
小型底栖动物生物量估算方法回顾与思考
2011-12-28 08:17:40张青田胡桂坤
海洋通报 2011年3期
张青田,胡桂坤
(天津科技大学 天津市海洋资源与化学重点实验室,天津 300457)
小型底栖动物生物量估算方法回顾与思考
张青田,胡桂坤
(天津科技大学 天津市海洋资源与化学重点实验室,天津 300457)
小型底栖动物是海洋生态系统的重要组成部分,其生物量估算的准确性影响着海洋生物资源分析和生态系统动力学的研究。对常用的小型底栖动物生物量估算方法进行了介绍,并指出了我国在使用体积估算法时存在的一些问题。在此基础上,对小型底栖动物生物量估算模型研究提出了几条建议。
小型底栖动物;生物量;估算方法
1 小型底栖动物概念及其生物量估算意义
小型底栖动物(meiofauna)的研究历史并不太长,但由于其为海洋生态系统动力学、生物地化循环和全球变化研究提供了重要背景和必要参数,逐渐成为了世界上海洋生态学研究的一个热点[1-2]。
从筛分角度来说,小型底栖动物通常指凡能通过孔径为0.5 mm套筛网目,而被孔径为0.042 mm网筛所阻留的底栖动物。从分类学角度来说,小型底栖动物主要指多细胞后生动物,但也包括部分原生动物,如有孔虫和纤毛虫等。截至目前,国内相关研究并未将原生动物包括在小型底栖动物范围内,仅指多细胞后生动物。在海洋无脊椎后生动物(metazoa)的33个门类(Phyla)中,22个门包含有小型底栖动物,其中动吻类(Kinorhyncha)、颚咽类(Gnathostomulida)、缓步类(Tardigrada)和铠甲类(Loricifera)等门类是小型底栖动物特有的。在上述小型底栖动物中,种类丰富度较高且现存量较大的优势类群为自由生活线虫、桡足类(猛水蚤为主)等。它们是海洋异养集合的重要组成部分,在底栖食物网中占据十分重要的地位,影响整个生态系统的物质和能量传递[3]。最初,人们认为小型底栖动物物与大型底栖动物之间的区分是人为的,其间并无明显的生物学界限。但后来研究认为小型底栖动物实际上应为一个独立的功能单位,具有与大型底栖生物不同的生活史和生殖对策[4]。
在河口和沿海底栖生态系统中,小型底栖动物是初级生产和较高营养级之间的重要联系者,不仅是小型光合底栖生物的重要消费者[5,6],而且是其他底栖动物以及鱼类幼体的食物源[7,8]。毋庸置疑,在能量流动和生态系统动力学研究中,生物量数据远比丰度(密度)数据重要。因此,在60余年前,人们就开始了小型底栖动物生物量计算方法的研究。中国缺乏对小型底栖动物生物量的系统性研究,仅简单地照搬国外的现有公式进行生物量估算。这影响了小型底栖动物生物量的准确度,进而影响了底栖生物资源的分布研究以及环境影响的分析[9]。为了提高海洋生物资源分析和生态系统动力学研究的准确度,亟需开展小型底栖动物生物量估算模型的研究。
2 生物量估算方法及存在问题
2.1 生物量估算方法
计算小型底栖动物生物量的方法有2大类,一是直接称重法,二是通过体积换算。无论是测量湿重,还是干重,直接称量法都需要使用超微量分析天平(感量0.1 µg),可能还需要配套的水分测定仪;而且这种方法对样品损害大,不利于后续分析,所以目前更多的学者采用测量估算的方法[9,10]。
国外对于体积估算的研究较早、较多,虽然所用公式可能不同(后文举例),但是基本思路和原理是一致的。即通过测量小型底栖动物的某些指标(体长,体宽等)来计算该生物的体积,然后乘以特定的密度(比重)值,得到个体重量值;再由个体重量值和丰度值估算小型底栖动物生物量。影响力比较大的研究人员和成果包括:Andrassy(1956),Wieser(1960),Warwick和 Price(1979),Warwick和 Gee(1984),Riemann等(1990),Gradinger 等(1999)等等[11-16]。
