种或无性系等是解决树木引种受冻的根本途径。不同种类、同一种类不同品种、同一品种不同生育期、不同器官、组织等抗寒力均表现出极大差异[1]。不同银杏单株、品种、无性系的遗传背景和生境存在较大差异, 因此在植株形态、抗寒性等方面均表现不同。其中抗寒性差异直接影响银杏在高纬度或高海拔地区的引种和种植,因此在高纬度或高海拔地区引种银杏之前,对引种无性系或品种进行抗寒性检测是必须的。在极端气候下,一个树种大面积受冻害死亡或树势衰弱,而保留下来的健壮单株则具有相当的抗寒性,将这种单株做为母本,经过抗寒力测定、选育,就可得到优良的抗寒无性系。因此,常有冻害发生的地区,引种银杏的同时,对各品种或无性系的抗寒力要做到胸中有数,选择抗寒力强的品种作为下期引种的对象,同时要特别注意引种地抗寒力强的品种、品系、无性系引种范围的扩大,争取从根本上解决银杏高纬度和高海拔引种受冻问题。本研究用-20℃对39个嫁接18年生的优良核用无性系的越冬枝条进行冰冻处理,测定其相关的生理生化指标,对39个核用无性系的抗寒性进行综合评定,初步筛选出抗寒性较强的无性系,为下一步在高海拔、高纬度地区的区组试验及银杏边缘分布区的引种提供理论依据和抗寒种质资源。
1 材料与方法
1.1 供试材料
材料来源于江苏省邳州市陈楼镇银杏种质资源圃,为嫁接18年生的39个核用银杏无性系。
1.2 方法
2008年1月20日采集生长健壮、粗细均匀的树冠外围的一两年生枝条,用自来水冲洗数遍,再用蒸馏水冲洗3次,然后用吸水纸吸干水分,最后将枝条末端进行蜡封,将每个无性系蜡封后的枝条分成相等的8份,其中1份用于低温处理,剩余7份用于半致死温度测定,放于冰箱中4℃保存。每次处理时,取参试材料的1份枝条放于程控冰箱内进行-20℃冰冻处理,降温速度4℃/h,达到目的温度后维持24 h, 然后逐步升温,升温速度亦为4℃/h。每处理重复3次。
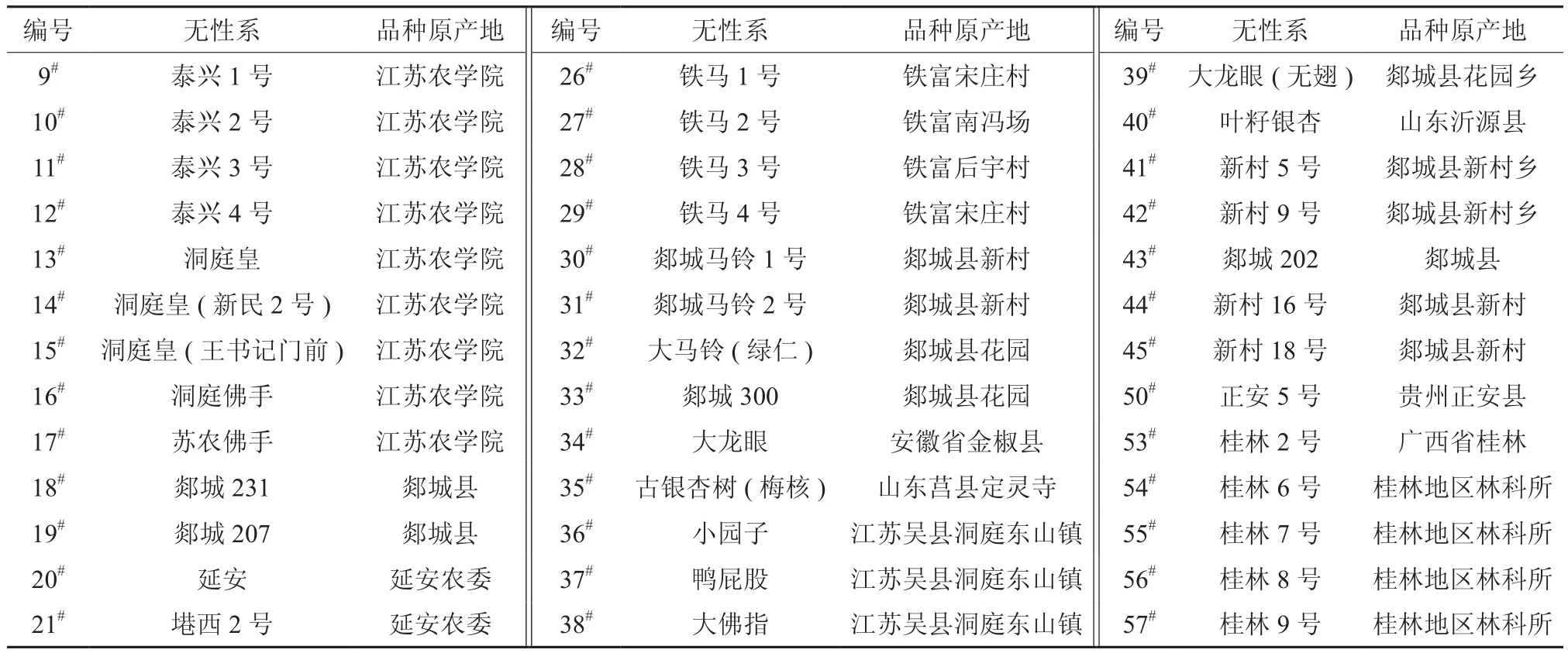
表1 银杏核用无性系编号及原产地Table 1 Number and origin place of stone-used ginkgo strains
