
土壤水分对山杏光合作用日变化过程的影响
2012-01-02 08:32张征坤张光灿刘顺生裴斌徐志强徐萍刘霞
中国水土保持科学 2012年3期
张征坤,张光灿,刘顺生,裴斌,徐志强,徐萍,刘霞
(山东省土壤侵蚀与生态修复重点实验室,山东农业大学林学院,国家林业局泰山森林生态站,271018,山东泰安)
在黄土丘陵区,干旱缺水是抑制农业生产最突出的生态环境问题,开展植物生理过程与水分关系的研究,对造林树种的合理选择与立地配置、合理利用水土资源、加快荒山荒沟绿化速度、控制水土流失等,无论是在理论上还是在实践上都具有重要的指导意义[1]。目前,植物光合作用过程与机制仍然是国际上植物生理生态学研究的热点问题,并在强光、高温、低温、干旱等逆境生理研究中得到广泛应用,取得了令人可喜的成果[2-4]。植物光合生理生态过程的研究在国内也日益受到重视,其热点问题之一便是水分与光合作用的关系[5-7]。植物光合作用日变化过程是在一定天气条件下,各种生理生态因子综合效应的最终反映[8],目前的研究已深入到不同植物的光合能力、叶绿素荧光、羧化效率、量子产量等光合效率指标以及某些生化指标的日变化规律方面[9-12];但已有的报道多见于以农作物为对象的研究[13-16],而针对不同造林树种开展的研究还相对较少。
山杏(Prunus sibirica L.)为落叶小乔木,萌蘖能力强,生长迅速,容易繁殖,根系发达,是黄土高原以及北方土石山区植树造林的主要树种之一。目前,国内对山杏的研究多见于开发利用价值、栽培技术、耗水性能等方面[17]。在山杏光合作用特征及其与土壤水分关系的研究上,尽管已有关于土壤水分影响光合作用光响应过程的研究报道[18-19],但是有关山杏光合作用日变化过程与土壤水分的关系等问题还不十分清楚。笔者以2 年生盆栽山杏苗木为试验材料,通过测定其在土壤水分连续变化条件下的光合作用日变化,探讨土壤水分变化对山杏光合日变化过程的影响,以期为山杏在半干旱黄土丘陵区的合理栽培提供技术参考。
1 试验地自然概况
试验地设在山西省吕梁山西麓的方山县峪口镇土桥沟流域(E110°02'55″,N37°36'58″),属于黄河中游黄土丘陵沟壑区,流域内最高海拔1 446 m,试验区平均海拔1 200 m。多年平均降水量416 mm,6—9 月降水量占全年的70%以上。多年平均气温7.3 ℃,多年平均蒸发量652.9 mm,干燥度1.3,最大蒸发量出现在4—6 月,具有典型的春季干旱的特征。土壤为黄绵土,pH 值8.0 ~8.4。属于森林草原灌丛植被区,山杏在流域的坡面及沟谷内呈零散的分布。
2 材料与方法
2.1 试验材料与处理
选用生长情况基本一致的2 年生山杏苗木作为试验材料。在4 月份对试验材料进行盆栽培育,共栽植6 盆(每盆1 株)。用环刀法测得盆栽的土壤密度为1.20 g/cm3、田间持水量为23.1%。在6 月份进行不同土壤水分条件下光合作用指标的测定。采用人为控制水分的方法获取不同的土壤水分梯度,即在试验观测2 d 前给试验植株浇水,使土壤水分饱和;通过自然耗水2 d 后获得初期水分含量,进行第1 次光合生理参数的测定;以后每2 d 获取1种土壤水分含量,共形成8 个土壤水分梯度系列,分别进行光合生理参数测定。利用烘干法测定土壤的质量含水量(Wm,%),分别为21.5%、17.8%、15.3%、13.0%、11.4%、8.6%、5.9%和4.4%;利用Wm与田间持水量的比值计算土壤的相对含水量(Wr,%),分别为93.2%、77.0%、66.1%、56.1%、49.3%、37.3%、25.7%和19.1%。