已有公式中,用于估算生物量的生物密度值通常采用1.13,这是近年来国内外研究海洋小型底栖动物普遍采用的数值。研究表明,这个密度值和小型底栖动物所处环境有关,以线虫为例,所有海水线虫的密度值一般记为1.13,而淡水线虫密度值为1.02,陆地线虫密度值则为1.084[11,17],相差较大。用不同公式估算的个体干重值并不完全相同,和称重法的结果也有差异(参见表1,以自由生线虫为例),这些差异和生物种类组成及网筛孔径等有关,当然也受到了公式中经验参数的影响。表1中的最大干重值是最小值的30.29倍,对整体生物量的估算影响巨大。用“失之毫厘,谬之千里”的成语来提醒人们注意些许的差异是很有必要的,因为线虫等小型底栖动物的丰度值很高(达数千个/10 cm2),相关参数的些许差异足以造成生物量结果的巨大差异。
中国近海已经开展了小型底栖动物生物量的研究,从20世纪90年代陆续有相关文献报道,积累了一些数据和研究经验。但是由于小型底栖动物生物量数据的获得比计算丰度值困难,所以其研究报道明显少于丰度的报道。目前有一些关于渤海、黄海等海域小型底栖动物生物量分布的报道。黄勇(2005)等进行了线虫个体干重的计算工作[4];Zhang 等(2010)提出了分段计数法,用来提高生物量估算的准确度,但是没有涉及体积计算模型的研究[9]。
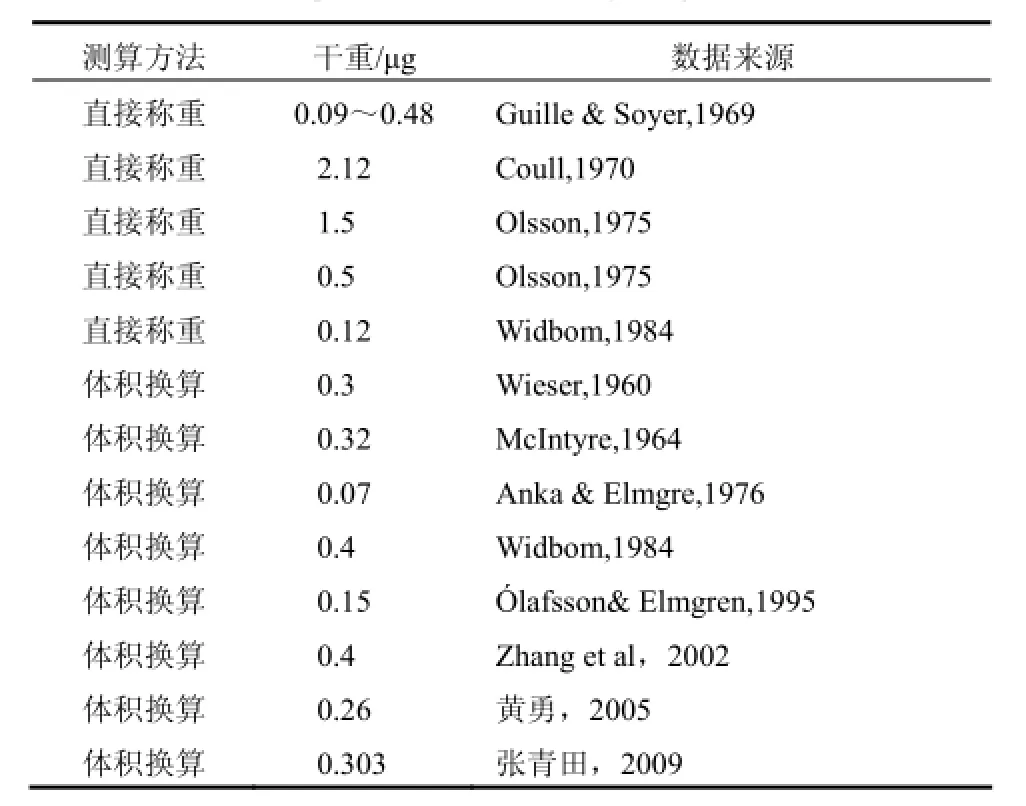
表 1 线虫个体干重值的比较[2,4]Tab.1 Comparison of individual dry weight of nematode
2.2 存在问题
在研究过程中,我们发现现有的体积估算方法存在缺陷,严重影响了生物资源分布和能量相关的研究。主要问题有以下几个方面:
(1)直接引用他人参数。直接引用他人的个体干重值而不自己测量会影响生物量值的准确度。目前我国研究常引用的线虫个体干重值为0.4 µg,由前述可知,其值偏大。黄勇(2005)在测量了1 382条线虫后提出了对用0.4 µg作为线虫个体干重值的担忧,认为该数值似乎过高了[4]。还有,在对渤海同样时间和站位的分析时,某些人采取了0.4 µg的线虫个体干重值,而其他人却用了0.16 µg,前者是后者的2.5倍。这势必给生物资源量生态分布的分析等研究带来麻烦。
(2)忽略了生物类群及个体间差异。小型底栖动物的分布和底质类型有很大关系,故而不同海域的生物组成是有差异的,这也会影响个体干重值的差异,从而成为不同学者研究结果(表1)差异的主要原因之一。因此,在所有海域用同一个个体干重值是不合适的;用单一的计算方程是不妥的。以渤海已有报道为例,线虫个体干重从0.06 µg(Tershellingia austenae)至2.08 µg(Belbollazhangi)[18],因种类不同引起的差异巨大。并且有研究表明,自由生活海洋线虫的个体干重值也会随季节和恶劣天气而变化[19,20]。这些足以证明即使线虫丰度值相同时,其生物量也不一定相同。也就是说,鉴于不同地点的线虫种类不同,发育阶段也不一定相同,采用单一的个体干重均值来估算小型底栖动物生物量是不妥当的。这不但影响了生物资源的估算,而且影响生物量与环境因子相关性分析[9]。
(3)估算公式不统一,准确度低。为便于比较和说明,下面列出几个主要估算公式(涉及线虫和桡足类,其他种类省略):
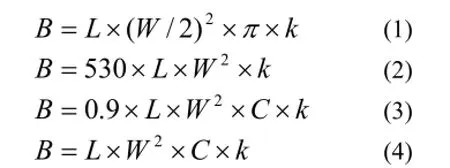