1.2.1 相对电导率的测定和半致死温度估算
称量每个无性系枝条7份,每份10.0 g剪成1.0 cm小段,密封在塑料标本袋内。1份在室温下测定电导率作为对照(CK),另外6份放入超低温冰箱内进行冰冻处理。冰冻处理温度分别为-15℃、-20℃、-25℃、-30℃、-35℃和-40℃。冷冻时降温幅度和解冻时的升温幅度都为4℃/h,冷冻到达所处理的温度后,维持24 h,然后解冻,3次重复。相对电导率的测定和半致死温度的估算参照焦奎宝等[2]的方法。
1.2.2 生理生化指标测定
采用蒽酮比色法测定淀粉和可溶性总糖含量,考马斯亮蓝法测定可溶性蛋白含量,磺基水杨酸法测定脯氨酸含量,均参照李合生的方法[3],略有改动。超氧物岐化酶(SOD)、过氧化物酶(POD)、丙二醛(MDA )和氧自由基(·O2-)产生速率的测定参照文献[4]的方法,略有改动。
采用多元统计主成分分析对39个银杏无性系进行分析,计算银杏无性系抗寒力,对银杏无性系的抗寒性进行分类,并进行分析讨论。
2 结果与分析
2.1 不同银杏无性系抗寒指标的差异
2.1.1 不同银杏无性系半致死低温的差异
如表2所示,不同银杏无性系枝条的LT50差异达到极显著水平(P<0.01)(见表3)。其中16#无性系的LT50最高,为-29.06℃,28#无性系的LT50最低,为-18.94℃,两者相差10.12℃。供试的39个核用无性系的LT50由低到高的顺序(见表2)是 16#、17#、38#、40#、53#、44#、15#、42#、45#、36#、57#、43#、19#、41#、18#、13#、10#、30#、35#、50#、33#、56#、26#、9#、20#、55#、32#、54#、37#、34#、11#、21#、14#、31#、12#、29#、27#、39#、28#。
2.1.2 不同银杏无性系可溶性糖含量的差异