2.2 光合作用日变化测定
分别从6 株试验植株的中部选取3 片生长健壮的成熟叶片,利用英国PPS 公司生产的CIRAS-2 型光合作用系统测定不同土壤水分条件下光合作用参数的日变化。观测时间为07:00—17:00,每2 h 测1次。在光合速率趋于稳定时,每个叶片重复记录3次数据,取平均值进行分析。仪器自动记录叶片的蒸腾速率(Tr/(mmol/(m2·s)))、净光合速率(Pn/(μmol/(m2·s)))、胞间CO2摩尔分数(Ci/(μmol/mol))、气温(θa/℃)、大气CO2摩尔分数(Ca/(μmol/mol))等生理生态参数。叶片水分利用效率(EL/(μmol/mmol))和气孔限制值(Ls/%)分别用下式计算:
日光合累积值和日蒸腾累积值的计算:将日变化测定的叶片净光合速率和蒸腾速率作累积处理,分别得到日光合累积值(PD/(μmol/(m2·d)))和日蒸腾累积值(TD/(mmol/(m2·d))),并由此计算得出日均水分利用效率(ELD/(μmol/mmol))[20],即:
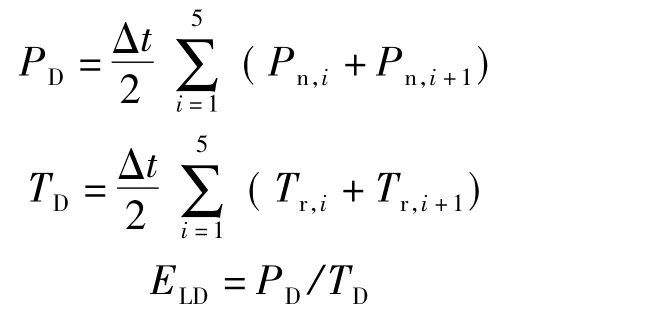
式中:Pn,i、Pn,i+1为相邻2 次测 定的叶 片净光 合 速率,μmol/(m2·s);Tr,i、Tr,i+1为相邻2 次测定的叶片蒸腾速率,mmol/(m2·s);Δt 为测定时间间隔,s,取7 200 s。
3 结果与分析
3.1 山杏叶片净光合速率的日变化
山杏叶片光合速率日变化及日光合累积值见图1。可以看出,不同土壤含水量山杏净光合速率Pn的日变化过程具有明显差别(图1(a))。当土壤相对含水量Wr在56.1%~66.1%之间时,山杏的光合作用没有发生午休现象,全天中的Pn相对较高,日变化过程为单峰曲线,Pn在09:00—11:00 时段的水平较高;当Wr在77.0%~93.2%之间时,山杏在全天中的Pn有所下降,日变化过程表现为不太明显的双峰曲线,Pn在09:00—13:00 时段的水平较高,但在11:00 左右出现微弱的午休现象;当Wr降低至49.3%以后,随土壤含水量减少,山杏在全天中的Pn明显降低,并且在13:00 左右的光合午休(Pn下降)现象逐渐加重。结合图1(b)可以看出:在Wr为56.1%时,山杏日光合累积值PD达到最高水平,显著大于其他土壤水分条件下的PD;当Wr在49.3%~93.2%之间时,山杏的PD水平较高,达到其最高水平的69%以上;当Wr降低至37.3%以后,山杏的PD显著下降。由此认为,山杏光合作用比较适宜的土壤相对含水量范围为49.3%~93.2%。
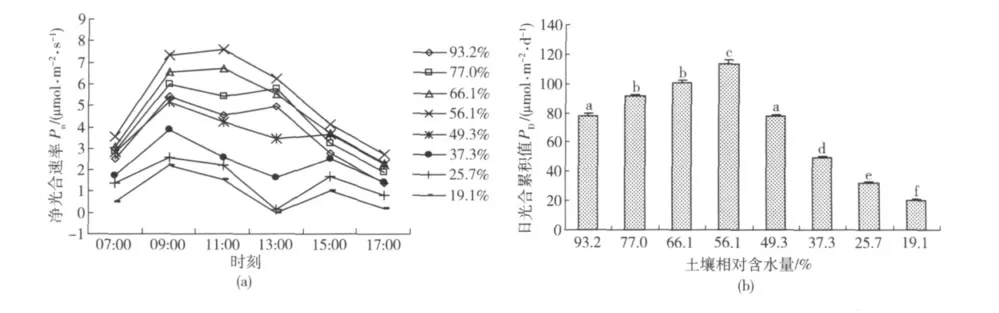
图1 不同土壤相对含水量山杏叶片光合速率日变化及日光合累积值Fig.1 Diurnal change of photosynthesis and the accumulation value of Prunus sibirica L.under different soil relative water content