上述公式中,B代表生物量湿重。公式1是Wieser(1960)关于线虫生物量估算公式[12],式中,L为个体长度;W为食管中间部位的体宽;k为生物比重,海洋线虫取1.13。公式2是Nozais 等(2005)关于线虫生物量估算的公式[8],式中,530为线虫的转换系数;L为个体总长(mm);W为个体最大体宽(mm);k值(μg/nl)同公式1。公式3是Nozais 等(2005)关于桡足类生物量估算的公式[8],式中,L和k与公式2相同;0.9为转换系数;W为体宽(mm);C为体形相关的转换系数,取值400~560。公式4是国家海洋局908专项办公室[21]和国家质量监督检验检疫总局[22]提供的估算公式,适用于小型底栖动物各主要类群。式中,L为体长(mm),长尾种类至锥状部,具丝状尾种类至肛门;W为身体最大宽度(mm);C为转换系数,和生物种类相关;k和前述一致。在计算线虫生物量时,公式2和公式3是一样的;但是计算桡足类时,公式4比公式2少了一个转换系数(0.9),其他转换系数基本一致。没有查到国内关于公式4的由来和推导,估计是引自国外资料,但是为什么省略一个转换系数则不可考。可见,用不同公式计算的小底栖动物生物量值是有差异的,并且已有公式中的经验值是在较早年代的简陋条件下获得的,制约了计算准确度。由以上公式和资料也可以知道,国外一直都在沿着体积估算的思路对公式进行修正,并未死守现有结果;不同研究区域的公式也不完全一样。
(4)估算模型中参数过少,难以准确描述种类差异。目前的估算方法(模型)往往只测量长、宽指标,而小型底栖动物在分类中有多项参数需要测量,例如包含线虫的De Man公式在内的体长、口咽腔等30余个测量参数[4]。既然不同类别的体型存在差异,势必引起体积估算模型的差异。引入更多的分类学测量参数来建立新的体积和生物量估算模型是一项很有意义的工作。
综上所述,照搬国外现有公式将会影响小型底栖动物生物量的估算准确度,进而影响生物资源的分布研究以及环境影响的研究。在生态系统动力学研究日益受到重视的前提下,更需要准确的小型底栖动物数据支持。吴增茂等(2001)在国内首次进行了浅海生态的水层-底栖系统动力耦合研究[23],与水层模型模拟结果相比,耦合模型的结果更加合理。这也证实了小型底栖动物生物量研究的迫切性和必要性。中国对于底栖亚系统的定量研究还相当少,远不能适应动力学模型研究的需要。开展相关研究将在一定程度上减轻海洋生态学研究的阻力,促进能量流动和生态系统动力学的研究,具有较高的科学意义和应用价值。还可以丰富中国对于小型底栖动物研究内容,促进国际交流合作,迎头赶上世界先进水平。
3 总结与建议
(1)中国小型底栖动物生物量估算过程中存在参数引用不当和照搬公式等问题,制约了生物资源分析和生态系统动力学等方面的研究。现有一些参数是在比较简陋条件下测得的,而当前先进的显微测量技术和计算机技术为参数测定和模型拟合提供了良好的研究基础。因而亟需在我国海域开展生物量估算模型的研究工作。研究时应该注意不同环境下生态类群的差异,选择河口、泥沙、砂质等环境开展工作,以保证模型的适用性。
(2)参考国外研究思路,这里以体积换算法为主线,舍弃直接称重法。这样,不但可以和国际上已有研究相统一,也可以为小型底栖动物研究保留生物样品,很适合中国研究相对薄弱的现实情况。不限于体宽、体长的参数,引入分类学中测量的一些参数,建立适合我国海域,并且准确度高于已有模型的小型底栖动物生物量估算模型。
受当时研究条件所限,以前的估算方程过于简单,影响了估算精度。新的计算方程并不唯一,和生物类群有关;在计算机技术的帮助下,现在可以选择较复杂的模拟方程用来计算生物量,提高计算准确度。为降低操作难度,减小计算过程中的错误几率,编写出计算小型底栖动物生物量的程序软件。借助软件来降低操作难度、增加估算准确度也是国际同行的思路[24]。在应用中,只需输入测定的特征值即可自动选择适宜方程,完成体积计算、数据统计分析等工作,化繁为简,便于更多人操作。