如表2和表3所示,不同银杏无性系枝条的可溶性糖含量差异性显著(P<0.05)。其中17#的可溶性糖含量最高,为2.78 mg/gFW,39#的最低,为1.52 mg/gFW,两者相差0.83倍。供试的39个核用无性系的可溶性糖含量为39#<29#<54#<27#<20#<11#<31#<34#<13#<21#<18#<9#<26#<55#<28#<37#<56#<33#<12#<32#<14#<50#<53#<30#<42#<10#<57#<35#<43#<15#<19#<44#<36#<45#<38#<40#<16#<41#<17#。
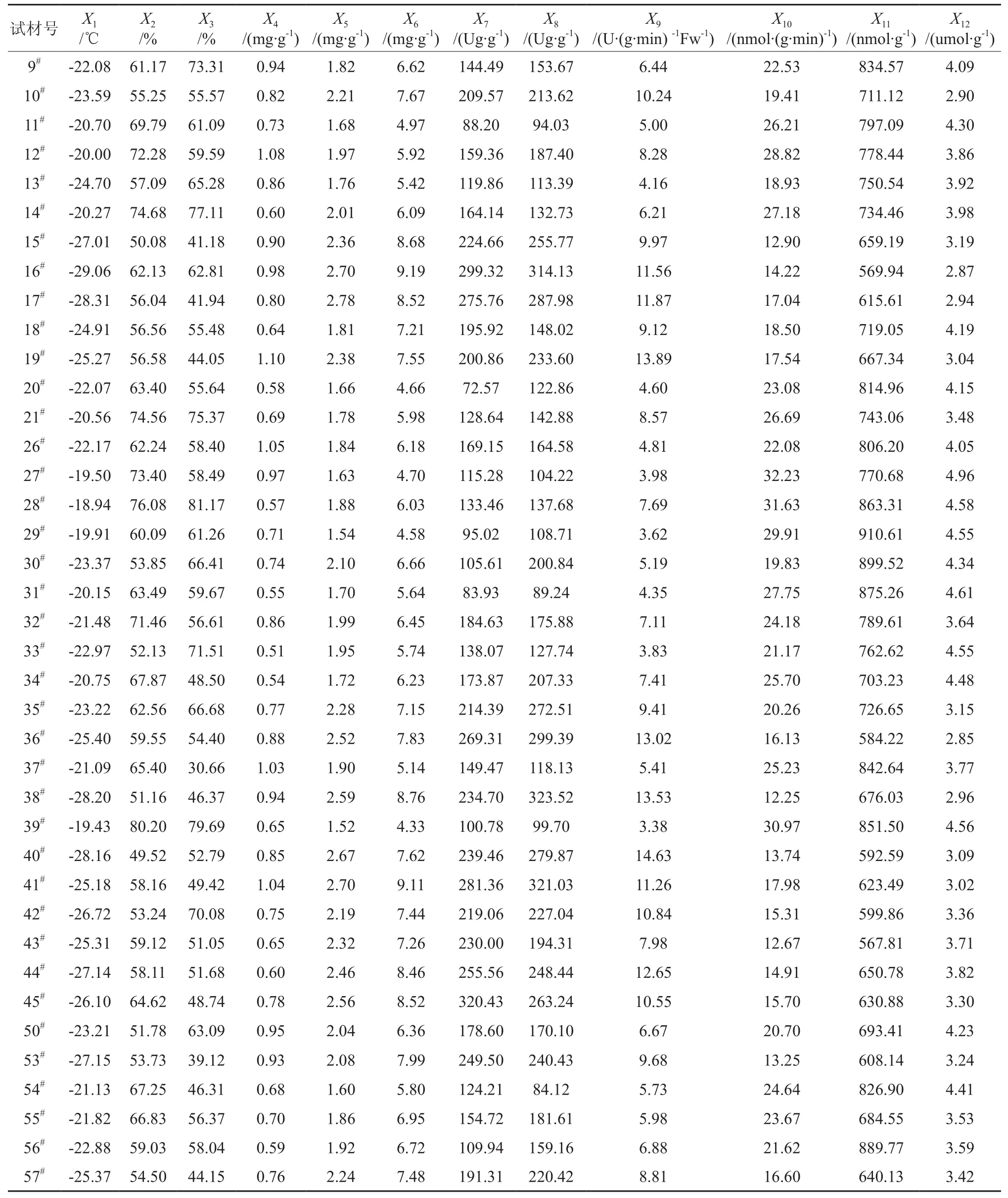
表2 不同银杏无性系枝条抗寒指标†Table 2 Cold-resistance indexes of different ginkgo clones
2.1.3 不同银杏无性系可溶性蛋白含量的差异
由表2和表3可知,不同银杏无性系枝条的可溶性蛋白含量差异性显著 (P<0.05)。其中16#的可溶性蛋白含量最高,为9.19 mg/gFW,39#的最低,为4.33 mg/gFW,两者相差1.12倍。供试的39个核用无性系的可溶性蛋白含量由低到高的 顺 序 ( 表2) 是 39#、29#、20#、27#、11#、37#、13#、31#、33#、54#、12#、21#、28#、14#、26#、34#、50#、32#、9#、30#、56#、55#、35#、18#、43#、42#、57#、19#、40#、10#、36#、53#、44#、17#、45#、15#、38#、41#、16#。
2.1.4 不同银杏无性系SOD活性的差异
如表2和表3所示,不同银杏无性系枝条的SOD活性差异不显著(P>0.05)。其中38#无性系的SOD活性最高,为323.52 U/gFw,54#无性系的最低,仅为84.12 U/gFw。供试的39个核用无性系的SOD活性的顺序(表2)是38#>41#>16#>36#>17#>40#>35#>45#>15#>44#>53#>19#>42#>57#>10#>34#>30#>43#>12#>55#>32#>50#>26#>56#>9#>18#>21#>28#>14#>33#>20#>37#>13#>29#>27#>39#>11#>31#>54#。