3.2 山杏叶片胞间CO2摩尔分数和气孔限制值的日变化
山杏叶片胞间CO2摩尔分数Ci和气孔限制值Ls的日变化曲线见图2。可以看出,山杏叶片胞间CO2摩尔分数Ci和气孔限制值Ls的日变化过程随土壤含水量的变化而明显不同。当Wr大于或等于49.3%时,不同土壤含水量条件下山杏Ci和Ls在全天的变化过程大体上都表现为反抛物线和抛物线形式,在09:00—13:00 时段(即光合作用午休、Pn下降期间,图1(a))的变化趋势也基本相似,即表现为Ci逐渐下降(图2(a))和Ls逐渐增加(图2(b));当Wr降低至37.3%后,山杏Ci和Ls的日变化过程发生了明显变化,尤其在09:00—13:00 时段Pn下降期间(图1(a)),表现为Ci明显增加(图2(a))和Ls明显下降(图2(b))。
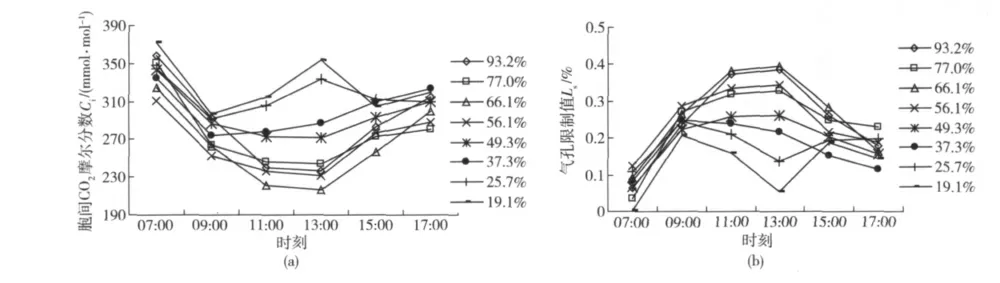
图2 不同土壤相对含水量山杏叶片胞间CO2摩尔分数Ci和气孔限制值Ls的日变化Fig.2 Diurnal change of Ci and Ls of Prunus sibirica L.under different soil relative water content
按照G.D.Farquhar 等[21]提出的光合作用限制因素判别标准,上述结果表明,不同土壤含水量条件下山杏光合作用午休的原因不同。即:当Wr大于49.3%时,山杏中午Pn下降的原因主要是气孔因素的限制(叶肉细胞CO2供应受阻);当Wr在49.3%~37.3%之间时,山杏光合午休的主要原因由气孔因素向非气孔因素转变;而当Wr降低至37.3%时,光合午休的主要原因已转变为非气孔因素的限制(叶肉细胞光合能力下降)。由此认为,山杏进行正常光合作用的土壤相对含水量最低值在37.3%左右。
3.3 山杏叶片蒸腾速率和水分利用效率日变化
山杏叶片蒸腾速率日变化及日累积值见图3。可以看出,在不同土壤含水量条件下,山杏叶片蒸腾速率Tr的日变化过程也会发生明显变化。Wr在56.1%~77.0%之间时,不同土壤水分条件下山杏全天的Tr水平较高,没有发生中午降低的现象,日蒸腾累计值TD之间没有显著差异(图3(b));但当Wr降低到49.3%以后,山杏Tr的日变化过程变为双峰曲线,呈现出与山杏光合速率日变化(图1(a))基本相似的过程,即随着土壤相对含水量降低,山杏在全天的和中午(11:00—13:00)的Tr水平逐渐降低,日蒸腾累计值产生显著差异(图3(b))。
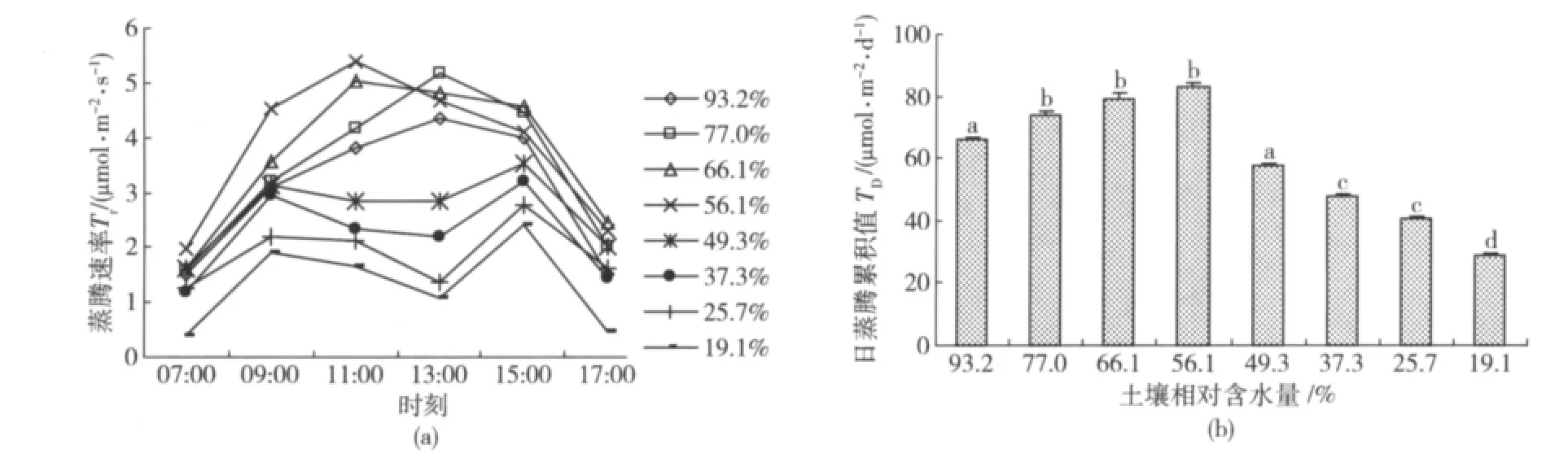
图3 不同土壤相对含水量山杏叶片蒸腾速率日变化及日累积值Fig.3 Diurnal change of transpiration rate and cumulative value of Prunus sibirica L.under different soil relative water content