[1]张志南,周红.国际小型底栖生物研究的一些进展 [J].中国海洋大学学报, 2004, 34(5): 799-806.
[2]张青田.天津近岸海域小型底栖动物生态学研究 [D].天津: 南开大学博士学位论文, 2009: 1-161.
[3]Li J, Vincx M, Herman P M J.Carbon flows through meiobenthic nematodes in the Westerschelde estuary [J].Fund and Appl Nematology, 1997, 20: 487-494.
[4]黄勇.南黄海小型底栖生物生态学和海洋线虫分类学研究[ D].青岛:中国海洋大学博士学位论文, 2005: 1-184.
[5]Moens T, Luyten C, Middelburg J J, et al.Tracing organic matter sources of estuarine tidal flat nematodes with stable carbon isotopes [J].Mar Ecol Prog Ser, 2002, 234: 127-137.
[6]Pinckney J L, Carman K R, Lumsden S E, et al.Micro- algalmeiofaunal trophic relationships in muddy intertidal estuarine sediments [J].Aqua Microb Ecol, 2003, 31:99-108.
[7]Tita G, Desrosiers G, Vincx M, et al.Predation and sediment disturbance effects of the intertidal polychaeteNereis virens(Sars) on associated meiofaunal assemblages [J].Journal of exp Mar Biol and Ecol, 2000, 243:261-281.
[8]Nozais C, Perissinotto R, Tita G.Seasonal dynamics of meiofauna in a South African temporarily open/closed estuary (Mdloti Estuary, Indian Ocean) [J].Estuar, Coast and Shelf Sci, 2005, 62:325-338.
[9]Zhang Q T, Wang X H, Hu G K.Evaluation of estimation methods for meiofaunal biomass from a meiofaunal survey in Bohai Bay [J].Chinese J of Oceanogr and Limnol, 2010, 28(1):82-87.
[10]郭玉清, 张志南, 慕芳红.渤海小型底栖动物生物量的初步研究[J].海洋学报, 2002, 24(6): 76-83.
[11]Andrassy I.Die Ramninhalts- und Gewichtsbestimmung der Fadenwuermer (Nematoden) [J].Acta Zool.(Budapest) , 1956, 2:1-15.