其他抗寒指标不再一一详述,详见文献5。对其他抗寒指标进行方差分析,结果表明,39个银杏无性系的失水率、H2O2含量、MDA含量的差异性均达到极显著性平(见表3),相对电导率、淀粉含量的差异性显著,而游离脯氨酸含量、POD活性、产生速率的差异性不显著。
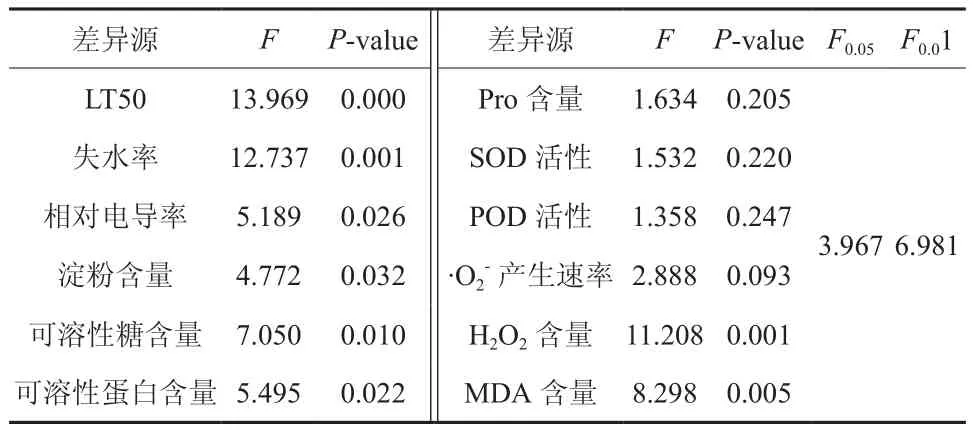
表3 不同银杏无性系抗寒指标方差分析Table 3 Variance analysis on cold-resistance indexes of 39 ginkgo strains
2.2 银杏无性系抗寒指标间相关性分析
对39个银杏无性系的12个抗寒指标进行相关分析,其抗寒生理化指标的相关阵(见表4)。由表4可知,半致死温度与可溶性糖含量、可溶性蛋白含量、Pro含量、SOD活性、POD活性呈极显著负相关,失水率、电导率、生速率、H2O2和MDA含量与半致死温度呈极显著正相关,说明银杏枝条在冰冻胁迫下通过增加保护酶(SOD、POD)活性和渗透调节物质(可溶性糖、蛋白质和Pro)含量,来降低膜脂的过氧化(MDA),维持膜的稳定性(相对电导率),从而提高银杏的抗寒性。另外,SOD和POD活性分别与·O·-产生速率和H2O2含量呈极显著负相关,而H2O2含量与MDA含量则呈极显著正相关,表明冰冻低温下银杏枝条通过保护酶活性的提高,降低活性氧在体内的积累,防止膜脂的过氧化作用。

表4 各抗寒指标间的相关系数Table 4 Correlation coeffeients of cold-resistance indexes
2.3 抗寒指标主成分分析
采用主成分分析方法,对39个银杏无性系进行抗寒性指标综合评价。主成分分析的数据由表5和表6提供。根据累积贡献率≥85.00%的标准,本研究中有3个主成分入选,其累积贡献率为86.18%,前3个主成分已反映了12个抗寒指标的绝大部分信息,因此,只需讨论前3个主成分即可。

表5 各主成分的特征根和贡献率Table 5 The eigenvalues and pereentage aeeumulated contribution of the 3 prineipal components