山杏叶片水分利用效率的日变化及日平均值见图4。可以看出,山杏叶片水分利用效率EL日变化对土壤水分变化的阈值响应特征为(图4(a)):当Wr在49.3%~77.0%之间时,不同土壤水分条件下山杏的EL日变化过程基本一致,表现为上午(07:00—09:00)较高、至午后(15:00 左右)逐渐降低,下午(15:00 之后)有所回升的变化趋势,EL在中午阶段(11:00—13:00 光合作用午休期间)没有发生降低现象(图4(a))。结合图4(b)可以看出,山杏的日均水分利用效率ELD在56.1%时达到最高水平,但不同土壤水分条件下的ELD之间没有显著差异;当Wr增加到93.2%时,山杏的EL日变化没有发生明显变化,但ELD与其最高水平相比显著下降;当Wr降低至37.3%以后,山杏在全天的和中午期间(13:00 左右)的EL均会发生明显的下降,ELD也比其他土壤水分条件下的降低显著(图4(b))。上述结果表明,维持山杏具有较高水分利用效率的土壤相对含水量范围为49.3%~77.0%。
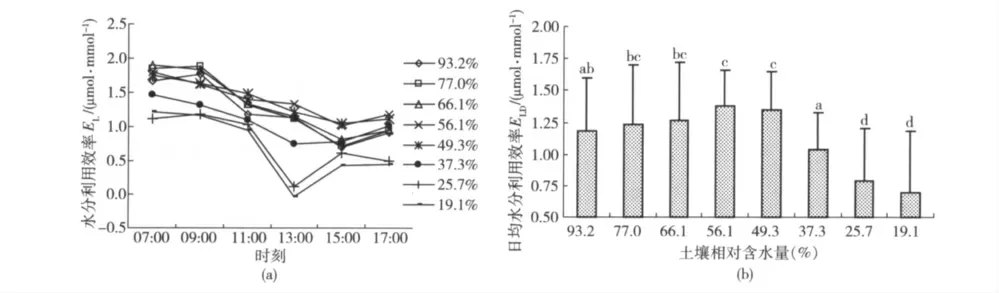
图4 不同土壤相对含水量山杏叶片水分利用效率的日变化及日平均值Fig.4 Diurnal change of water use efficiency and cumulative value of Prunus sibirica L.under different soil relative water content
4 结论与讨论
1)山杏光合作用、蒸腾作用的日变化过程以及光合作用午休的原因,对土壤相对含水量的变化具有明显的阈值响应。当土壤相对含水量在56.1%~66.1%之间时,山杏在全天的光合速率(Pn)和蒸腾速率(Tr)均较高,都没有发生午间降低的现象;当土壤相对含水量在93.2%~77.0%之间时,山杏在全天的Pn和Tr均有所下降,Pn日变化出现微弱的午间(11:00 左右)降低现象,主要原因是气孔因素限制;当土壤相对含水量在49.3%~19.1%之间时,随着土壤含水量的降低,山杏全天的Pn和Tr均发生显著下降,而且二者的日变化过程基本相似,都表现出明显的午间降低现象,光合作用午休的主要原因开始由气孔因素向非气孔因素转变;当土壤相对含水量为37.3%时,由于山杏叶片的胞间CO2浓度明显增加和气孔限制值明显下降,光合作用午休的主要原因已转变为非气孔因素的限制,即光合机构受到损伤、叶肉细胞光合能力下降。
上述现象表明,山杏蒸腾作用与光合作用的日变化过程,对土壤水分变化的响应规律基本一致,这可能是山杏适应土壤水分胁迫等不良环境的一种生理机制和生态策略。即在土壤水分过高或过低,以及中午强光、高温、低湿等不利于光合作用的环境下,山杏可以通过降低叶片的蒸腾作用来防止植株过度失水,同时提高水分利用效率。
2)山杏光合速率和水分利用效率(EL)的较高水平并非在土壤水分充足时获得,而是在适度的水分胁迫范围之内时获得。山杏Pn日累计值(PD)和EL日均值(ELD),在土壤相对含水量为56.1%时都达到最高水平,二者在土壤相对含水量为49.3%~77.0%之间时同时具有较高水平,不同土壤水分条件下的PD和ELD都能达到其最高水平的69%和89%以上。由此可知,山杏光合作用和高效用水的土壤相对含水量范围为49.3%~77.0%,这与通过不同土壤水分条件下山杏光合作用光响应过程研究[18]得出的结论(土壤相对含水量为44.7% ~80.9%)基本相同。对比其他树种具有较高光合作用和水分利用效率的土壤相对含水量范围,丁香(Syzygium aromaticum)的为59% ~76%[22]、核桃(Juglans regia)的为41%~60%[23]、刺槐(Robinia pseudoacacia)的为48%~64%[24]、沙棘(Hippophae rhamnoides Linn)的为41.5%~59.9%[25],可知,山杏是对土壤水分适应范围较广、抗旱能力较强的树种。