[12]Wieser W.Benthic studies in Buzzards bay II the meiofauna [J].Limnol and Oceanogr, 1960, 5(2):121-137.
[13]Warwick R M, Price R.Ecological and metabolic studies on free-living nematodes from an estuarine mud-flat [J].Estuar, Coast and Shelf Sci, 1979, 9:257-271.
[14]Warwick R M, Gee J M.Community structure of estuarine meiobenthos [J].Mar Ecol Prog Ser, 1984, 18:97-111.
[15]Riemann F, Ernst W, Ernst R.Acetate uptake from ambient water by the free-living marine nematodeAdoncholaimus thalassophygas[J].Mar Biol, 1990, 104: 453-457.
[16]Gradinger R, Friedrich C, Sprindler M.Abundance, biomass and composition of the sea ice biota of the Greenland Sea pack ice [J].Deep-Sea Res Pt II Topical Studies in Oceanography, 1999, 46: 1 457-1 472.
[17]Nielsen C O.Studies on the soil microfauna.II The soil inhabiting nematodes [J].Nature Jutlandica, 1949, 2: 1-131.
[18]Guo Y Q, Warwick R M, Zhang Z N.Freeliving marine nematode as a pollution indicator of the Bohai Sea [J].J of Environ Sci, 2002, 14(4): 558-562.
[19]Mu F H, Somerfield P J, Warwick R M, et a1.Large-scale spatial patterns in the community structure of harpacticoid copepods in the Bohai Sea, China [J].The Raffles Bullet of Zool, 2001, 50(1): 17-26.
[20]华尔, 张志南, 张艳.长江口及邻近海域小型底栖生物丰度和生物量.生态学报, 2005, 25(9): 2235-2242.
[21]国家海洋局908专项办公室.海洋生物生态调查技术规程 [M] .北京: 海洋出版社.2006.
[22]国家质量监督检验检疫总局.海洋调查规范(海洋生物调查, GB/T12763.6-2008) [S].北京: 国家标准局出版社, 2008.
[23]吴增茂, 翟雪梅, 张志南, 等.胶州湾北部水层-底栖耦合生态系统的动力数值模拟分析 [J].海洋与湖沼, 2001, 32(6): 588-597.
[24]Baguley J C, Hyde L J, Montagna P A.A semi-automated digital microphotographic approach to measure meiofaunal biomass [J].Limnol Oceanogr:methods, 2004, 2: 181-190.
Review and thinking about the estimation method of meiofaunal biomass
ZHANG Qing-tian, HU Gui-kun
(Tianjin Key Laboratory of Marine Resources and Chemistry, Tianjin University of Science and Technology, Tianjin 300457, China)
Meiofauna is one of the important parts of marine ecosystem, and the estimation veracity of meiofaunal biomass would affect the analysis of marine bio-resources and the study on ecological dynamics.The general estimation methods of meiofaunal biomass were introduced briefly in this paper, and we also pointed out some problems which appeared when the estimation method was used in China.Based on this, some advice about estimation study of meiofaunal biomass was given.
meiofauna; biomass; estimation method
Q178.1
A
1001-6932(2011)03-0357-04
2010-07-31;收修改稿日期:2011-01-26
天津科技大学引进人才科研基金(20090413);国家科技支撑计划重点项目(2010BAC68B04);天津科技大学科研基金项目(20100220)。
张青田 ( 1974- ),男,博士,副教授,主要从事海洋生物学方面的教学、科研工作。电子邮箱:qtzhang@163.com。
猜你喜欢
今日农业(2022年14期)2022-09-15 01:44:26
今日农业(2020年19期)2020-12-14 14:16:52
建筑科技(2018年6期)2018-08-30 03:40:54
中成药(2018年2期)2018-05-09 07:20:04
浙江大学学报(农业与生命科学版)(2017年1期)2017-04-17 07:05:02
中国交通信息化(2016年5期)2016-06-06 03:51:43
天然产物研究与开发(2016年6期)2016-06-05 10:29:30
中国科技信息(2015年2期)2015-11-16 08:18:32
植物营养与肥料学报(2014年1期)2014-03-11 20:24:38
天津冶金(2014年4期)2014-02-28 16:52:58