由表6可见,第一主分量中X4、X5、X6、X7、X8、X9为正向 载荷,Xl,X2、X3、X10、X11、X12为负向载荷,各指标对第一主分量的贡献较为均匀,表明第一主分量是反映银杏无性系在低温下生理生化发生逆变的综合指标。银杏枝条在低温胁下通过增强抗氧化能力(提高SOD、POD活性)和渗透调节能力(可溶性糖、Pro、可溶性蛋白质含量增加),来降低膜脂的过氧化(MDA),维持膜的稳定性(相对电导率),从而提高银杏的抗寒性。第二主分量主要反映了失水率与抗寒性的关系。第三主分量主要反映了枝条电导率和淀粉含量与抗寒性的关系。
2.4 银杏无性系抗寒性划分
选用前两个主成分得分值绘制二维坐标图(见图1)。根据二维坐标图,39个银杏无性系抗寒性强弱可划分为3大类:第一类,抗寒性强{16#、41#、36#、45#、19#、17#、38#、40#};第二类,抗寒性较强 {35#、10#、42#、57#、44#、43#、53#、15#};第三类,抗寒性中等 {9#、26#、13#、30#、50#、18#、56#、33#};第四类,抗寒性弱 {11#、29#、20#、31#、54#、12#、27#、37#、32#、55#、34#、14#、21#、28#、39#}。
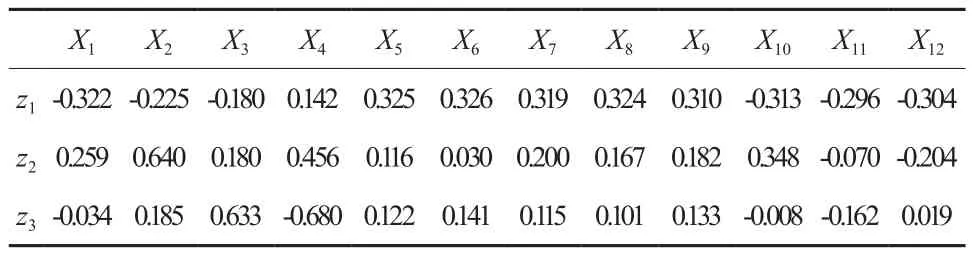
表6 前3个主成分分析因子载荷Table 6 Load of principal components of first 3 factors
对上述4类无性系的指标进行分析,第一、二类,尤其是第一类,具有较好的保水能力、抗氧化能力和渗透调节能力,从而表现出较强的抗寒力,其中第一类具有在高纬度地区推广试验的潜力。第三类,相对电导率和H2O2含量最高,推测高水平的H2O2,破坏了膜的半透性,造成低温胁迫下枝条中内含物的大量外渗,表现为相对电导率很高,致使该类抗寒性较差。第四类,保水能力、抗氧化能力和渗透调节能力均较差,从而表现出较差的抗寒性。