[1] 张淑勇,周泽福,张光灿,等.水分胁迫下天然次生灌木山桃和山杏光合气体交换特征[J].西北植物学报,2008,28(12):2492-2499
[2] Sharkyt D.Water tress effects on photosynthesis[J].Photosynthetica,1990,24:65-70
[3] Holmgren M.Combined effects of shade and drought on tulip poplar seedlings:trade-off in tolerance or facilitation[J].Oikos,2000,90:67-78
[4] Nobel P S.Physicochemical and environmental plant physiological responses to environmental parameters[J].San Diego: Academic Press,1999:275
[5] 王克勤,王斌瑞.土壤水分对金矮生苹果光合速率的影响[J].生态学报,2002,22(2) :206-214
[6] 肖春旺,周广胜.不同浇水量对毛乌素沙地沙柳幼苗气体交换过程及其光化学效率的影响[J].植物生态学报,2001,25(4):444-450
[7] 张往祥,曹福亮.高温期间水分对银杏光合作用和光化学效率的影响[J].林业科学研究,2002,15(6):672-679
[8] 赵会杰,邹琦,于振文.叶绿素荧光分析技术及其在植物光合机理研究中的应用[J].河南农业大学学报,2000,34(3):248-251
[9] 许大垒,徐宝基,沈允钢.C3 植物光合效率的日变化[J].植物生理与分子生物学学报,1990,16(1):1-4
[10]Kuppers M,Wheeler A M,Kuppers B I L,et al.Carbon dioxide fixation in eucalypts in field analysis of diurnal variations in photosynthetic capacity[J].Oecologia,1986,70:273-282
[11]Wise R R,Sparrow D H,Ortiz Lopez,et al.Biochemical regulation during the midday decline of photosynthesis in field grown sunflower[J].Plant Sci,199l,74:45-52
[12]Tenhunen J D,Lange O L,Gebel J,et al.Changes in photosynthetic capacity,carboxylation efficiency,and CO2compensation point associated with midday stomatal closure and midday depression of net CO2exchange of leaves of Quercus Suber[J].Planta,1984,162:193-203