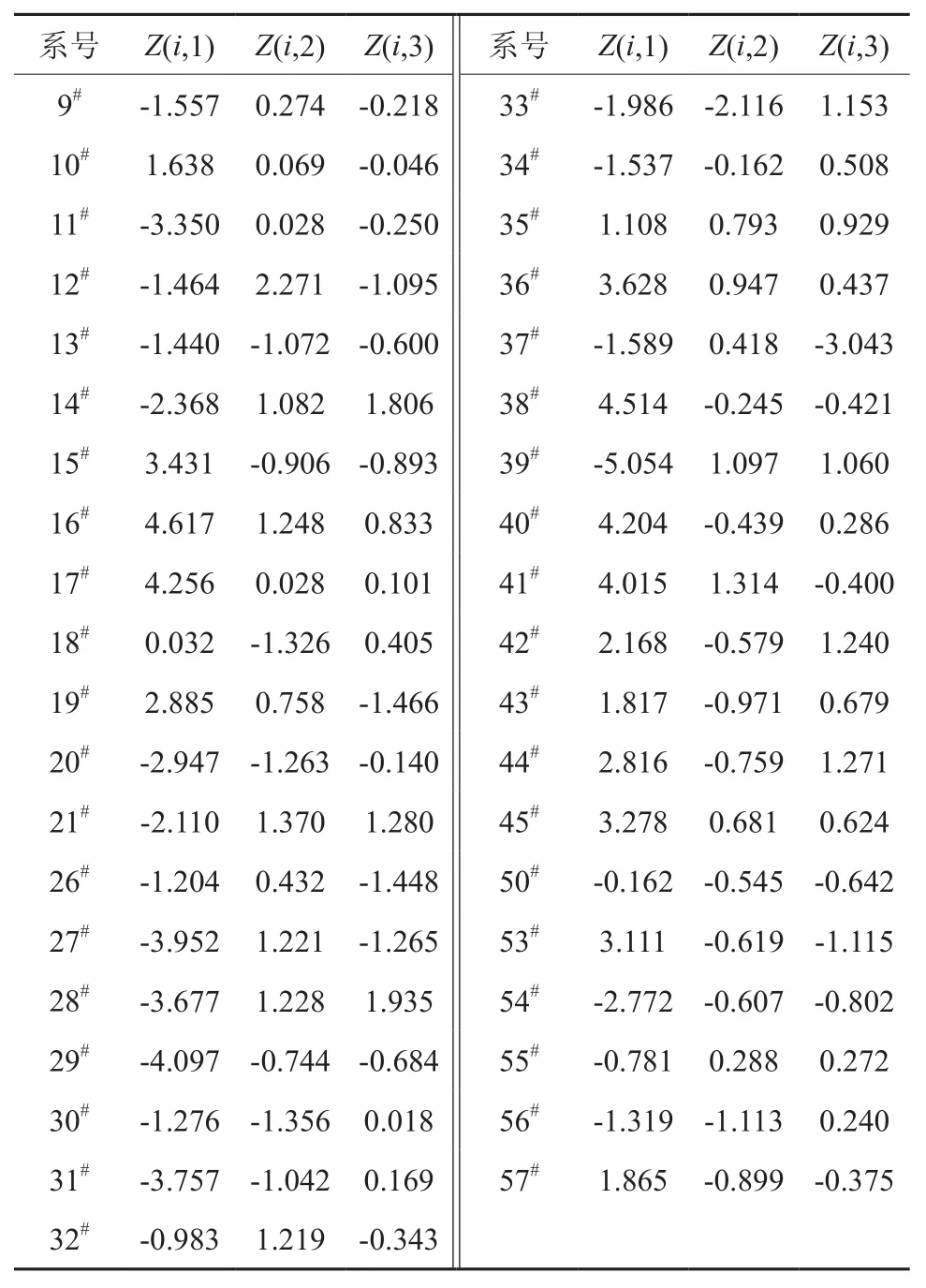
表7 银杏无性系的前3个主成分的得分值Table 7 The values of the first 3 prineipal components

图1 主成分分析二维坐标Fig.1 Two dimensional coordinate graphs of principal component analysis
2.5 银杏无性系抗寒性排序
表7列出了39个银杏无性系的主成分值和表5列出的贡献率,根据公式Σni=1AjBj(A:主成分贡献率,B:银杏无性系主成分值,j:1~3,n:39),可求出39个无性系抗寒性大小及抗寒力顺序(见表8)。39个银杏无性系抗寒力的顺序如下:16#>38#>17#>40#>41#>36#>45#>15#>53#>44#>19#>42#>57#>10#>35#>18#>50#>55#>32#>26#>12#>30#>56#>9#>13#>34#>12#>37#>14#>33#>54#>20#>28#>11#>31#>27#>29#>39#。
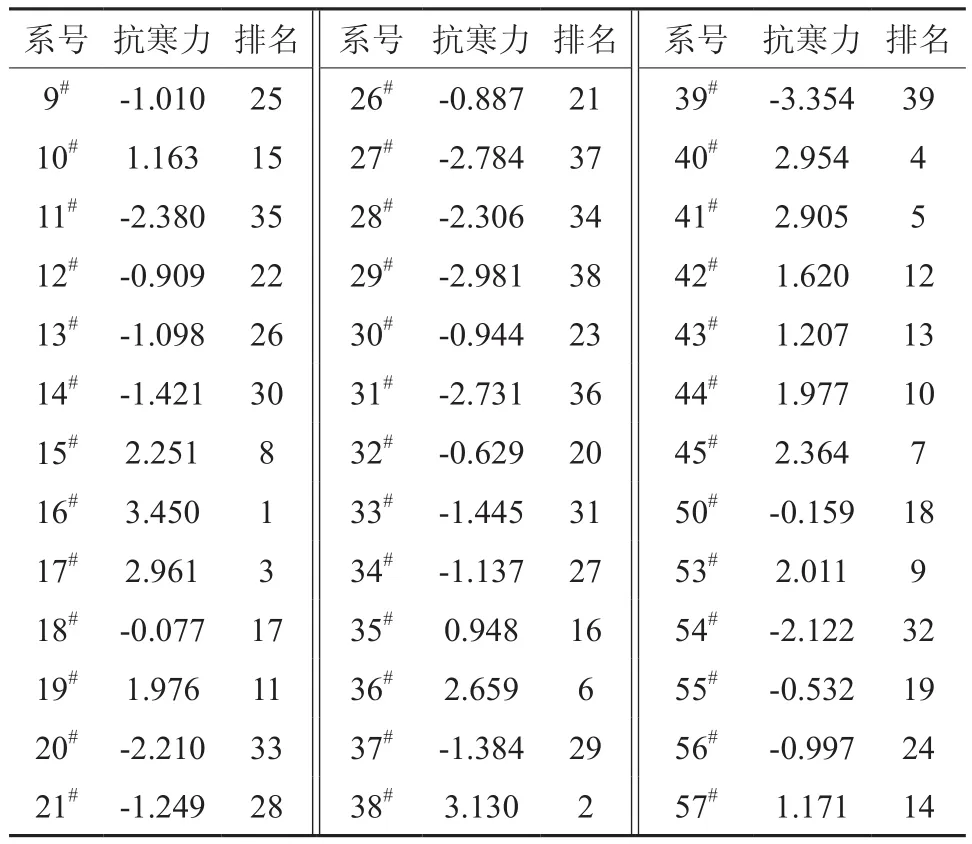
表8 银杏无性系抗寒力排序Table 8 Cold resistance ability ranking of ginkgo strains
3 讨论与结论
在冰冻低温下,如何有效的避免胞内结冰是林木抵抗冰冻胁迫的第一步。低温胁迫下, 植物体的抗寒性越强, 可溶性蛋白含量也越高。可溶性糖能够减少低温对细胞的伤害, 在低温下讨论可溶性糖含量时, 常常提及可溶性蛋白含量, 它们常作为鉴定植物抗寒性的指标[6]。植物体内游离脯氨酸对机体器官具有保护作用, 可作为植物抗寒性测定的指标。许多研究表明,这些物质具有降低细胞的渗透势和组织冰点,稳定膜系统,保护蛋白质和调节抗性基因表达等多方面的作用,与抗寒性变化密切相关[7-8]。本研究表明,越冬过程中,银杏枝条主动转化生成大量的可溶性糖、Pro等渗透调节物质,调节细胞中渗透势,降低组织冰点,保护蛋白质。同时低温诱导抗寒基因的表达,合成了大量与抗寒相关的蛋白类物质,如抗寒蛋白、抗寒调控性酶类,造成可溶性蛋白质含量的增加。