[13]Poskuta J W,Nelosn C J.Role of photosynthesis and photorespiration and of leaf area in detemining yield of tall fescue genotypes[J].Photosynthetica,1986,20:94-101
[14]Govindjee.Photosynthesis:development,carbon metabolism,and plant Productivity[J].New York: Academic Press,1982,2:499-511
[15]许大全.光合作用效率[M].上海: 上海科技出版社,2002:163-170
[16]付芳婧,赵致,张卫星.水分胁迫下玉米抗旱性与光合生理指标研究[J].山地农业生物学报,2004,23(6):471-474
[17]王百田,张府娥.黄土高原主要造林树种苗木蒸腾耗水特性[J].南京林业大学学报:自然科学版,2003,27(6):93-97
[18]郎莹,张光灿,张征坤,等.不同土壤水分下山杏光合作用光响应过程及其模拟[J].生态学报,2011,31(16):4499-4508
[19]夏江宝,张光灿,孙景宽,等.山杏叶片光合生理参数对土壤水分和光照强度的阈值效应[J].植物生态学报,2011,35(3):322-329
[20]梁宇,高玉葆,陈世苹,等.干旱胁迫下内生真菌感染对黑麦草实验种群光合、蒸腾和水分利用的影响[J].植物生态学报,2001,25(5):537-543
[21]Farquhar G D,Sharkey T D.Stomatal conductance and photosynthesis[J].Annual Review of Plant Physiology,1982,33:317-342
[22]陈新军,张光灿,周泽福,等.黄土丘陵区紫丁香叶片气体交换参数的日变化及光响应[J].中国水土保持科学,2004,2(4):102-107
[23]李小磊,张光灿,周泽福,等.黄土丘陵区不同土壤水分下核桃叶片水分利用效率的光响应[J].中国水土保持科学,2005,3(1):43-47
[24]张光灿,刘霞,贺康宁.黄土半干旱区刺槐和侧柏林地土壤水分有效性及生产力分级研究[J].应用生态学报,2003,14(6):858-862
[25]张益源,贺康宁,董梅,等.水分胁迫对银水牛果和沙棘叶水势日过程及水分利用效率的影响[J].中国水土保持,2011(4):22-25
猜你喜欢
现代农村科技(2022年11期)2022-12-16
农业工程学报(2022年8期)2022-08-08
农业工程学报(2022年5期)2022-06-22
水土保持学报(2022年3期)2022-05-26
河南水利与南水北调(2022年1期)2022-03-16
现代农村科技(2021年11期)2021-11-05
现代农村科技(2021年3期)2021-04-11
今日农业(2019年14期)2019-01-04
中国农业信息(2017年16期)2017-11-02
种子科技(2017年8期)2017-09-03