低温胁迫会使植物体自由基产生速率升高[9]、清除剂含量降低[10]、清除酶类的活性不均衡的降低[11-12],植物细胞内活性氧的积累,引发膜脂过氧化、交联蛋白质和攻击DNA等,而且其氧化产物多会对植物造成严重的伤害。大量研究表明,低温锻炼可诱导植物体内活性氧清除酶类的活性提高或增加其在低温下的稳定性,提高体内的活性氧清除剂,如SOD、POD、GSH活性,从而有效地避免活性氧对蛋白尤其是酶蛋白伤害[8-9,13]。本研究表明,低温胁迫下银杏体内保护酶类,SOD、POD等活性升高,有效地清除活性氧自由基的产生,维持较好的活性氧代谢平衡,且酶活性高的无性系其抗寒性强,这与阐文靖等[14]的研究结果一致。
综上所述,部分银杏无性系的枝条在低温胁下能够通过增强抗氧化能力(提高SOD、POD活性)和渗透调节能力(可溶性糖、Pro、可溶性蛋白质含量增加),来降低膜脂的过氧化(MDA),维持膜的稳定性(相对电导率),表现出较强的抗寒性。39个银杏无性系枝条抗寒性的顺序是16#、38#、17#、40#、41#、36#、45#、15#、53#、44#、19#、42#、57#、10#、35#、18#、50#、55#、32#、26#、12#、30#、56#、9#、13#、34#、12#、37#、14#、33#、54#、20#、28#、11#、31#、27#、29#、39#。其中抗寒性较高的银杏无性系有16#、41#、36#、45#、19#、17#、38#、40#,表现为较好的保水能力、抗氧化能力和渗透调节能力,从而表现出较强的抗寒力。所以这8个无性系具有在高纬度地区推广试验的潜力。
[1] 刘 艳,李晓燕,王丽雪,等.苹果枝条冬季淀粉粒动态变化与抗寒力的关系[J].内蒙古农业大学学报,2006,27(2):79-83.
[2] 焦奎宝,高庆玉.葡萄嫁接苗根系低温半致死温度的测定[J].黑龙江农业科学,2010, (3):67-68.
[3] 李合生.植物生理生化实验原理和技术[M].北京:高等教育出版社,2000.
[4] 中国科学院上海植物生理研究所,上海市植物生理学会.现代植物生理学实验指导[M].北京:科学出版社,1999.
[5] 郁万文.银杏抗寒机理及种质资源抗寒性评定的初步研究[D].南京:南京林业大学,2008.
[6] 司剑华,卢素锦.低温胁迫对5 种柽柳抗寒性生理指标的影响[J].中南林业科技大学学报,2010,30(8):78-81
[7] 朱宁华,李志辉,李芳东.桉树耐寒性与超氧化物歧化酶关系研究[J].中南林业科技大学学报,2000,20(3):63-66.
[8] 张蜀宁,张振超,张红亮,等.低温胁迫对不同倍性不结球白菜生长及生理生化特征的影响[J].西北植物学报,2008,28(1):109-112.
[9] 李 晶,阎秀峰,祖元刚. 低温下红松幼苗活性氧的产生及保护酶的变化[J]. 植物学报,2000,42(2): 148-152.
[10] 陈少裕.膜脂过氧化对植物细胞的伤害[J].植物生理学通讯.1991,27(1):84-90.
[11] Hull Mark R, Stephen P L, Leland S J. Instantaneous and developmental effects of low temperature on the catalytic properties of antioxidant enzymes in two Zea species [J]. Aust. J.Plant Physiol, 1997, 24:337-343.
[12] Seebba Franeesea, Luca Sebastiani, Claudio Vitagliano. Changes in activity of antioxidative enzymes in wheat (Tririeum aesrivum)seedlings under cold acclimation [J]. Physiologia Plantarum,1998, 104:747-752.
[13] Zhao SY, Eduardo B. Changes in oxidation-reduetion state and antioxidant enzymes in the roots of jack pine seedlings during cold acclimation [J]. Physiologia Plantarum, 1998, 104:134-142
[14] 阐文靖,朱宁华,邱运亮. 桉树对低温反应的生理生态学基础[J]. 经济林研究,1994,12(增刊):1-6.
Selection and breeding of Ginkgo cold-resistant fi ne clones
YU Wan-wen, CAO Fu-liang, WANG Gui-bin
(College of Forest Resources and Environment, Nanjing Forestry University, Nanjing 210037, Jiangsu, China)
By using -20℃low temperature processing methods, the changes of semilethal temperature, soluble sugar, soluble protein contents, SOD activity and others of 12 indexes measured for 39 Ginkgo clones branches were analyzed. The cold resistant performance of the tested materials were sorted from strong to weak : 16#,38#,17#,40#,41#,36#,45#,15#,53#,44#,19#,42#,57#,10#,35#,18#,50#,55#,32#,26#,12#,30#,56#,9#,13#,34#,12#,37#,14#,33#,54#,20#,28#,11#,31#,27#,29# and 39#. Of them, the clones having relatively stong cold resistant were 16#,41#,36#,45#,19#,17#,38# and 40#, they had good water retaining capacity, antioxidant capacity and osmotic regulation ability, thus they showed a relatively stong cold resistant, and have the potentiality of popularizing and planting in high latitudes or high altitude areas.
Ginkgo; clones; freezing stress; cold resistance; selection and breeding
S722.1+3
A
1673-923X(2012)12-0132-07
2012-06-13
高校博士点专项基金项目“银杏三个脂肪酸去饱和关键酶基因的克隆、表达与载体构建”( 20113204120007);国家科技支撑计划课题“银杏和印楝珍贵材用和药用林定向培育关键技术研究与示范”(2012BA021D04);林业公益性行业科研项目“银杏叶和外种皮加工产业化关键技术研究”(201004015)资助
郁万文( 1979-),男,山东临沂人,博士,讲师,主要从事经济林栽培的研究。E-mail: youeryuww@163. com
曹福亮(1957-),男,江苏姜堰人,博士导师,教授,主要从事经济林培育与加工研究
[本文编校:罗 列]
猜你喜欢
儿童时代·快乐苗苗(2022年10期)2022-12-09
蔬菜(2022年10期)2022-11-10
热带作物学报(2022年5期)2022-06-01
今日农业(2021年20期)2021-11-26
时代邮刊·下半月(2021年10期)2021-10-23
河北果树(2020年4期)2020-11-26
山西农业科学(2020年10期)2020-10-16
河北果树(2020年1期)2020-02-09
现代园艺(2018年2期)2018-03-15
浙江农业科学(2017年8期)2017-08-30
